
Abstract
Context
Various species of forest trees are commonly used for ornamental purposes and are therefore frequently found in nonforest ecosystems. They constitute an important component of the so-called trees outside forests (TOF). Not much is known, however, about the drivers of TOF spatial distribution either in urbanized or in agricultural landscapes since they are generally absent from forest inventories.
Objective
The present study focused on the spatial distribution of TOF across agricultural landscapes and their potential role in the dispersal of a forest pest insect, the pine processionary moth, Thaumetopoea pityocampa (PPM).
Methods
All the TOF belonging to the genera Pinus, Cedrus and Pseudotsuga were considered as potential hosts and inventoried within a 22 × 22 km study window. We fitted a nonstationary Poisson process to the empirical data and used the distance to the nearest building as a covariate.
Results
Both empirical and simulated data indicated that TOF associated to human artifacts/urbanized areas constituted the main source of landscape connectivity for the PPM in the open fields under study. Because they do not account for TOF, forest inventories dramatically underestimate landscape connectivity and provide an erroneous picture of the PPM habitat distribution.
Conclusions
We conclude that TOF, especially the ornamental component, must be taken into account when it comes to understanding forest insect landscape dynamics or genetics. The omnipresence of TOF also suggests a potentially huge role in pest dispersal and invasive species expansion.
Similar content being viewed by others


Introduction
Habitat degradation and fragmentation is considered to be one of the primary threats to biodiversity worldwide (Balmford et al. 2003). By disrupting dispersal, fragmentation alters a critical component of metapopulation dynamics and population persistence which strongly affects biological diversity (Wiens 2001; Hanski 2011). For that reason, species conservation issues are often viewed from a landscape perspective and this is one of the reasons why landscape ecology has become increasingly relevant for management and conservation of biological diversity (Gutzwiller 2002; Lindenmayer and Hobbs 2007; Wiens 2009; Hanski 2011). One widespread management option consists of developing green infrastructures and corridors to help connecting rural and urban or periurban territories in order to limit landscape fragmentation and facilitate species dispersal (Benedict and McMahon 2006; Clergeau and Blanc 2013). Trees and shrubs are major elements of these so-called green belts. Trees outside forests (TOF) are increasingly recognized as a prominent feature of either agricultural lands or built-up areas (Bellefontaine et al. 2001; Gutzwiller 2002; Konijnendijk et al. 2005; de Foresta et al. 2013). TOF are “trees on land not defined as forest and other wooded land,” a definition that is dependent on how forests and woodlands are defined themselves (Kleinn 2000; de Foresta et al. 2013). In agricultural landscapes, TOF are often referred to as scattered, isolated or paddock trees (Manning et al. 2006; Gibbons et al. 2008). In cities and towns, TOF are street and roadside trees, trees planted in parks, gardens and private yards (Tyrvainen et al. 2005; Wu 2007). TOF provide various ecosystem services such as control over soil erosion, nutrient and water cycling, biodiversity conservation or pest control (Plieninger et al. 2004; Lumsden and Bennett 2005; Manning et al. 2006). Urban woods and isolated trees also provide important social, aesthetic, and economic benefits (Tyrvainen et al. 2005; McDonnell et al. 2009).
A proportion of TOF are ornamental or amenity trees grown for decorative purposes in gardens and landscape design projects. These trees are native, naturalized or recently introduced exotic species. In recent decades, the exotic component of ornamental planting has increased as the international horticultural industry has developed and changed toward a global market. This evolution is associated with important consequences on the risk of emerging plant diseases (Dehnen-Schmutz et al. 2010; Ööpik et al. 2013). However, while TOF benefits and services are increasingly acknowledged, it is worth noting that they also provide ecosystem disservices such as the production of allergens, potential damage of infrastructure and the introduction of new pests and pathogens (Roy et al. 2012; Bigsby et al. 2014; Dobbs et al. 2014).
Despite their ubiquity, little is known about ornamental TOF spatial distribution and the ecological and/or socio-economical processes that drive it at the landscape scale. Ornamental species are species reported as being frequently planted near houses and buildings (Pauleit et al. 2005) so that their spatial distribution is likely to be linked to human settlements but, to our knowledge, quantitative data describing such association are lacking. In addition, TOF (either ornamental or not) are generally absent from national forest inventories (Kleinn 2000; de Foresta et al. 2013) and this lack of data precludes a proper assessment of their contribution to landscape connectivity with regards to associated species.
In the present study, we tackle the question of TOF contribution to landscape connectivity with regards to a forest defoliating insect, the pine processionary moth (PPM) (Thaumetopoea pityocampa). The PPM is an important forest pest in the Mediterranean regions and its geographical range is currently expanding in relation to climate change (Roques 2015). Little is known about the landscape scale spread of the PPM, particularly in non-forest areas. We worked in large open-field agricultural landscapes of the Centre region of France where available forest inventories indicate a very low coverage of tree species suitable for our model insect (IFN 2010). In this recently colonized area, we thus expected a high degree of habitat fragmentation with regards to the PPM host species (referred to as host trees outside forests, HTOF) coverage. Using systematic inventory, we georeferenced all isolated PPM host trees either located on public or private lands within a large sampling window. We examined the possible relationship between human artifacts and HTOF distribution using data from the French institute IGN (Institut National de l’Information Géographique et Forestière) using point process modeling. Because exhaustive inventories are tedious and costly, spatial simulation may complete or enhance field data (O’Sullivan and Perry 2013). We explored how the resulting model could be used to simulate HTOF distribution over a large region of open-field where no HTOF inventory was done. Finally, we used landscape metrics to quantify HTOF contribution to landscape structure and particularly fragmentation with regard to the PPM.
Methods
Model species and study site
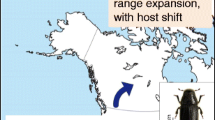
The PPM is the most important pine and cedar defoliator in southern Europe and northern Africa (Cielsa 2011). Pinus nigra, one of its preferred hosts, has been used for large-scale afforestation and ornamental plantations throughout France. The distribution of PPM is controlled by the minimum winter temperatures (Huchon and Demolin 1970), and the species range is currently expanding northward and in elevation due to climate change (Battisti et al. 2005; Robinet et al. 2007). Since the early 90s, the PPM’s range has expanded towards north of France without being noticeably affected by the presence of large wide open-field areas such as the Beauce region (Roques et al. 2015a) despite limited female dispersal ability (Robinet et al. 2012) (Fig. 1). PPM dispersal in nonforest landscapes may strongly depend on scattered host trees although large-scale field data are lacking (Roques et al. 2015b). In order to unravel the potential role of HTOF as elements of landscape connectivity for the dispersal of PPM, we designed a field survey just north of the open-field region of Beauce in the centre of France (Fig. 1). This area is amongst Europe’s most productive cereal regions and thus appeared to be a sensible choice given the scarcity of coniferous forest cover, the typical habitat of the PPM. Figure 1 shows the position of the Centre region of France (Fig. 1 left) and the expansion front of the PPM as recorded in the winters of 2005–2006 and 2010–2011.

Study site. a Centre Region, France. Shaded area indicates the PPM range in winter 2005–2006, dark area represents the expansion of the PPM between 2006 and 2010. b Survey plot (22 × 22 km) where an exhaustive inventory of all trees outside forests belonging to the genera Pinus, Cedrus and Pseudotsuga was carried out (referred to as HTOF for host trees outside forests). c The ecoregion of “Beauce” where the HTOF spatial distribution was simulated
Field sampling
An exhaustive inventory of PPM HTOF was carried out in a 22 × 22 km = 484 km2 area located in the north of the Centre region of France (Fig. 1). All trees belonging to the genera Pinus, Cedrus and Pseudotsuga were considered as potential hosts and are hereafter referred to as HTOF for host trees outside forests. The study site was located in the southern part of an ecoregion referred to as the Drouais-Thymerais region (Conseil régional Centre 2009) where the landscape mostly consisted of wide arable lands (cereal). All motorable roads and tracks were visited during autumn and winter 2009–2010. This period of the year was preferred because sighting and identification of PPM host trees (evergreen coniferous species) is easier when deciduous trees have lost their leaves (Rousselet et al. 2013). Every individual or small group of host trees (single trees, linear groups of trees, small woodlands) was observed by eye, and with binoculars when necessary, from the road and public land. The geographic coordinates were recorded using a GarminTM GPS12 (or the location was mapped onto a georeferenced aerial photo using PhotoExploreur Bayo© in case of distant observation). In so doing, we obtained an exhaustive inventory of locations of PPM HTOF. The inventory data are available in Rousselet et al. (2015).
Forest inventories and land use information available in the study area
The French institute IGN (Institut National de l’information Géographique et Forestière http://www.ign.fr/) provides a national forest inventory (referred to as IFN for “Institut National Forestier” in French) of all the stands with an area >2.25 ha, i.e. approximately corresponding to a resolution of 150 × 150 m pixels (http://inventaire-forestier.ign.fr/carto/carto/afficherCarto/28) (Fig. 2). This institute also develops and maintains a database called BD Topo®IGN http://professionnels.ign.fr/bdtopo, which is a vector database documenting both geographical and administrative entities at a 1:25,000 scale over the whole French territory.

Spatial distribution of trees outside forests, forest cover and the distance to the nearest building in a 22 × 22 km open-field agricultural landscape. a Distribution of the 3834 TOF belonging to the genera Pinus, Cedrus and Pseudotsuga (crosses). b Binary mapping of the PPM host trees (Pinus, Cedrus and Pseudotsuga) as provided by the IFN database. c Map of the distance to the nearest building in the survey plot. The farther from a building a pixel, the larger this distance (data from BD Topo®IGN, édition 2013 http://professionnels.ign.fr/bdtopo). d Barplot depicting the frequency distribution of the observed distance to the nearest building for 3834 inventoried TOF
Modelling HTOF spatial distribution
We modeled the HTOF spatial distribution in order to explore the link between tree spatial pattern and the distribution of buildings over the study area. The resulting model provided a convenient way of simulating HTOF distribution over unsampled areas. HTOF were modeled by fitting an inhomogeneous Poisson process, i.e. nonstationary Poisson process (Diggle 2003), upon the observed tree location within the experimental plot (Fig. 2a). The inhomogeneous Poisson process allows the introduction of covariates in the analysis of the observed point pattern by considering an intensity function λ(x). In our case, the model was λ(x) = exp(α + β1z(x) + β2z(x)2 + β3z(x)3 + β4z(x)4 + β5z(x)5), where λ(x) denotes the intensity function at location x and z(x) stands for the covariate at location x. We used the log of 1+ the distance to the nearest object corresponding to any sort of building as the covariate. The covariate values were computed from the data of the BDO TOPO 2.1 using the SIG software Quantum GIS (Quantum GIS Development Team 2014) (Fig. 2c).
Our rationale behind the use of a Poisson process was to mimic citizen behavior with regard to ornamental tree plantations. People plant ornamental species in the close vicinity of their house, for example in their yards or in front of their property (Pauleit et al. 2005). Public land managers plant on lands distributed all over the built-up areas, especially along public thoroughfares serving the buildings (Konijnendijk et al. 2005). We account for this behavior by using a nonstationary model where the nature of the covariate directly conveys the effect of building closeness. On the other hand, not all private or public owners would plant ornamental trees suitable for the PPM and this random aspect is accounted for through the Poisson process itself.
The fitted model was evaluated by performing residual analysis (Baddeley and Turner 2006) in which the point process is transformed into a Poisson process with uniform intensity on a given interval. The deviation of the transformed point process from a homogenous Poisson process indicates a possible model misfit. We used the Stoyan-Grabarnik diagnostic tool “exponential energy marks” (Stoyan and Grabarnik 1991) based on the reciprocal of the conditional intensity. Each point is given a weight depending on the conditional intensity of the fitted model and the sum of the weights for all the points divided by the area of the study plot has an expected value of 1 (Stoyan and Grabarnik 1991; Baddeley et al. 2005). Model fitting was done using a maximum pseudolikelihood fitting algorithm implemented in the R statistical software (R Core Team 2014) within the package spatstat (Baddeley and Turner 2005, 2006). Once the model linking HTOF to the covariate was fitted, it was used to perform stochastic simulations of the point process at hand within the sampling window (Møller and Waagepetersen 2004; Baddeley and Turner 2005). A total of 1000 simulations were used to examine the model performance with regards to a set of selected landscape metrics intended to characterize landscape features. The simulated point patterns were rasterized with a resolution equal to that of the IFN database i.e. pixels of 2.25 ha (150 × 150 m, Fig. 2) using the R package raster (Hijmans 2014). The resulting binary maps were used to compute the PLAND, PD and ED metrics (described below). The results were compared to the values computed for the empirical data set. Similarly, we simulated the HTOF distribution on the scale of the Beauce region (Fig. 1c).
Landscape configuration from the PPM viewpoint
We investigated the structure of landscapes derived from (i) the IFN raster database (ii) the HTOF database, and (iii) the IFN + HTOF database by means of 3 key landscape metrics: percentage of landscape, patch density and edge density (Turner et al. 2001). The percentage of landscape (PLAND) was used to quantify the proportional abundance of the patches that corresponded to a suitable habitat for the PPM, i.e. corresponding to pixels where at least one individual host tree was present. Patch density (PD) described landscape structure by quantifying the degree of fragmentation of PPM host distribution. In addition, we used the edge density metric (ED) to quantify the shape and complexity of the habitat patches (Turner et al. 2001). This index appeared of interest given that the PPM is known to preferentially colonize forest stand edges (Samalens and Rossi 2011). Computations were done under R with the package SDMTools (VanDerWal et al. 2014).
Results
Descriptive statistics in the inventory plot
According to the IGN, the exhaustively sampled area mainly consisted of an intensively managed agricultural landscape with only 12 % of forest and less than 1 % of pure or mixed stands with coniferous trees. We recorded a total of 3834 individuals or small groups of PPM HTOF in the survey window (Fig. 2a), which corresponded to a density of 7.9 occurrences per 100 ha. Figure 2 shows the observed HTOF (black crosses) (Fig. 2a) and the pixels (150 × 150 m) where the IFN inventory reported the presence of host trees (Fig. 2b). Of the 3834 HTOF, 285 (7.4 %) were separated from the nearest building by a distance <10 m; 1113 (29 %) were separated by a distance ranging from 10 to 20 m; and 627 (16.35 %) were separated by a distance ranging between 20 and 30 m (Fig. 2c, d). The spatial configuration of HTOF is depicted in Fig. 3. The average separating distance between trees and their closest neighbors was <50 m for more than 70 % HTOF (Fig. 3a). Figure 3b shows experimental data illustrating the flight abilities of female moths in the laboratory (data from Robinet et al. 2012). From this barplot, it can be seen that more than 54 % of female moths fly between 0 and 1 km; 16.21 % fly a distance between 1 and 2 km, while this proportion is 13.51 % for distances ranging from 2 to 3 km. These flying abilities are compatible with the inter-HTOF distances reported above.

Spatial configuration of HTOF. a Frequency distribution of the distance between each HTOF and its closest neighbor. b Frequency distribution of the flight capacities of female PPM (data from Robinet et al. 2012)
The percentage of landscape associated with PPM potential hosts appeared to be very limited when estimated on the basis of the IFN data (1.5 %), while it was more than 10 times higher when measured from observed HTOF (12.4 %) (Table 1). In addition, both patch density and edge density were strongly underestimated when measured from the forest inventory alone (Table 1).
Inhomogeneous Poisson model and landscape distribution of HTOF
The coefficients of the fifth degree model were α = −9.40; β1 = −18.77; β2 = 16.30; β3 = −4.99; β4 = 0.64; β5 = −0.029. The Stoyan-Grabarnik diagnostic tool divided by the plot area was 1.005526, which revealed a good fit. The values of the PLAND, PD and ED metrics for the empirical and simulated data set are given in Table 1. On average, the landscape patterns resulting from the model slightly overestimated the percentage of landscape (PLAND) where HTOF are present (16.8 %) as compared to the observed value (12.4 %). Similarly, patch density and edge density were slightly overestimated (Table 1).
Simulations over the Beauce ecoregion
Figure 4a shows the binary maps of the HTOF distribution over the Beauce region available from the IFN (see also Fig. 1). Figure 4b gives an example of a simulated HTOF distribution over the same region. Table 2 gives the landscape metric values computed from the IFN data, the simulated HTOF distributions and the sum of these data sets. The coverage of trees suitable for the PPM given in traditional forest inventory (IFN) appeared to be markedly lower than the value estimated from the simulated distributions. Similarly, both the patch and the edge density were higher according to the model simulations.

Binary mapping of the regional distribution of host trees suitable for the pine processionary moth (Thaumetopoea pityocampa) in the ecoregion of “Beauce” (see Fig. 1). a Data available for the French National Forest Inventory (IFN, currently grouped with the Institut National de l’Information Géographique et Forestière http//:inventaire-forestier.ign.fr). b Example of simulated HTOF distribution. The raster resolution is of 2.25 ha, i.e. 150 × 150 m pixels in both maps
Discussion
Spatial distribution of TOF and the prominence of ornamental trees
The results reported here unambiguously indicate that most of the tree formations suitable for the PPM in our inventory area are TOF. While native broadleaved TOF located within or bordering the fields are rare and considered relictual in the intensively managed agricultural landscapes (Plieninger 2012), the PPM host tree species were not rare and appeared to be scattered across the landscape. Interestingly, no pine, cedar or Douglas fir (i.e. the host species of the PPM referred to as HTOF in this study) is native to the survey region even if natural regeneration sometimes occurs, especially in unused lands. The trees considered here are planted near human artifacts such as buildings (public land and residential, commercial and industrial areas) for decorative purposes. This leads to a strong spatial relationship between trees and buildings. As a consequence, 70 % of the inventoried tree formations were found in 4.5 % of the territory defined as urbanized areas in the BD CARTO database (BD Topo®IGN 2013). If HTOF are locally aggregated in the vicinity of buildings, the villages are evenly distributed in the study plot (Fig. 2) and this leads to a regular distribution tree patches throughout the landscape with an average inter-tree distance well below the flight capacity of the PPM. As a consequence, the landscape functional connectivity clearly appears high enough to allow the expansion of the moth across that open-field agricultural landscape. Such an even host tree distribution was previously unsuspected because our knowledge of coniferous species distribution in the region relied solely on the available forest inventory databases. Beyond the case of the PPM and its host trees, our results suggest that accounting for TOF in forest inventories would be important for many ecological studies especially when dealing with metapopulations or metacommunities (Hanski and Gaggiotti 2004; Holyoak et al. 2005).
The present work focuses on a set of tree species mainly used for ornamental purposes. As such, the drivers of their dynamics fall largely within social forces such as consumers’ choice and market fluctuations. In 2010, for instance, 475.2 million outside plants were purchased by the French people (FranceAgriMer 2011). Of these plants, 52.8 million were trees and shrubs including 15.1 million small fruit bushes, 4.2 million fruit trees and 33.5 million ornamental trees and shrubs. In the Beauce region, these ubiquitous trees and shrubs are planted in various types of environment depending on the buyer’s location and cause lasting changes in local vegetation. It should be noted that some forest tree species are commonly used as ornamental trees, especially Pinus nigra, the preferred host of the PPM. In other words, citizens and local communities have a strong impact on host tree availability for the PPM through the choice of ornamental species. Addressing this point is beyond the scope of the present paper but it is worth noting that understanding the drivers of ornamental TOF dynamics would necessarily involve economic and social sciences.
Point process models for TOF
The model satisfactorily fitted the data, although simulated point patterns proved to slightly underestimate tree dispersion as indicated by the overestimation of PLAND, patch and edge density. A possible improvement would be to deepen the level of details taken into account when considering urban landscapes, for instance by distinguishing different types of buildings with regard to human occupation and function (farms, houses, public buildings…). In agricultural open-field landscapes like the Beauce region considered here, we suspect that ornamental trees are often planted in residential areas and in villages, but not at the same level in farmhouses and outbuildings. The pattern of ownership in urbanized areas may strongly affect the selection of ornamental species, their density and spatial arrangement in cities (Pham et al. 2011). Again, this goes beyond the scope of the present paper and a proper consideration of these aspects would require broadening the scope of our approach by including socioeconomic components.
The strength of point process modeling is that it allows simulating point patterns corresponding to the model fitted to observed data over an unsampled region (Diggle 2003; O’Sullivan and Perry 2013). This approach could be used to draw maps of TOF at large geographical scales where exhaustive data might be impossible to collect for obvious practical reasons. One very important limit of such an approach is that a model can only be used for simulating data in a landscape of similar nature and assuming that TOF generating processes at work are identical. Given this intrinsic limitation, simulations may provide useful—and cheap—information. TOF are generally assessed using stratified sampling and remote sensing-based approaches (Holmgren et al. 1994; Corona and Fattorini 2006; Tokola 2006; Lam et al. 2011; Baffetta et al. 2011; Lister et al. 2012; Meneguzzo et al. 2013; Westinga et al. 2013). A possible area of research would be to explore how point process models could be used to optimize field sampling and/or how point process simulation could be used to map TOF and complete the data available in forest inventories.
Landscape connectivity, pest spatial dynamics and management
Data from the available forest inventory suggested that the Beauce ecoregion offers poor habitat resources for the PPM due to a very limited forest cover, especially in terms of conifers (IFN 2010) (Fig. 2a, b). Despite their very low density in comparison with forest stands, ornamental TOF associated with built-up areas provide numerous potential relays for the PPM females during the range expansion. Percentage of landscape, patch density and edge density all indicate that landscape permeability is strongly underestimated when only considering forest stands. Ornamental TOF therefore constitute an unsurprisingly, although rarely explicitly considered, important network corresponding to low tree densities. The resulting landscape connectivity with regards to associated invertebrate or pathogen species is thus of high functional importance. In the case of the PPM, given its flight capacities (Fig. 3), it appears that there is no barrier to spread of that forest insect in large open-field agricultural regions. More generally, models for pest dynamics may benefit from an explicit account for TOF as habitat resources and dispersal corridors. For example, taking TOF into consideration in landscape genetics may lead to a refined and more detailed perception of landscape constraints towards gene flows, hence a better understanding of species spatial ecology and spatial genetic variation (Kerdelhué et al. 2015; Roques et al. 2015b).
Urban forests harbor a large number of insect pests also present in planted or natural forests, orchards or forest nurseries (Tello et al. 2005). In addition to native and naturalized pest species, urban trees are exposed to an increasing number of exotic pests and pathogens due to international trade (Smith et al. 2007; Tubby and Webber 2010; Kowarik 2011). Before spreading across large areas, most successful invasive species are firstly introduced in urbanized areas that act as hubs of international transport and then constitute major gateways by which species spread (Zippeter 2008; Colunga-Garcia et al. 2010). Urban areas are also hot spots of plant species richness, either for native or introduced species (Kowarik 2011) which, in turn, could be a facilitating factor of establishment success of insect pests. For example Colunga-Garcia et al. (2010) showed that the risk of invasion increased near to urbanized areas. Spread of invasive species can also be facilitated by green infrastructures that are intended to improve urban–rural gradients with regards to biodiversity management. So far, such ecosystem disservice has received less attention than beneficial effects (but see Säumel and Kowarik 2010). In the case of the PPM, P. nigra is amongst its preferred pine hosts and has massively been used as an ornamental tree in private and public lands. An obvious management option would be to discourage massive planting of P. nigra in areas frequented by public at risk like schools. Quantifying the contribution of different types of ownership (private yards, industrial and commercial areas managed by landscape gardeners, street trees or public green spaces managed by local public authorities) to the dynamics of ornamental tree plantings would help develop a better understanding of the dynamics of green infrastructures and their role in the spatial ecology of pests.
References
Baddeley A, Turner R (2005) spatstat: an R package for analyzing spatial point patterns. J Stat Soft 12:1–42
Baddeley A, Turner R (2006) Modelling spatial point patterns in R. In: Baddeley A, Gregori P, Mateu J et al (eds) Case studies in spatial point process modeling. Springer-Verlag, New York, pp 23–74
Baddeley A, Turner R, Møller J, Hazelton M (2005) Residual analysis for spatial point processes (with discussion). J R Stat Soc B 67:617–666
Baffetta F, Corona P, Fattorini L (2011) Assessing the attributes of scattered trees outside the forest by a multi-phase sampling strategy. Forestry 84:315–325
Balmford A, Green RE, Jenkins M (2003) Measuring the changing state of nature. Trends Ecol Evol 18:326–330
Battisti A, Stastny M, Netherer S, Robinet C, Schopf A, Roques A, Larsson S (2005) Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol Appl 15:2084–2096
BD Topo®IGN (2013) BD Topo®IGN. http://professionnels.ign.fr/bdtopo. Accessed 15 May 2015
Bellefontaine R, Petit S, Pain-Orcet M, Deleporte P, Bertault J (2001) Les arbres hors forêt: vers une meilleure prise en compte. Cahiers FAO conservation, 35. FAO, Rome
Benedict MA, McMahon ET (2006) Green infrastructure: linking landscapes and communities. Island Press, Washington
Bigsby KM, Ambrose MJ, Tobin PC, Sills EO (2014) The cost of gypsy moth sex in the city. Urban For Urban Green 13:459–468
Cielsa WM (2011) Forest entomology. Wiley-Blackwell, Chichester
Clergeau P, Blanc N (2013) Trames vertes urbaines. Le Moniteur, Paris
Colunga-Garcia M, Magarey RA, Haack RA, Gage SH, Qi J (2010) Enhancing early detection of exotic pests in agricultural and forest ecosystems using an urban-gradient framework. Ecol Appl 20:303–310
Conseil régional Centre (2009) Cartographie du réseau écologique sur le territoire de la région Centre – BIOTOPE, Orléans
Corona P, Fattorini L (2006) The assessment of tree row attributes by stratified two-stage sampling. Eur J For Res 125:57–66
de Foresta H, Somarriba E, Temu A, Boulanger D, Feuilly H, Gauthier M (2013) Towards the assessment of trees outside forests forest. Resources Assessment Working Paper 183. FAO, Rome
Dehnen-Schmutz K, Holdenrieder O, Jeger MJ, Pautasso M (2010) Structural change in the international horticultural industry: some implications for plant health. Sci Hortic 125:1–15
Diggle PJ (2003) Statistical analysis of spatial point patterns. Oxford University Press, Oxford
Dobbs C, Kendal D, Nitschke CR (2014) Multiple ecosystem services and disservices of the urban forest establishing their connections with landscape structure and sociodemographics. Ecol Indicat 43:44–55
FranceAgriMer (2011) Végétaux d’extérieur : Achat des français en 2010. http://www.franceagrimer.fr. Accessed 15 May 2015
Gibbons P, Lindenmayer DB, Fischer J, Manning AD, Weinberg A, Seddon J, Ryan P, Barrett G (2008) The future of scattered trees in agricultural landscapes. Conserv Biol 22:1309–1319
Gutzwiller K (ed) (2002) Applying landscape ecology in biological conservation. Springer-Verlag, New York
Hanski I (2011) Habitat loss, the dynamics of biodiversity, and a perspective on conservation. Ambio 40:248–255
Hanski I, Gaggiotti OE (eds) (2004) Ecology, genetics, and evolution of metapopulations. Elsevier, Amsterdam
Hijmans RJ (2014) Raster: geographic data analysis and modeling. R package version 2.1-49. http://CRAN.R-project.org/package=raster. Accessed 15 May 2015
Holmgren P, Masakha EJ, Sjöholm H (1994) Not all African land is being degraded: a recent survey of trees on farms in Kenya reveals rapidly increasing forest resources. Ambio 23:390–395
Holyoak M, Leibold MA, Holt RD (2005) Metacommunities: spatial dynamics and ecological communities. University of Chicago Press, Chicago
Huchon H, Demolin G (1970) La bioécologie de la processionnaire du pin: dispersion potentielle-dispersion actuelle. Rev For Fr 22:220–233
IFN (2010) La forêt françaises, les résultats pour la région Centre. http://inventaire-forestier.ign.fr/spip/IMG/pdf/IFN_Publi_2010_Centre.pdf. Accessed 15 May 2015
Kerdelhué C, Battisti A, Burban C, Branco M, Cassel-Lundhagen A, İpekdal K, Larsson S, Lopez-Vaamonde C, Magnoux E, Mateus E, Mendel Z, Negrisolo E, Paiva M-R, Pivotto I, Rocha S, Ronnås C, Roques A, Rossi J-P, Rousselet J, Salvato P, Santos H, Simonato M, Zane L (2015) Genetic diversity and structure at different spatial scales in the processionary moths. In: Roques A (ed) Processionary moths and climate change: an update. Springer, Dordrecht, pp 163–226
Kleinn C (2000) On large-area inventory and assessment of trees outside forests. UNASYLVA FAO 51:3–10
Konijnendijk CC, Nilsson K, Randrup TB (eds) (2005) Urban forests and trees: a reference book. Springer, Berlin
Kowarik I (2011) Novel urban ecosystems, biodiversity, and conservation. Environ Pollut 159:1974–1983
Lam TY, Kleinn C, Coenradie B (2011) Double sampling for stratification for the monitoring of sparse tree populations: the example of Populus euphratica Oliv. forests at the lower reaches of Tarim River, Southern Xinjiang, China. Environ Monit Assess 175:45–61
Lindenmayer DB, Hobbs RJ (eds) (2007) Managing and designing landscapes for conservation: moving from perspectives to principles. Blackwell, Malden
Lister AJ, Scott CT, Rasmussen S (2012) Inventory methods for trees in nonforest areas in the great plains states. Environ Monit Assess 184:2465–2474
Lumsden LF, Bennett AF (2005) Scattered trees in rural landscapes: foraging habitat for insectivorous bats in south-eastern Australia. Biol Conserv 122:205–222
Manning AD, Fischer J, Lindenmayer DB (2006) Scattered trees are keystone structures—implications for conservation. Biol Conserv 132:311–321
McDonnell MJ, Hahs AK, Breuste JH (2009) Ecology of cities and towns: a comparative approach. Cambridge University Press, Cambridge
Meneguzzo DM, Liknes GC, Nelson MD (2013) Mapping trees outside forests using high-resolution aerial imagery: a comparison of pixel- and object-based classification approaches. Environ Monit Assess 185:6261–6275
Møller J, Waagepetersen RP (2004) Statistical inference and simulation for spatial point processes. Chapman & Hall/CRC, Boca Raton
Ööpik M, Bunce RGHB, Tischler M (2013) Horticultural markets promote alien species invasions: an Estonian case study of herbaceous perennials. NeoBiota 17:19–37
O’Sullivan D, Perry GLW (2013) Spatial simulation. Exploring pattern and process. Wiley-Blackwell, Chichester
Pauleit S, Jones N, Nyhuus S, Pirnat J, Salbitano F (2005) Urban forests resources in european cities. In: Konijnendijk CC, Nilsson K, Randrup TB (eds) Urban forests and trees: a reference book. Springer, Berlin, pp 49–80
Pham T-T-H, Apparicio P, Séguin A-M, Gagnon M (2011) Mapping the greenscape and environmental equity in Montreal: an application of remote sensing and GIS. In: Caquard S, Vaughan L, Cartwright W (eds) Mapping environmental issues in the city: arts and cartography. Springer-Verlag, Berlin
Plieninger T (2012) Monitoring directions and rates of change in trees outside forests through multitemporal analysis of map sequences. Appl Geogr 32:566–576
Plieninger T, Pulido FJ, Schaich H (2004) Effects of land-use and landscape structure on holm oak recruitment and regeneration at farm level in Quercus ilex L. dehesas. J Arid Environ 57:345–364
Quantum GIS Development Team (2014) Quantum GIS geographic information system. Open source geospatial foundation project. http://qgis.osgeo.org. Accessed 15 May 2015
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0. http://www.R-project.org/. Accessed 15 May 2015
Robinet C, Baier P, Pennerstorfer J, Schopf A, Roques A (2007) Modelling the effects of climate change on the potential feeding activity of Thaumetopoea pityocampa (Den. & Schiff.) (Lep., Notodontidae) in France. Global Ecol Biogeogr 16:460–471
Robinet C, Imbert C-E, Rousselet J, Sauvard D, Garcia J, Goussard F, Roques A (2012) Human-mediated long-distance jumps of the pine processionary moth in Europe. Biol Invasions 14:1557–1569
Roques A (ed) (2015) Processionary moths and climate change: an update. Springer, Dordrecht
Roques A, Rousselet J, Avcı M, Avtzis D, Basso A, Battisti A, Ben Jamaa M, Bensidi A, Berardi L, Berretima W, Branco M, Chakali G, Çota E, Dautbašić M, Delb H, El Alaoui El Fels M, El Mercht S, El Mokhefi M, Forster B, Garcia J, Georgiev G, Glavendekić M, Goussard F, Halbig P, Henke L, Hernańdez R, Hódar J, İpekdal K, Jurc M, Klimetzek D, Laparie M, Larsson S, Mateus E, Matošević D, Meier F, Mendel Z, Meurisse N, Mihajlović L, Mirchev P, Nasceski S, Nussbaumer C, Paiva M-R, Papazova I, Pino J, Podlesnik J, Poirot J, Protasov A, Rahim N, Sańchez Peña G, Santos H, Sauvard D, Schopf A, Simonato M, Tsankov G, Wagenhoff E, Yart A, Zamora R, Zamoum M, Robinet C (2015a) Climate warming and past and present distribution of the processionary moths (Thaumetopoea spp.) in Europe, Asia Minor and North Africa. In: Roques A (ed) Processionary moths and climate change: an update. Springer, Dordrecht, pp 81–161
Roques L, Rossi J-P, Berestycki H, Rousselet J, Garnier J, Roquejoffre J-M, Rossi L, Soubeyrand S, Robinet C (2015b) Modeling the spatio-temporal dynamics of the pine processionary moth. In: Roques A (ed) Processionary moths and climate change: an update. Springer, Dordrecht, pp 227–263
Rousselet J, Imbert C-E, Dekri A, Garcia J, Goussard F, Vincent B, Denux O, Robinet C, Dorkeld F, Roques A, Rossi J-P (2013) Assessing species distribution using Google Street View: a pilot study with the pine processionary moth. PLoS One 8:e74918
Rousselet J, Garcia J, Roques A, Rossi J-P (2015) An exhaustive inventory of coniferous trees in an agricultural landscape. Biodivers Data J 3:e4660
Roy S, Byrne J, Pickering C (2012) A systematic quantitative review of urban tree benefits, costs, and assessment methods across cities in different climatic zones. Urban For Urban Green 11:351–363
Samalens J-C, Rossi J-P (2011) Does landscape composition alter the spatiotemporal distribution of the pine processionary moth in a pine plantation forest? Popul Ecol 53:287–296
Säumel I, Kowarik I (2010) Urban rivers as dispersal corridors for primarily wind-dispersed invasive tree species. Landsc Urban Plan 94:244–249
Smith RM, Baker RHA, Malumphy CP, Hockland S, Hammon RP, Ostojá-Starzewski JC, Collins DW (2007) Recent non-native invertebrate plant pest establishments in Great Britain: origins, pathways, and trends. Agric For Entomol 9:307–326
Stoyan D, Grabarnik P (1991) Second-order characteristics for stochastic structures connected with Gibbs point processes. Math Nachr 151:95–100
Tello M-L, Tomalak M, Siwecki R, Gáper J, Motta E, Mateo-Sagasta E, (2005) Biotic urban growing conditions—threats, pests and diseases. In: Konijnendijk CC, Nilsson K, Randrup TB (eds) Urban forests and trees: a reference book. Springer, Berlin, pp 325–365
Tokola T (2006) Asia. In: Kangas A, Maltamo M (eds) Forest inventory. Methodology and applications. Springer, Dordrecht, pp 309–324
Tubby KV, Webber JF (2010) Pests and diseases threatening urban trees under a changing climate. Forestry 83:451–459
Turner MG, Gardner RH, O’Neill RV (2001) Landscape ecology in theory and practice. Springer-Verlag, New York
Tyrvainen L, Pauleit S, Seeland K, de Vries S (2005) Benefits and uses of urban forests and trees. In: Konijnendijk CC, Nilsson K, Randrup TB (eds) Urban forests and trees: a reference book. Springer, Berlin, pp 81–114
VanDerWal J, Falconi L, Januchowski S, Shoo L, Storlie C (2014) SDMTools. Species distribution modelling tools: tools for processing data associated with species distribution modelling exercises. R package version 1.1-13. http://CRAN.R-project.org/package=SDMTools. Accessed 15 May 2015
Westinga E, Mukashema A, van Gils H (2013) A comparison of fine resolution census and image-based national forest inventories: a case study of Rwanda. Forestry 86:453–461
Wiens JA (2001) The landscape context of dispersal. In: Clobert J, Danchin E, Dhondt AA et al (eds) Dispersal. Oxford University Press, Oxford, pp 96–122
Wiens JA (2009) Landscape ecology as a foundation for sustainable conservation. Landscape Ecol 24:1053–1065
Wu J (2007) Toward a landscape ecology of cities: beyond buildings, trees, and urban forests. In: Carreiro MM, Song YC, Wu J (eds) Ecology, planning, and management of urban forests: international perspectives. Springer, New York, pp 10–28
Zippeter WC (2008) Applying ecosystem management to urban forestry. In: Carreiro MM, Song Y-C, Wu J (eds) Ecology, planning, and management of urban forests: international perspectives. Springer-Verlag, New York, pp 97–108
Acknowledgments
We are grateful to Francis Goussard and Christelle Robinet (INRA Orléans) for valuable discussions regarding PPM inventory procedure. This research was partly funded by the Région Centre (project ADRIEN) and the INRA meta-program SMaCH—Sustainable Management of Crop Health (project SESAME). We are indebted to the editor and anonymous reviewers for their insightful comments and invaluable suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Rossi, JP., Garcia, J., Roques, A. et al. Trees outside forests in agricultural landscapes: spatial distribution and impact on habitat connectivity for forest organisms. Landscape Ecol 31, 243–254 (2016). https://doi.org/10.1007/s10980-015-0239-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-015-0239-8