
Abstract
One strategy to identify neurochemical pathways of addiction is to map the relevant genes. In the present study we used 43 B6.C and 35 B6.I inbred RQI mouse strains, carrying <3% donor genome on C57BL/6ByJ background, for gene mapping. The strains were phenotyped for consumption of alcohol (12% v/v) in a two-bottle-choice paradigm, and genotyped for 396 microsatellite markers. The current mapping study extends our earlier experiment scanning five mouse chromosomes (Vadasz et al. (2000) Scanning of five chromosomes for alcohol consumption loci. Alcohol 22:25–34) to a whole-genome study, and discusses the differences and limitations. Data were analyzed with composite interval (CIM) and multiple interval (MIM) QTL mapping methods. CIM of B6.C strains detected significant QTLs on chrs. 6 and 12. A suggestive, but not significant, locus was detected in the B6.I strains on chr. 12. The best MIM model for B6.C strains confirmed one QTL on chr. 6 and one QTL on chr. 12, while the MIM model for the B6.I strains confirmed the suggestive locus on chr. 12. Some of the QTLs for alcohol consumption are new, while others confirm previously reported QTLs for alcohol preference, and alcohol acceptance.
Similar content being viewed by others
Introduction
The basic neurochemical processes underlying alcoholism and drug addiction are not well understood. One strategy to identify neurochemical pathways of addiction is to map the relevant genes. Accumulated evidence in quantitative genetics suggests that most complex traits and diseases (including alcoholism) are substantially affected by genetic factors. Genetic variation in alcohol drinking was first described by McClearn and Rodgers nearly half a century ago by comparing well established inbred mouse strains [1]. However, in spite of intensive research in the past decades, the biological basis of oral alcohol self-administration is not well understood. Recent availability of new genetic tools, such as high throughput SNP genotyping and gene expression microarrays, raise hope that many of the genes responsible for alcoholism can be identified in the near future. This is particularly important because individually effective therapy can only be expected if the molecular interaction between the genetic basis and the affected biochemical pathways are well understood.
Alcohol preference in rodents is accepted as an important animal model for hedonic aspects of alcoholism. Since the early 1990s, a large number of Quantitative Trait Loci (QTLs) have been mapped in genetic studies on alcohol preference, consumption, and acceptance. After many years of intensive work and detection of numerous QTLs by several laboratories using C57BL/6J and DBA/2J based genetic designs, only very few QTLs have been considered as consistently confirmed on chrs. 2, 3, 4, and 9 [2]. Recently, retinaldehyde binding protein 1 (Rlbp1) and syntaxin 12 (Stx12) [3, 4], and syntaxin binding protein 1 (Stxbp1) [5] were found to be strong candidate genes for alcohol preference drinking. All the above studies relied on the “gene pool” of two inbred strains, C57BL/6J and DBA/2J. Because in the species many gene variants may segregate which are not represented by these two strains, a tendency to carry out most studies only on two strains and their descendants will limit our understanding at the species level, and we will be able to identify fewer genes in animal models for testing in human alcoholism. Accordingly, there is a need to identify the most representative genes at species level. To this end, only a few laboratories devoted efforts to use strains other than C57BL/6J and DBA/2J in mapping QTLs for alcohol consumption and preference (e.g., A/J [6, 7] and 129P3/J [8]). Recently, HAP/LAP selection lines were developed [9] from HS/Ibg mice and used in mapping QTLs for alcohol preference [10]. The HS/Ibg mice were derived from eight inbred strains, including A, BALB/c, C57BL/6, DBA/2 [11].
Differences in alcohol drinking behavioral phenotypes between C57BL/6ByJ, BALB/cJ, CXBI/ByJ, progenitors and RQI strains have been reported previously [12–14]. We developed a complex trait gene mapping strategy, Recombinant QTL Introgression, [15–19], which was applied to map QTLs on five chromosomes for alcohol preference and consumption in 80 RQI strains of the b5i7 series [12]. In this study we scanned five mouse chromosomes [1, 2, 8, 12 and 15] with polymorphic microsatellite markers for QTLs for alcohol consumption. We used 44 B6.C and 36 B6.I inbred congenic Recombinant QTL Introgression (RQI) mouse strains of the b5i7 series carrying genes of BALB/cJ or CXBI origin on C57BL/6ByJ genetic background. In the B6.C set of strains, multiple regression analysis yielded a model with three microsatellite markers, which explained 32% of the genetic variance. The two markers with the highest significance levels in the model, D1Mit167 and D2Mit74, have been mapped to chromosome regions close to the gene opioid receptor kappa 1 (chr. 1) and opioid receptor kappa 3 (chr. 2), respectively. The results of this gene-mapping study suggested that genetic polymorphisms in kappa opioid receptors may contribute to genetic predisposition to voluntary alcohol-drinking behavior. Our follow-up studies on opioid receptor kappa 1 knock-out mice indicated that complete constitutional dysfunction of the Oprk1 gene can reduce alcohol consumption [20], however chromosome position of microsatellite marker “ D1Mit167” in the MIT database was assigned incorrectly, and the proposed chr. 1 region was not associated with genetic variation in alcohol consumption [21].
Here we present results of a study, in which we [12] expanded genome scanning to include all autosomal chromosomes, and [1] increased sample sizes in alcohol preference tests of b5i7 RQI strains. The combined data were analyzed by composite interval mapping (CIM) which shows a better performance than interval mapping in the case of multiple linked QTLs [22], and by multiple interval mapping (MIM).
Methods
Animals
The care and use of animals met the standards and recommendations of the IACUC of the Nathan S. Kline Institute for Psychiatric Research in accordance with US Department of Agriculture and US Public Health Service guidelines.
Nomenclature
B6, C57BL/6ByJ (background strain); C, BALB/cJ (donor strain), I, CXBI/By (donor strain); B6.C introgression type, strain set in which BALB/cJ donor segments are distributed on B6 background; B6.I, strain set in which CXBI donor segments are distributed on B6 background; b5i7, strain developed by 5 backcrosses and 7 intercrosses; b4i5, strain developed by 4 backcrosses and 5 intercrosses. We use abbreviated RQI strain names. For example, the full name of one of the RQI strains is B6.Cb5i7-β3/Vad, and it is abbreviated as C5B3. The first letter, C (or I), stands for the donor strain name; 5 (or 4) designates the backcross-intercross series b5i7 (or b4i5); B (or A) indicates the replicate line β (or α); and the last character, 3, is the identification number of the strain (the numbers range from 1 to 34 in each replicate line).
Recombinant QTL introgression strains
B6.C and B6.I inbred quasi-congenic RQI mouse strains were constructed as described previously [12, 15, 16, 23]. Recombinant QTL Introgression is a method, which was developed for the genetic analysis of complex quantitative traits by combining short-term phenotypic selection, congenicity, recombination and inbreeding [18]. The method can collect separately the inheritable increaser or decreaser factors, which control the phenotype, and preserve them in recombinant form in numerous quasi-congenic (near-isogenic) highly inbred strains. For phenotype a mesotelencephalic dopamine-system related trait was chosen because of its critically important role in the control of motor activity, motivation, emotion, addiction, and learning. Briefly, QTLs that are responsible for the continuous variation of mesencephalic tyrosine hydroxylase (TH/MES) activity were introgressed onto B6 background strain from BALB/cJ and CXBI donor strains. CXBI is a recombinant inbred strain carrying B6 and BALB/cBy genes [24]. Two types of F2s, (B6XC and B6XI), were produced, and, in each type, replicate lines (α and β) were created by equal division of each F2 litter. In each of the four lines, at least 45 F2 males were tested for the phenotype, and 15 were selected for the first backcross to B6 females. Then, at least 45 backcross1 (b1i0) male offspring were tested, and 15 males were selected and intercrossed with nonlittermate females, resulting in b1i1 generation. The QTL transfer was carried out in two directions by backcross–intercross cycles with concomitant selection for the extreme high and low expressions of TH/MES activity in replicates, resulting in four QTL introgression lines. In these lines, the top and bottom one third of each generation in the expression of TH/MES activity was selected. These steps were repeated for four (b4i5 series) or five (b5i7 series) cycles. The QTL introgression phase was followed by initiation of brother sister (b × s) mating for at least 30 generations in closed lines. RQI strains of the b4i5 and b5i7 series carry <5% and <3% of the donor BALB/cJ genome, respectively, on the background B6 genome.
Eight sublines of the b5i7 series (C5A26, C5A32A, C5A3A, C5B19A, C5B5A, I5A16at, I5B1A, I5B25) have been excluded from the analysis for ensuring independent derivation of strains from the population of the last backcross–intercross cycle [cf. 12].
Average of donor genome contribution in carrier strains was calculated as Q% = N CC/(N CC + N BB)*100 ± SD, where N CC = number of markers with CC (donor) genotype; N BB = number of markers with BB (background) genotype. Estimates of donor genome contribution were based on RQIgbase, release November 20, 2004 (http://rqigenetics.org/).
Behavioral tests
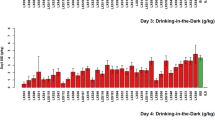
Adult (11 ± 2-week-old ) male mice of C57BL/6By, BALB/cJ, CXBI, and RQI strains were used and had been in the study room for at least 1 week prior to the behavioral tests [12]. The capacity of the testing system allowed us to carry out ethanol-preference tests in batches of 70 subjects. Depending on the availability and total number of animals tested, in each batch 3 littermate males of available RQI strains were tested, along with 3 males of each of the progenitor strains. The latter served as standard reference throughout the phenotyping to assess inter-assay variability. We used a “two-bottle choice” paradigm with escalating ethanol concentration. The test consisted of five 3-day trials, in which mice were allowed to choose between ethanol solution and tap water. To acclimate the animals to the taste of ethanol, the ethanol solution was offered in escalating concentrations: a 3% solution for trial 1 (day 1–3) was increased to 6% in trial 2 (day 3–6), and further increased to 12% for trials 3 (day 6–9), 4 (day 9–12), and 5 (day 12–15). This arrangement provided triplicate measures of ethanol preference at 12% concentration (v/v).
Because offering ethanol solutions of 3% and 6% served the purpose of acclimation of animals to 12% solution, we did not wish to carry out experiments in duplicate for 3% and 6% ethanol consumption. Our data analysis focused on consumption of 12% ethanol solution, measured in triplicate (i.e., three subsequent 3-day trial periods), the values used for analysis were 3-day access values. The liquids were offered in custom-made drinking tubes composed of centrifuge tubes fitted with single-hole rubber stoppers into which stainless steel sippers were inserted. Stainless-steel washers were glued to the bottom part of the rubber stoppers, which faced the cage-covers, to prevent chewing and playing with the drinking tubes. We also used stainless-steel springs to fasten the tubes firmly to the top of the cage covers. Two empty cages with alcohol and tap water drinking tubes were put on the racks to obtain control weights of leakage and evaporation. The position of the water and alcohol drinking tubes on the cage cover was alternated in each 3-day preference trial to avoid a position effect. The weights of the drinking tubes were measured before and after a 3-day trial by an A&D electronic analytical balance connected to an IBM AT computer. Data were entered automatically using A&D COLLECT software and QUATTRO spreadsheets. Alcohol consumption data previously collected for RQI strains [12] and new data collected in this study using the same methods were co-analyzed.
Alcohol consumption (AC) phenotype definition
AC34GKD represents alcohol consumption g BW −1kg day−1. Consumption of 12% alcohol was averaged over trials 3 and 4 for an individual, then mean value for a strain was obtained across individuals. For the b4i5 series of B6.C RQI strains only trials 3 and 4 were tested systematically, the AC34GKD phenotype does not include trial 5 data. For the b5i7 series of B6.C and B6.I RQI strains data for trials 3, 4, and 5 were available. Accordingly, for AC345GKD consumption of 12% alcohol was averaged over trials 3, 4 and 5 for an individual, then mean for a strain was obtained across individuals, and expressed as g BW −1kg day−1. AC34GKD and AC345GKD data are shown in Tables 1 and 2.
DNA extraction, polymerase chain reaction, capillary gel electrophoresis
DNA was isolated from tail tips of mice of each RQI strain by the method described by Miller et al. [25]. Markers polymorphic for B6 and C strains were chosen as PCR primers with information from the Mouse Genome Database (Mouse Genome Informatics, The Jackson Laboratory). For QTL mapping 396 microsatellite markers were used. In order to analyze PCR products with our ABI Prism 310 Genetic Analyzer, dye (FAM, TET or HEX)-labeled microsatellite markers were custom-synthesized, fluorescently labeled, and purified by Integrated DNA Technologies, Inc., Coralville, IA. Using a 310 Genetic Analyzer one base size difference could be be detected. The reproducibility of the size identification was good. However, because there are fluctuations, standard DNA products (in our case, PCR products derived from C57BL/6 and BALB/c DNA) were included.
Because in backcrosses B6 females were used, RQI strains contain the X chromosome of the background strain, therefore no markers need to be tested for this chromosome.
Data analysis
Descriptive statistics, eta-squared, Post Hoc Test LSD 0.05 (least significant difference at 0.05 probability level) for alcohol consumption data in RQI strains were analyzed using GLM Univariate ANOVA procedure of SPSS software ver.13 (Table 2).
QTL mapping
For mapping, we used M-estimators, robust alternatives to the sample mean and median for estimating the center of location. The original phenotypic data were processed in the Explore function of SPSS ver.13 to obtain Tukey’s biweight M-Estimator.
Jansen [26] and Zeng [27] proposed similar strategies called composite interval mapping (CIM) by combining interval mapping with multiple regression analysis. If there is no epistasis, the partial regression coefficient of a trait on a marker depends only on those QTLs that are in the interval bracketed by the two neighboring markers and is independent of QTLs located in other intervals [22]. QTL position and effect size was estimated by CIM using Windows QTL Cartographer version 2.5 [28]. We used the standard model Zmapqtl 6 in the CIM procedure with a 10 cM window size and a 2 cM walking speed, forward and backward method of regression with probability of into or out of 0.1. Threshold values of significance for QTLs was established by permutation analysis [29]. This involved reassigning the phenotypic data to RQI lines at random and redoing the Likelihood Ratio Test Statistic analysis. This procedure was repeated 1000 times, for each repetition the global maximum was recorded. Values were ranked in increasing order of significance. The 950th, 900th, and 800th LRS values gave the LRS values for the P = 0.05, P = 0.1, and P = 0.2 levels of significance. Tests were performed for AC345GKD in each population of b5i7 RQI strains. Significant QTLs exceeded the 0.05 genomewide adjusted threshold. QTLs exceeding the 0.2 threshold were considered suggestive.
Strain sets were also analyzed by the MIM option of QTL Cartographer version 2.5 [28]. The MIM method combines QTL mapping analysis with the analysis of genetic architecture of quantitative traits. It utilizes multiple marker intervals simultaneously to construct multiple putative QTLs [30]. The MIM model is based on Cockerham’s model for interpreting genetic parameters and the method of maximum likelihood for estimating genetic parameters. This method fits all QTLs into the model altogether and has the ability for analyzing QTL epistasis and the associated statistical issues. Through a search algorithm, the method can obtain information about the QTLs simultaneously such as number, positions, effects and interaction of the significant QTLs. The search strategy of MIM method is to select the “best” genetic model in the parameter space. Model selection is critically important in data analysis and interpretation. It is well known that the appropriate criteria or stopping rules used for model selection are difficult to decide. In the Windows QTL Cartographer program we used Bayesian Information Criteria (BIC) to control the type I and type II errors at a reasonable level for QTL mapping situation [31]. BIC was defined as: BIC = n*ln(Q*Q) + p*c(n), where n = sample size, Q*Q = residual variance of model, p = regressor (marker) number, c(n) is a penalty function, which can take different forms. Criteria of MIM model selection was based on BIC-M0, where c(n) = ln(n). MIM search walk speed was 1 cM. The model was created by MIM forward search method.
Bioinformatics
Chromosome segments carrying significant or suggestive QTL peaks were investigated for relevant QTLs and genes using http://www.informatics.jax.org and http://omicspace.riken.jp [32]. Range of QTL peak was approximated by inspecting marker genotype patterns in all strains, identifying all donor segments which contained the peak position (cM), and recording the positions of the most proximal and most distal markers on any of the donor segments. This rough approximation gives an estimate of minimum range of donor segments because positions of segment-limiting donor-type markers were used.
Results
The quasi-congenic nature of the B6.C and B6.I RQI strains
Genome-wide genotyping allowed us to obtain a more precise estimate of the donor genome content of each strain. As expected, the average donor genome content in RQI strains varied as a function of the number of backcross–intercross cycles, and the BALB/cJ genome content of the donor strain. On the average (± standard deviation), the B6.Cb4i5, B6.Cb5i7−α, B6.Cb5i7−β, B6.Ib5i7−α, and B6.CI5b7−β strains contained 4.4 ± 1.3%, 2.3 ± 1.6%, 2.4 ± 1.5%, 1.4 ± 1.4%, and 0.99 ± 0.86% of the BALB/cJ genome, respectively. The B6.Cb4i5 (13 strains), B6.Cb5i7 (47 strains), and B6.Ib5i7 (39 strains) mice cover 32.5%, 43.12%, and 20.11% of the BALB/cJ genome, respectively (RQIgbase, release November 20, 2004, www.RQIgenetics.org). Collectively, the 107 RQI strains of RQIgbase cover more than half of the BALB/cJ genome (65.34% of all microsatellite markers have been detected as BALB/cJ-type alleles). B6.Ib5i7 mice carry about half as many CC markers in comparison with B6.Ci5b7 mice because the CXBI donor strain, being a recombinant inbred (RI) strain with C57BL/6ByJ and BALB/cByJ progenitors, is expected to carry only 50% of the BALB/cByJ genome. In comparison, the present study indicated that in the CXBI strain 39.95% of the microsatellite markers showed BALB/cJ genotype. Potentially, the small genomic difference between BALB/cJ and its subline BALB/cByJ could also contribute to the lower proportion of CC markers in CXBI and its B6.I-type descendants.
Genetic variation in oral alcohol self-administration in quasi-congenic B6.Cb5i7 and B6.Ib5i7 RQI strain sets and their progenitors
Table 1 shows the published [12], new, and combined alcohol consumption data for RQI strains and for the progenitors. Descriptive statistics for strain sets is shown in Table 2. New data (strain means for 59 strains) showed highly significant correlation with old data (Pearson r = 0.73 P < 0.001, two-tailed).
Differences in alcohol consumption among the progenitors were highly significant (F 2,312 = 369.65, P < 0.001). Pairwise comparisons with Bonferroni adjustments showed highly significant differences (B6 vs. BALB/cJ, P < 0.001; B6 vs. CXBI (aka CXB-5/ByJ), P < 0.001; BALB/cJ vs. CXBI, P = 0.001). Our results are consistent with previous reports in demonstrating that BALB/cJ and CXBI are alcohol avoiding strains, while C57BL/6ByJ expresses substantial oral alcohol self-administration [1, 12, 33]. Univariate ANOVA across RQI strains indicated that genetic (between-strain) variation of alcohol consumption was highly significant for both the B6.Cb5i7 (F 42,969 = 5.59, P < 0.001) and B6.Ib5i7 (F 34,729 = 7.37, P < 0.001) inbred strain sets. Originally, two-bottle choice test data for B6.Cb4i5 mice were analyzed as alcohol preference phenotype [14]. For better comparability, these data were recalculated and are shown in Tables 1 and 2 as alcohol consumption (g BW −1kg day−1).
QTL mapping
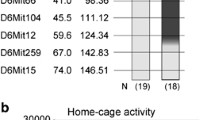
Results of Composite Interval Mapping are shown in Table 3. QTLs are designated as Eac1–6 (ethyl alcohol consumption 1–6). CIM with 43 B6.Cb5i7 RQI strains yielded significant peaks on chr.6 at 36.5 cM (P < 0.001), 42.5 cM (P < 0.001) 62.2 cM (P < 0.01), and 73.5 cM (P < 0.001), and on chr.12 at 51 cM (P < 0.01). Peaks were also detected on chrs. 1, 5, and 15, however these peaks did not reach the significance threshold. Only data on the chr. 15 peak are shown, because this QTL was included in the MIM model.
In the B6.Ii5b7 set of 35 RQI strains two QTLs were found significant at LOD = 2.0 (chr. 12: 21 cM, and chr. 19: 38 cM), but not at empirical threshold determined by 1000 permutations. CIM detected several other nonsignificant peaks on chrs. 1, 3, 4, 12, 13, and 16 with LOD < 2. Only data for the chr. 12 peak are shown, because this QTL was included in the MIM model. In all QTLs shown in Table 3 the B6 alleles increased alcohol consumption. CIM detected several QTLs where the donor allele was associated with increaser effect, however, none of these reached the P = 0.05 level as determined empirically by 1000 permutations. Markers closest to the QTL peak were identified, and the number of strains carrying the donor allele in homozygous condition were counted. Also, for each carrier-strain donor genome contribution (Q%) was calculated (Table 3).
Further analysis of B6.Cb5i7 RQI strains with MIM, using the BIC-M0 relative criterion of QTL Cartographer, yielded a model of three QTLs on chrs. 6, 12, and 15 (genetic R 2 = 0.47). Tests for epistasis indicated no significant additive–additive interactions. MIM with the B6.Ii5b7 set of strains resulted a model of three QTLs positioned on chrs. 8, 12, and 4 (genetic R 2 = 0.5). As with the B6.Ci5b7 strains, no epistatic effect was detected in the B6.Ii5b7 set. Testing the MIM identified QTLs indicated that all QTLs were significant according to the criterion. Results of MIM are summarized in Table 4.
Validity of CIM results (Table 3), and range of peaks were assessed by comparison of microsatellite marker genotype patterns across quasi-congenic RQI strains. In the 43 B6.Cb5i7 RQI strains we assessed validity of QTLs on chrs. 1, 5, 6, 12, and 15. The nonsignificant chr.1 QTL (7.5 cM) was excluded because it was associated with D1Mit167, which also mapped to chr. 5 and chr. 14. As our earlier studies indicated, in the RQI strains D1Mit167 does not co-segregate with chr. 1 markers, but it does with proximal markers of chr. 5 [21]. The nonsignificant chr. 5 QTL (59.01 cM) showed small negative additive effect (−1.29) in the B6.Cb5i7 population, it was also excluded from further consideration. QTLs on chrs. 6, 15, and 12 were retained for bioinformatic analysis (Table 5). In the 35 B6.Ib5i7 RQI strains only QTLs on chrs. 12 and 19 were further investigated (P < 0.2). The chr. 12 QTL was retained for further consideration because the donor allele of the peak marker was represented in several strains (N = 4) and the average donor genome content of these strains was low (1.86%), approaching congenic conditions (Table 5). The chr. 19 QTL was excluded from further consideration because no donor allele was found near the peak position in any of the strains, and the gap between the two markers flanking the peak was relatively large (7 cM).
Discussion
Inspection of marker haplotypes of B6.Cb5i7 strains indicated that the possible minimum range for Eac1 and Eac2 across strains was identical, 33.5–48.2 cM. Because we used a limited number of markers, and the range was defined by the position of the donor marker alleles at the proximal and distal endings of the segments, the real sizes of the segments are not known. In a search of the MGI database (NCBI build 34) at http://www.informatics.jax.org for relevant QTLs in this range we found Eila2, Rear1, Bits2, and Taste5 ([34–36]; see Table 5).
In a recent study on alcohol preference, using BcA Recombinant Congenic strains, Gill and Boyle [7] reported significant correlation between D6Mit178 and alcohol preference in a region containing A/J donor alleles on B6 background between 26.35 and 38.5 cM. However, no chr. 6 QTLs were found for alcohol preference in AcB RC strains, and in the reciprocal AXB/BXA RI strains [7]. Interestingly, the region between the centromer and 33.5 cM contains Neuropeptide Y (Npy, 26 cM) and Ethanol induced locomotion 3 (Etlm3, 30 cM), which were suggested as candidate gene and QTL by Gill and Boyle for their QTL region of 26.35–38.5 cM [7]. In the RQI strains these candidates fell on a relatively large non-genotyped interval between the proximal donor allele (33.5 cM) and the flanking proximal background allele (25.5 cM) of our candidate region, therefore their involvement cannot be excluded.
The 33.5–48.2 cM region is also rich in candidate genes. Using bioinformatic tools, such as the Genome-Phenome Superbrain computational system (GPS; http://omicspace.riken.jp/gps/full.jsp), genes located in the interval were evaluated. Syntenic vomeronasal receptor genes are potential candidates because olfaction may influence alcohol preference. Some other genes may affect neurotransmission (Table 5). In particular, the region contains Grm7, the metabotropic glutamate receptor type 7 gene. Since disturbances in glutamate function have been implicated in the pathophysiology of behavioral disorders, which may underlie some types of addiction [e.g., 37], Grm7 is a promising candidate for further research. Currently, for testing this hypothesis, we are in the process of deriving congenic strains from quasi-congenic RQI strains, which carry segments of the 33.5–48.2 cM region.
In the range of Eac3 (chr.6, 46–65.5 cM) additional relevant QTLs are located ([38–40]; Table 5). It appears that Qui coincides with the taste receptor, type 2 (Tas2r) on distal chromosome 6. Haplotypes at Tas2r locus vary with quinine taste sensitivity in inbred mice, C57BL/6 mice were found quinine sensitive, while DBA/2J mice were quinine insensitive [41]. Haplotype variation suggests that BALB/cJ mice may carry Tas2r alleles associated with quinine insensitivity. However, the specific effects of Tas2r haplotype variation on alcohol consumption remain to be clarified. Taste8 and Taste9 were mapped to 49.5 cM and 63 cM, respectively, suggesting a common polygenic basis for quinine and PROP avoidance in mice [42]. Ethanol conditioned taste aversion 6 [Etohcta6 (63.6)] was mapped close to Taste9 [36]. As Table 5 shows, within an approximately 1 cM region (63–64 cM) we can find three bitterness related loci, Tas2r taste receptor gene members, and an ethanol conditioned taste aversion locus. The accumulation of taste-related QTLs and taste receptor type 2 on a short chromosome region is tempting to hypothesize that Eac3 detected effects of variations in members of Tas2r, however, further evidence is required. Also, a coherent model for the explanation of the complex genotype–alcohol related behavior relationships remains to be developed.
Eac5 on chr. 12 spans an interval of minimum 12 cM (46–58 cM) and has a peak at 51.01 cM. Recently, Gill and Boyle [7] reported a QTL for alcohol intake on chr. 12 at 1.0 cM (range: 1.0–5.0 cM), which does not overlap the range of Eac5. Interestingly, Marq4, the locomotor stimulant effect of another psychostimulant (methamphetamine), was recently mapped to 47 cM on mouse Chromosome 12 which falls in the range of Eac5 [43].
Eac6, a suggestive QTL which did not reach the significance threshold (P < 0.05) by permutation, was mapped to 21 cM (6–29 cM) on chr. 12 in the B6.I set of strains. It is unlikely that the range of Eac6 overlaps the range of the alcohol consumption QTL detected by Gill and Boyle [7] on chr. 12. Psychophysiological processes involved in exploratory behavior (Rear2, [44]), cocaine induced activation (Cocia 1, [45]), dopamine receptor binding (Drb5, [46]), and alcohol acceptance (Aaq2, [47]) may be relevant to choice-based alcohol consumption (Table 5). Syntaxin binding protein 6 is involved in synaptic vesicle-mediated transport. Similarly to Stxbp1, a candidate for an ethanol preference drinking locus on mouse chromosome 2 [5], Stxbp6 may be a candidate gene for Eac6. Interestingly, MIM identified a locus on chr. 4 (64.2 cM), which was positioned near Stx12 (60 cM), a potential contributor to ethanol preference in mice [3, 4], and about 20 cM from Ap3q [9, 48, 49]. However, this locus is a weaker candidate for confirmation studies, because it was not significant in CIM after permutation (P > 0.2).
MIM results supported the CIM results, inasmuch the best model included chr. 6 and chr. 12 QTLs in addition to other QTLs. However, because for purposes of discussion we consider only QTLs with genome-wide significance, QTLs on chromosomes other than chr. 6 and chr. 12 will not be discussed here.
Composite and multiple interval mapping of the whole genome of B6.C and B6.I sets of quasi-congenic RQI strains detected different sets of QTLs for alcohol consumption. This not surprising, because the donor strains were different. BALB/cJ and CXBI provided donor material for B6.C and B6.I, respectively. BALB/cJ and CXBI strains are related because CXBI is one of the CXB recombinant inbred strains. CXBI is expected to carry about 50% BALB/cByJ and 50% C57BL/6By genome on the average. Another differentiating factor is directional selection, which was employed in the development of the B6.C and B6.I strain populations. B6.C mice were developed by five backcross-intercross cycles with 13 generations of concomitant selection for high expression of a brain dopamine-system related trait (mesencephalic tyrosine hydroxylase activity), while B6.I mice were subjected to the same gene introgression manipulation, but selected for extreme low expression of the same phenotype [15, 19, 23]. Therefore, selection pressure in the opposite directions favored the transfer of different sets of donor genes onto the B6 background. Transfer of alcohol preference- and consumption-related genes could be expected either because the mesotelencephalic dopamine system plays a critical role in addiction and alcohol self-administration, or because both donor strains show very low alcohol preference in comparison to the background strain [1, 12, 33], and passenger genes for alcohol-related behaviors could be fixed in the RQI strains.
Several factors may explain the lack of confirmation of our earlier alcohol preference QTL mapping study, which was limited to five chromosomes [12]. First, public database information for the most significant marker (D1Mit167) was insufficient at the time of the analysis because a single proximal position on chr. 1 was assigned for D1Mit167 (chr. 1: 6.5 cM; [50]; http://www.genome.wi.mit.edu/cgi-bin/mouse/index). Our recent results suggested that D1Mit167 in RQI strains cosegregated with markers located on chr. 5 [21], which is consistent with positions for D1Mit167 given by current databases (cf. The T31 Radiation Hybrid Maps Frameworked to TJL BSB / BSS Backcrosses; http://www.jax.org/resources/documents/cmdata/rhmap/maps/). Based on NCBI m34 mouse assembly, D1Mit167 is currently mapped to 3 different Ensemble locations (v.37—Feb 2006): basepairs 3207990–3208105 on chromosome 5, basepairs 17728608–17728725 on chromosome 14, and basepairs 7466556–7466667 on chromosome 1; http://www.ensembl.org/Mus_musculus/markerview? marker = D1Mit173.1. The initial insufficient information on D1Mit167 in public databases also explains why our candidate gene hypothesis for Oprk1 as a gene for alcohol consumption [12], was not tenable, and it is not supported by the current results either. Second, genotyping of additional 14 chromosomes provided new genotype information, and affected mapping statistics. Third, a more advanced mapping methods were applied (CIM, MIM). The increased sample size in some strains slightly changed the phenotypic mean value, however, it is unlikely that sample size changes substantially affected the mapping results.
The Genome–Phenome Superbrain computational system (http://omicspace.riken.jp/; as of March 2006) in a search for alcohol or ethanol related QTLs yielded 40 hits in the MGI QTL database. None of the alcohol preference or consumption QTLs have been assigned to chr. 6, only one related QTL (Aaq2 [47]) was found on chr.12. However, Aaq2 is female specific, while Eac5 and Eac6 were detected in males. Our results, and data by others [6–8], indicate that mapping with strains other than the often used C57BL/6J and DBA/2J progenitors can lead to detection of novel QTLs for alcohol preference and consumption.
In conclusion, we identified new QTLs for alcohol consumption with genome-wide significance on chrs. 6 and 12. Eac1 and Eac6 overlap previously reported QTLs for alcohol preference [7] and alcohol acceptance [47], respectively. Future confirmation studies can take advantage of the quasi-congenic RQI strains by rapidly capturing candidate segments in congenic strains, or by creating segregating populations with low genetic background noise. QTG identification in B6.C RQI strains is now greatly aided by The Mouse Genetic Variation Mapping (Vmap) Initiative by sequencing the genomes in 15 mouse strains, making DNA sequence information available for both progenitors of the RQI strains (http://www.niehs.nih.gov).
References
Mcclearn GE, Rodgers DA (1959) Differences in alcohol preference among inbred strains of mice. Q J Stud Alcohol 20:659–691
Belknap JK, Atkins AL (2001) The replicability of QTLs for murine alcohol preference drinking behavior across eight independent studies. Mamm Genome 12:893–899
Treadwell JA, Pagniello KB, Singh SM (2004) Genetic segregation of brain gene expression identifies retinaldehyde binding protein 1 and syntaxin 12 as potential contributors to ethanol preference in mice. Behav Genet 34:425–439
Treadwell JA (2006) Integrative strategies to identify candidate genes in rodent models of human alcoholism. Genome 49:1–7
Fehr C, Shirley RL, Crabbe JC, Belknap JK, Buck KJ, Phillips TJ (2005) The syntaxin binding protein 1 gene (Stxbp1) is a candidate for an ethanol preference drinking locus on mouse chromosome 2. Alcohol Clin Exp Res 29:708–720
Gill K, Desaulniers N, Desjardins P, Lake K (1998) Alcohol preference in AXB/BXA recombinant inbred mice: gender differences and gender-specific quantitative trait loci. Mamm Genome 9:929–935
Gill K, Boyle AE (2005) Genetic analysis of alcohol intake in recombinant inbred and congenic strains derived from A/J and C57BL/6J progenitors. Mamm Genome 16:319–331
Bachmanov AA, Reed DR, Li X, Li S, Beauchamp GK, Tordoff MG (2002) Voluntary ethanol consumption by mice: genome-wide analysis of quantitative trait loci and their interactions in a C57BL/6ByJ × 129P3/J F2 intercross. Genome Res 12:1257–1268
Grahame NJ, Li TK, Lumeng L (1999) Selective breeding for high and low alcohol preference in mice [In Process Citation]. Behav Genet 29:47–57
Bice PJ, Foroud T, Carr LG, Zhang L, Liu L, Grahame NJ, Lumeng L, Li TK, Belknap JK (2006) Identification of QTLs influencing alcohol preference in the high alcohol preferring (HAP) and low alcohol preferring (LAP) mouse lines. Behav Genet 36:248–260
Mcclearn GE, Wilson JR, Meredith W (1970) The use of isogenic and heterogenic mouse stocks in behavioral research. In: Lindzey G., Thiesse DD (eds) Contributions to behavior genetic analysis: the mouse as a prototype. Appleton-Century-Crofts, New York, pp 3–22
Vadasz C, Saito M, Gyetvai B, Mikics E, Vadasz C 2nd (2000) Scanning of five chromosomes for alcohol consumption loci. Alcohol 22:25–34
Vadasz C, Fleischer A, Lafrancois J, Mao RF (1996) Self-administration of ethanol: towards the location of predisposing polygenes in quasi-congenic animal models. Alcohol 13:617–620
Vadasz C, Saito M, Balla A, Kiraly I, Vadasz C 2nd, Gyetvai B, Mikics E, Pierson D, Brown D, Nelson JC (2000) Mapping of quantitative trait loci for ethanol preference in quasi-congenic strains. Alcohol 20:161–171
Vadasz C (1990) Development of congenic recombinant inbred neurological animal model lines. Mouse Genome 88:16–18
Vadasz C, Sziraki I, Murthy LR, Sasvari-Szekely M, Kabai P, Laszlovszky I, Fleischer A, Juhasz B, Zahorchak R (1994) Transfer of brain dopamine system-specific quantitative trait loci onto a C57BL/6ByJ background. Mamm Genome 5:735–737
Vadasz C, Sziraki I, Sasvari M, Kabai P, Laszlovszky I, Juhasz B, Zahorchak R (1996) Genomic characterization of two introgression strains (B6.Cb4i5) for the analysis of QTLs. Mamm Genome 7:545–548
Abiola O, Angel JM, Avner P, Bachmanov AA, Belknap JK, Bennett B, Blankenhorn EP, Blizard DA, Bolivar V, Brockmann GA, Buck KJ, Bureau JF, Casley WL, Chesler EJ, Cheverud JM, Churchill GA, Cook M, Crabbe JC, Crusio WE, Darvasi A, Haan G De, Dermant P, Doerge RW, Elliot RW, Farber CR, Flaherty L, Flint J, Gershenfeld H, Gibson JP, Gu J, Gu W, Himmelbauer H, Hitzemann R, Hsu HC, Hunter K, Iraqi FF, Jansen RC, Johnson TE, Jones BC, Kempermann G, Lammert F, Lu L, Manly KF, Matthews DB, Medrano JF, Mehrabian M, Mittlemann G, Mock BA, Mogil JS, Montagutelli X, Morahan G, Mountz JD, Nagase H, Nowakowski RS, O’hara BF, Osadchuk AV, Paigen B, Palmer AA, Peirce JL, Pomp D, Rosemann M, Rosen GD, Schalkwyk LC, Seltzer Z, Settle S, Shimomura K, Shou S, Sikela JM, Siracusa LD, Spearow JL, Teuscher C, Threadgill DW, Toth LA, Toye AA, Vadasz C, Zant G Van, Wakeland E, Williams RW, Zhang HG, Zou F (2003) The nature and identification of quantitative trait loci: a community’s view. Nat Rev Genet 4:911–916
Vadasz C, Sziraki I, Sasvari M, Kabai P, Murthy LR, Saito M, Laszlovszky I (1998) Analysis of the mesotelencephalic dopamine system by quantitative-trait locus introgression. Neurochem Res 23:1337–1354
Kovacs KM, Szakall I, O’brien D, Wang R, Vinod KY, Saito M, Simonin F, Kieffer BL, Vadasz C (2005) Decreased oral self-administration of alcohol in kappa-opioid receptor knock-out mice. Alcohol Clin Exp Res 29:730–738
Saito M, Ehringer MA, Toth R, Oros M, Szakall I, Sikela JM, Vadasz C (2003) Variants of kappa-opioid receptor gene and mRNA in alcohol-preferring and alcohol-avoiding mice. Alcohol 29:39–49
Zeng ZB (1993) Theoretical basis for separation of multiple linked gene effects in mapping quantitative trait loci. Proc Natl Acad Sci USA 90:10972–10976
Vadasz C, Sziraki I, Murthy LR, Vadasz I, Badalamenti AF, Kobor G, Lajtha A (1987) Genetic determination of mesencephalic tyrosine hydroxylase activity in the mouse. J Neurogenet 4:241–252
Bailey DW (1971) Recombinant-inbred strains. An aid to finding identity, linkage, and function of histocompatibility and other genes. Transplantation 11:325–327
Miller SA, Dykes DD, Polesky HF (1988) A simple salting out procedure for extracting DNA from human nucleated cells. Nucl Acids Res 16:1215
Jansen RC (1993) Interval mapping of multiple quantitative trait loci. Genetics 135:205–211
Zeng ZB (1994) Precision mapping of quantitative trait loci. Genetics 136:1457–1468
Wang S, Basten CJ, Zeng ZB (2005) Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC
Churchill GA, Doerge RW (1994) Empirical threshold values for quantitative trait mapping. Genetics 138:963–971
Kao CH, Zeng ZB, Teasdale RD (1999) Multiple interval mapping for quantitative trait loci. Genetics 152:1203–1216
Kao CH, Zeng ZB (1997) General formulas for obtaining the MLEs and the asymptotic variance–covariance matrix in mapping quantitative trait loci when using the EM algorithm. Biometrics 53:653–665
Toyoda T, Wada A (2004) Omic space: coordinate-based integration and analysis of genomic phenomic interactions. Bioinformatics 20:1759–1765
Belknap JK, Crabbe JC, Young ER (1993) Voluntary consumption of ethanol in 15 inbred mouse strains. Psychopharmacology 112:503–510
Downing C, Rodd-Henricks KK, Flaherty L, Dudek BC (2003) Genetic analysis of the psychomotor stimulant effect of ethanol. Genes Brain Behav 2:140–151
Le Roy I, Pager J, Roubertoux PL (1999) Genetic dissection of gustatory sensitivity to bitterness (sucrose octaacetate) in mice. C R Acad Sci III 322:831–836
Risinger FO, Cunningham CL (1998) Ethanol-induced conditioned taste aversion in BXD recombinant inbred mice. Alcohol Clin Exp Res 22:1234–1244
Kugaya A, Sanacora G (2005) Beyond monoamines: glutamatergic function in mood disorders. CNS Spectr 10:808–819
Lush IE (1984) The genetics of tasting in mice. III. Quinine. Genet Res 44: 151–160
Singer JB, Hill AE, Nadeau JH, Lander ES (2005) Mapping quantitative trait loci for anxiety in chromosome substitution strains of mice. Genetics 169:855–862
Primo-Parmo SL, Sorenson RC, Teiber J, La Du BN (1996) The human serum paraoxonase/arylesterase gene (PON1) is one member of a multigene family. Genomics 33:498–507
Nelson TM, Munger SD, Boughter JD Jr (2005) Haplotypes at the Tas2r locus on distal chromosome 6 vary with quinine taste sensitivity in inbred mice. BMC Genet 6:32
Harder DB, Whitney G (1998) A common polygenic basis for quinine and PROP avoidance in mice. Chem Senses 23:327–332
Palmer AA, Verbitsky M, Suresh R, Kamens HM, Reed CL, Li N, Burkhart-Kasch S, Mckinnon CS, Belknap JK, Gilliam TC, Phillips TJ (2005) Gene expression differences in mice divergently selected for methamphetamine sensitivity. Mamm Genome 16:291–305
Kelly MA, Low MJ, Phillips TJ, Wakeland EK, Yanagisawa M (2003) The mapping of quantitative trait loci underlying strain differences in locomotor activity between 129S6 and C57BL/6J mice. Mamm Genome 14:692–702
Boyle AE, Gill K (2001) Sensitivity of AXB/BXA recombinant inbred lines of mice to the locomotor activating effects of cocaine: a quantitative trait loci analysis. Pharmacogenetics 11:255–264
Jones BC, Tarantino LM, Rodriguez LA, Reed CL, Mcclearn GE, Plomin R, Erwin VG (1999) Quantitative-trait loci analysis of cocaine-related behaviours and neurochemistry. Pharmacogenetics 9:607–617
Fernandez JR, Vogler GP, Tarantino LM, Vignetti S, Plomin R, Mcclearn GE (1999) Sex-exclusive quantitative trait loci influences in alcohol-related phenotypes. Am J Med Genet 88:647–652
Bachmanov AA, Kiefer SW, Molina JC, Tordoff MG, Duffy VB, Bartoshuk LM, Mennella JA (2003) Chemosensory factors influencing alcohol perception, preferences, and consumption. Alcohol Clin Exp Res 27:220–231
Tarantino LM, Mcclearn GE, Rodriguez LA, Plomin R (1998) Confirmation of quantitative trait loci for alcohol preference in mice. Alcohol Clin Exp Res 22:1099–1105
Copeland NG, Jenkins NA, Gilbert DJ, Eppig JT, Maltais LJ, Miller JC, Dietrich WF, Weaver A, Lincoln SE, Steen RG et al (1993) A genetic linkage map of the mouse: current applications and future prospects. Science 262:57–66
Acknowledgements
Research described in this article was supported by NIH NIAAA grant AA11031. We thank Rui Fen Mao, Janos Piturca, and Ray Wang for excellent assistance in animal care.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Vadasz, C., Saito, M., Gyetvai, B.M. et al. Mapping of QTLs for Oral Alcohol Self-Administration in B6.C and B6.I Quasi-Congenic RQI Strains. Neurochem Res 32, 1099–1112 (2007). https://doi.org/10.1007/s11064-006-9234-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-006-9234-4