
Abstract
Introduction
Clinical observations in patients with specific antibody deficiency treated for periods of time with IgG infusion have suggested that IgG may have a positive immunoregulatory effect on the production of specific antibodies against pneumococcal polysaccharides. We developed an in vitro model to test the effect of an IgIV preparation on the antibody production in response to a pneumococcal polysaccharide serotype and on the antibody and cytokine production in response to both a protein antigen and a pneumococcal polysaccharide antigen.
Methods
We studied 37 consecutive patients referred to our clinics for evaluation of their recurrent respiratory infections. Subjects were divided into two groups: 22 patients without SAD and 15 patients with SAD. PBMCs were left unstimulated or were stimulated with tetanus toxoid or pneumococcal polysaccharide serotype 19, in the presence of human albumin or IgIV. IgG anti-Pn-19 antibody, IL-4 and IFN-γ concentrations in culture supernatants were determined by ELISA.
Results
An increase in IgG anti-Pn-19 antibodies, associated with an increase in IFN-γ and a decrease in IL-4 production was observed in cultures stimulated with pneumococcal polysaccharide in the presence of IgIV when patients were analyzed together. The enhancing effect of IgIV was more significant for both IgG anti-Pn19 antibodies and IFN-γ in patients without SAD. In contrast, IgIV caused a significant decrease in IL-4 secretion in patients with SAD, which was associated with an increase in IgG anti-Pn19 antibodies in 3 of 7 of these patients.
Conclusions
These results suggest that IgIV has some immunomodulatory effect on the in vitro production of IgG anti-Pn19 antibodies and cytokine production in cell cultures stimulated with Pn-19 antigen and that this modulation may be associated with a Th1/Th2 regulatory mechanism. Further studies at a cellular and molecular level are in progress to examine if the differences in the in vitro modulatory response to IgIV in these two groups of patients may point to a functional defect in patients with SAD.
Similar content being viewed by others


Introduction
Therapeutic intravenous immunoglobulin (IgIV) consists of normal polyclonal IgG obtained from the plasma of several thousand healthy donors. It has been used over the past 25 years as a substitutive treatment for primary and secondary immunodeficiences, including severe cases of specific antibody deficiency (SAD) with normal serum immunoglobulin levels [1]. We have observed that in SAD patients treated with IgIV for 12 months to 2 years, re-evaluation of their immune response 6 months after discontinuing IgG replacement often revealed that specific anti-pneumococcal antibody production in vivo was improved, both spontaneously and after revaccination with the 23-valent pneumococcal polysaccharide vaccine. This was observed in young patients in whom it could be due to the effect of an ongoing maturation of the immune system, but also in older patients in whom a delayed maturation was less likely (R.U.S., unpublished observation). Based on these observations we decided to study the modulatory effect of IgIV on the in vitro production of antibodies and Th1/Th2 cytokines in patients with SAD and in patients with recurrent infections without SAD (No SAD).
IgIV has also been shown to have an immunomodulatory effect in autoimmune and inflammatory diseases [2–4]. In some situations, the presence of specific antibodies to an antigen has been shown to inhibit the production of antibodies. This is the case in the inhibition of antibody responses to Rh blood group antigens by anti-Rh hyperimmune gammaglobulin (Rhogam) and in the suppression of antibody responses to live viral vaccines (MMR) when maternal antibodies are still present in children at the time of immunization [5, 6]. However, several observations suggest that the presence of specific IgG antibodies may also have a positive immunoregulatory effect on the production of antibodies [7, 8]. Passive maternal antibodies do not alter responses to protein vaccines (DPT) that do not include live, attenuated viruses [6]. Transplacentally transmitted maternal antibodies against specific pneumococcal polysaccharides have not shown an inhibitory effect on antibody production against the same serotype in 6 and 12 months-old infants [9]. In contrast, specific anti-hepatitis B antibodies have been shown to promote the response to hepatitis B antigens in patients unresponsive to hepatitis B vaccine [7, 8, 10]. The immunomodulatory effect of IgIV in patients is likely to be a result of a variety of complex mechanisms that act in synergy.
Here we report the results of an investigation to determine if IgIV added to cell cultures from patients with recurrent infections has a modulatory effect on the immune response to protein or polysaccharide antigens in vitro. We were particularly interested in determining if there could be a differential effect on patients that had a normal specific antibody response to pneumococcal polysaccharides in vivo and those diagnosed to have SAD, based on poor responses to pneumococcal immunization.
Material and Methods
Patient Population
We studied 37 consecutive pediatric patients, two years of age or older of both genders, referred to our Children’s Hospital Allergy/ Immunology clinics for recurrent infections. Subjects were recruited regardless of race or ethnic background. Patients were evaluated in the clinic according to a standard protocol, which includes a detailed history of immunizations, infections and antibiotic use, laboratory evaluation, including immunoglobulin and IgG subclass determinations and the measurement of IgG antibodies specific to 14 pneumococcal polysaccharide serotypes by the WHO-recommended enzyme-linked immunosorbent assay (ELISA) [11]. As part of their evaluation, patients in this study received one dose of a 23-valent pneumococcal vaccine (Pneumovax 23; Merck, Whitehouse Station, New Jersey). Anti-pneumococcal polysaccharide antibodies were measured before and 4 to 6 weeks after immunization. Patients were enrolled into the study at the time of the post-immunization samples, before knowing the outcome of their responses to immunization. Thus, in patients diagnosed with SAD who required IgIV replacement therapy, all studies were performed on blood samples taken prior to the start of treatment.
For analysis, subjects were divided into two groups, 15 patients with diagnosis of SAD and 22 patients without SAD (No SAD). Patients with SAD were defined as having normal IgG, IgM, IgA and IgG subclass levels but poor IgG antibody responses to pneumococcal vaccines. Patients with immunoglobulin deficiencies, acute infections or debilitating conditions were excluded from the study. One 10 cc peripheral blood sample, collected in a sodium heparin tube was obtained 4–6 weeks after pneumococcal immunization, at the time of laboratory re-evaluation of the patient’s immune status. Consent from the subject’s legally authorized representative was obtained by the attending physician. Assent was obtained from children 7 years old or older.
Adequate IgG Antibody Response
For the purposes of establishing the diagnosis of SAD, an adequate IgG antibody response to an individual serotype was arbitrarily defined as a post-immunization antibody concentration of 1.3 μg/ml or greater. A normal response was defined according to the percentage of serotypes in the vaccine to which the patient responded. Normal children 2 to 5 years of age with a normal response were expected to have an adequate response to more than 50 % of the serotypes tested. Patients 6 years or older were normally expected to respond to more than 70 % of the serotypes tested [1].
Cell Preparation
Peripheral blood mononuclear cells (PBMC) were isolated from blood samples by Ficoll-Hypaque (Histopaque-1077, Sigma, St. Louis, MO) gradient centrifugation. The plasma fractions of the PBMC isolation procedures were collected and stored at −20 °C for future testing. PBMCs were washed in RPMI-1640 (Sigma), counted and resuspended in RPMI-1640 containing L-glutamine and 25 mM Hepes buffer and supplemented with 10,000 U/ml penicillin/10,000 μg/ml streptomycin (Sigma) and 10 % heat-inactivated fecal calf serum (FCS; Invitrogen, Carlsbad, CA).
PBMC Cultures for in vitro Antibody Production
The in vitro assay for antigen induced antibody secretion was performed according to a modified protocol developed by Chang & Sack (2001). This assay was originally developed to measure vaccine induced antibody production without any in vitro antigen stimulation [12]. We modified the assay by stimulating the PBMC with an optimal concentration of Pneumococcal polysaccharide serotype 19 F (Pn19F) antigen to evaluate the effect of IgIV on antigen-stimulated antibody production. Pn19F was selected because this serotype has been reported to be the best choice for in vitro stimulation [13].
PBMC were cultured at a concentration of 4 × 106 cells/ml in 12 × 75 mm culture tubes at 37 °C in 5 % CO2 in complete RPMI-1640 containing10% heat-inactivated FCS (Invitrogen) without (unstimulated) or with 20 μg/ml Pn19F (American Type Cell Culture Collection, Manassas, VA) in the presence of 5 mg/ml IVIG (Gamunex; Talecris Biotherapeutics, Research Triangle Park, NC) or of an equal amount of human albumin (HA; 25 % Solution, ZLB Behring AG, Switzerland). Unstimulated cells cultured in the presence of IgIV or HA were used as controls. After one day, cell cultures were washed twice with RPMI, re-suspended in complete RPMI medium without added IgIV or HA and were cultured for 5 additional days. At the end of the incubation period, the culture supernatants were removed and stored at −20 ° C. In vitro-produced IgG anti Pn19F antibodies were measured in culture supernatants by ELISA.
Measurement of in vitro Anti-Pneumococcal Polysaccharide Antibody Concentration by ELISA
IgG anti-Pn19 concentration was measured in culture supernatants using a modified ELISA protocol calibrated against the FDA 89-SF reference sample as previously described [11]. Microtiter 96-well plates (Nunc Maxisorp) were coated with 100 μl of 10 μg/ml pneumococcal serotype 19 F polysaccharide (Pn19; ATCC, Rockville, MD) in 0.01 M phosphate-buffered saline (PBS) at 37 °C for 5 hours. A standard serum sample with known concentration of IgG anti-Pn19F antibodies, was diluted 1:10 in PBS-Tween 20 containing 1.0 % bovine serum albumin (BSA) and pre-absorbed for 30 min at room temperature with S. pneumoniae C-polysaccharide (CPS; 500 μg/ml; Statens Seruminstitut, Denmark). This standard was serially diluted and run with each sample batch to obtain a calibration curve with measurement ranges of 1.3–10.0 ng/ml for IgG anti-Pn19F antibodies. The specificity and sensitivity of this method was tested in preliminary experiments, with serum samples with known anti-Pn19F IgG antibodies (previously measured using the WHO ELISA assay). The samples were serially diluted to reach concentrations within this measurement range for antibodies present in culture supernatants. The specificity for anti- Pn19F polysaccharide was not lost using this procedure. A control sample with known anti-IgG anti-Pn19 antibodies was also included in each run. Culture supernatants were added straight or diluted 1:2 to their respective wells in duplicate and the plates were incubated for 2 h at room temperature. The wells were then washed 3 times with PBS-0.05 % Tween 20 and a titrated amount of horse radish peroxidase labeled mouse anti-human IgGFc antibodies (Hybridoma Reagent Laboratories, Baltimore, MD) was added and the plates were incubated in the dark for 2 h at room temperature. Following another washing step, the enzyme bound was detected by the addition of tetramethyl-benzidine-dihydrochloride (TMB; Sigma Chemical Co., St. Louis, MO) in citrate phosphate buffer. The IgG anti-Pn19F antibody concentration (in nanograms per milliliter) was calculated by measuring the absorbance (optical density at 450 nm) against the standard curve. IgG anti-Pn19F antibody levels in this standard was previously determined using the FDA 89-SF reference sample (CBER, U.S. Food and Drug Administration, Rockville, MD). Complete medium incubated without cells for the same time period and tested for anti-Pn19F antibody levels was used as negative control. No antibody concentrations were detected.
PBMC Cultures for in vitro Cytokine Production
PBMCs were cultured at a concentration of 5 × 105 cells/ml in 12 × 75 mm culture tubes at 37 °C in 5 % CO2 in RPMI-1640 containing10% heat-inactivated FCS (Invitrogen) with 20 μg/ml Pn19F or with tetanus toxoid (TT) adsorbed USP (1:500; Sanofi Pasteur, Swiftwater, PA) in the presence of 5 mg/ml IgIV or of an equal amount of HA. Unstimulated cells cultured in the presence of IgIV or HA were used as controls for specificity of response. Cultures were incubated for 48 h and supernatants were removed, transferred into cryovials and stored at −20 ° C. In vitro-produced interferon gamma (IFN-γ) and interleukin 4 (IL-4) were measured in culture supernatants using commercial ELISA kits (BD Biosciences, San Diego, CA and Invitrogen, Carlsbad, CA). The minimum detectable concentrations were 1.0 pg/ml for IFN-γ and 2.0 pg/ml for IL-4. Complete medium incubated without cells for the same time period and tested for concentrations of IFN-γ and IL-4 was used as negative control. No cytokine concentrations were detected.
Statistical Analysis
Statistical analysis was performed using the GraphPad Prism software. Differences between cultures and patient groups were analyzed for statistical significance using the Wilcoxon matched pairs test (one-tailed) and the threshold of significance was set at a p value of <0.05.
Approval for the study was obtained from the Louisiana State University Health Science Center and from the Children’s Hospital Institutional Review Board Offices.
Results
Effect of IgIV on the in Vitro IgG Anti-Pn19F Antibody Production
Analysis of the IgG anti-Pn19F antibody concentrations revealed that there were no significant differences between unstimulated and antigen-stimulated cultures from any of the patient groups (Fig. 1). However, an enhancing effect on IgG anti-Pn19F antibody concentrations was observed in cultures with IgIV in comparison to control cultures with HA, when results from all patients where compared (Fig. 1a, p < 0.05). Interestingly, when patients were divided into those without SAD and those with SAD, the enhancing effect was statistically significant (p < 0.05) for patients without SAD but not for patients with SAD (Fig. 1b and c). However, when each patient was analyzed individually, we found the enhancing effect of IgIV in Pn19F antigen- stimulated cultures from 8 of 15 (53.3 %) patients with SAD (Not shown).

Effect of IgIV on the production of IgG anti-Pn19 antibodies by PBMC from patients with recurrent upper respiratory infections (a), patients without SAD (No SAD) (b) and patients with SAD (c). Cell cultures were left unstimulated (Uns) or were stimulated with pneumococcal polysaccharide serotype 19 F (Pn-19), in the presence of human albumin (HA) or IgIV for 24 h, as described in Materials and Methods. Results are mean ng/ml ± SEM of IgG anti-Pn19 concentrations. p < 0.05 values between cultures with IgIV vs HA are considered significant
Effect of IgIV on the in vitro Production of INF-γ and IL-4
Antibody production is regulated in part by cytokine production. We selected IFN-γ and IL-4 production to test if IgIV had any effect on the Th1/Th2 immune regulation in PBMC cultures stimulated for 48 h with TT or Pn19F antigens. Of interest is the observation that both, IFN-γ and IL-4 concentrations in cultures from patients with SAD are significantly lower than the concentrations in cultures from patients without SAD (Figs. 2 and 3).

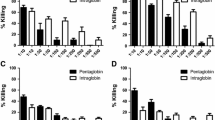
Effect of IgIV on the production of IFN-γ by PBMC from patients with recurrent upper respiratory infections (a), patients without SAD (No SAD) (b) and patients with SAD (c). Cell cultures were left unstimulated (Uns) or were stimulated with tetanus toxoid (TT) or pneumococcal polysaccharide serotype 19 (Pn-19), in the presence of human albumin (HA) or IgIV for 48 h, as described in Materials and Methods. Results are mean pg/ml ± SEM of IFN-γ concentrations. p < 0.05 values between cultures with IgIV vs HA are considered significant

Effect of IgIV on the production of IL-4 by PBMC from patients with recurrent upper respiratory infections (a), patients without SAD (No SAD) (b) and patients with SAD (c). Cell cultures were left unstimulated (Uns) or were stimulated with tetanus toxoid (TT) or pneumococcal polysaccharide serotype 19 (Pn-19), in the presence of human albumin (HA) or IgIV for 48 h, as described in Materials and Methods. Results are mean pg/ml ± SEM of IL-4 concentrations. p < 0.05 values between cultures with IgIV vs HA are considered significant
Unstimulated cultures in the presence of HA or IgIV alone, and cultures stimulated with either TT or Pn19F antigens in the presence of HA were used as controls to analyze the effect of IgIV in cultures. Significant increase in IFN-γ production over concentrations in unstimulated cultures in the presence of either IgIV or HA alone was seen in all of the antigen- stimulated cultures when all patients were analyzed together (Fig. 2a, p < 0.05). The addition of IgIV to antigen-stimulated cultures induced a significant increase (p < 0.05) in IFN-γ production in Pn19F but not in TT antigen-stimulated cultures (Fig. 2a and b). The enhancing effect of IgIV was significant in Pn19F antigen-stimulated cultures from patients with No SAD (Fig. 2b, p < 0.05) but not in SAD patients (Fig. 2c). However, individual analysis demonstrated that 4 of 10 patients with SAD showing enhanced in vitro anti-Pn19 antibody production also showed increased IFN-γ concentrations in the same cultures with IgIV (results not shown).
The effect of IgIV on in vitro IL-4 production in cultures from all patients is shown in Fig. 3a. Baseline concentrations of IL-4 were similar in supernatants from unstimulated cultures in the presence of either IgIV or HA. IL-4 production was significantly higher in TT antigen-stimulated cultures in comparison to cultures stimulated with Pn19F antigen in cultures from all patients (Fig. 3a, p < 0.05). An inhibitory effect of IgIV on IL-4 production was significant in cultures stimulated with Pn19F in patients with SAD but not in patients without SAD (Fig. 3b and c, p < 0.05). Individual analysis of results from SAD patients showed that 3 of 7 patients with increased in vitro anti-Pn19 antibody had decreased IL-4 concentrations in cultures with IgIV (results now shown).
Discussion
There has been a concern that the use of IgIV and therefore of passive IgG antibodies might interfere with the development of the patient’s own immune response [14, 15]. Our results show that IgIV does not inhibit an antibody response in vitro. On the contrary, we demonstrate that IgIV had an enhancing effect on the spontaneous production of IgG antibodies in cell cultures from patients with recurrent respiratory infections.
We ruled out that this observation was due to adherence and later shedding of anti Pn19F antibodies in the IgIV added to cell cultures. The increase in specific IgG antibodies was not observed in unstimulated cultures. However, there was background production of antibodies in all cell cultures regardless of the addition of IgIV. This background IgG antibody concentration may have been due to spontaneous in vitro antibody production by the patient’s cells in culture. The presence of these antibodies was dependent on the presence of cells in cultures, as no antibodies were detected in the negative controls. A recent study demonstrated a brief appearance of a subset of B cells in the peripheral blood of healthy individuals after receiving a booster dose of a heptavalent pneumococcal –CRM197 conjugate vaccine, that spontaneously secreted IgG [16]. A similar B cell subpopulation, stimulated by immunization with one dose of the Pneumovax 23 vaccine, may have been responsible for the spontaneous IgG antibody production in vitro. This possibility would have to be further investigated in our patients. Our experimental results measuring anti-polysaccharide antibodies do not allow to differentiate if IgIV has an effect specific for the polysaccharide response or if it is a non-specific effect increasing antibody production in general. These two possibilities would need to be resolved in further experimental studies.
We also demonstrate here that antigen-stimulated PBMC from both patients with SAD and patients without SAD have the ability to produce IFN-γ and IL-4 in vitro. Both IFN-γ and IL-4 were found to be lower in patients with SAD in comparison to patients without SAD, which might be one of the immune regulatory mechanisms affected in SAD.
The enhancement of in vitro antibody production reached significance in patients without SAD, but not in patients with SAD, although a similar trend could be observed in this group. The group size (15 SAD vs 22 No SAD patients) might be the reason for not reaching significance. However, when the results were analyzed individually we observed that this enhancing effect was also seen in cultures from 8 of 15 patients with SAD. This observation corresponds to the clinical observation that some patients do and others do not improve anti-pneumococcal antibody production in vivo after IgIV treatment. Those that do not improve their antibody production in vivo frequently need to be put back on IgG replacement treatment if their infections also continue. Whether or not IgIV has a similar mechanism of action in vivo and in vitro cannot be evaluated based on these results. However, the effect of IgIV in vitro was observed at a concentration of 5 mg/ml, which corresponds to the serum level achieved therapeutically with intravenous infusion of 0.5 g/kg body weight [17].
The possible mechanisms of action of IgIV reflect the importance of natural antibodies, generated independently of exposure to foreign antigens and present in normal serum, in the maintenance of immune homeostasis in healthy people, which may have implications in the immune response to vaccines [4, 18, 19]. Natural antibodies are believed to be essential for the immunoregulatory effects of immune globulin. These “natural” antibodies are mainly autoantibodies, capable or interacting with idiotypes of other antibodies in the IgIV preparation to form dimmers with complementary idiotypes. The formation of idiotype-idiotype dimeres may account for some of the modulatory mechanisms and clinical effects of IgIV [2, 20].
The mechanisms for the enhancing effect of IgIV on antibody production in our study may have been the result from a direct effect of IgIV on B cells or through a more complex modulatory mechanism involving other cells [21]. A direct interaction between IgIV and a series of proteins expressed in human cells was recently demonstrated using immunoprecipitation and LC-MS analysis [22]. This interaction was found to be involved in antigen processing and presentation in human B cells [23]. It has also been shown that a direct IgIV modulation of human B cells could be linked to increased intracellular transduction pathways induced by the phosphorylation of extracellular signal-regulated kinases in human B cells [24]. These and other possibilities still need to be investigated as possible regulatory mechanisms.
The mode of action of IgIV is complex, involving modulation of expression and function of Fc receptors, interference with activation of complement and the cytokine network, modulation of idiotype network, and activation, differentiation and effector functions of T and B cells and of antigen presenting cells [3, 19]. From our study and many other studies, it is clear that IgIV has immune modulatory properties that are still being investigated. Our observation of spontaneous cytokine production in unstimulated cultures in our study has also been reported earlier [25]. The up-regulation of IFN-γ and down regulation of IL-4 by the presence of IgIV in antigen-stimulated cultures suggests that a Th1/Th2 regulatory mechanism could be involved in the production of antibodies against polysaccharides. Our finding of an effect both on IFN-γ and IL-4 production in a subgroup of patients with SAD further supports the modulatory role of IgIV in vitro and its association with the clinical observation seen in some of these patients after IgIV treatment.
In conclusion, our results show that IgIV modulates, to some extent, the in vitro production of IgG anti-Pn19 antibodies and that the observation of cytokine production changes in vitro suggests that this modulation is associated with a Th1/Th2 regulatory mechanism. Our observations of antibody and cytokine production in patients with SAD suggest that this is a heterogeneous group of patients caused by different regulatory mechanisms that could also explain important differences in the long-term response to IgG replacement therapy. Further studies at a cellular and molecular level are in progress to examine if these differences may point to a functional defect in patients with SAD.
References
Paris K, Sorensen RU. Assessment and clinical interpretation of polysaccharide antibody responses. Ann Allergy Asthma Immunol. 2007;99(5):462–4.
Kazatchkine MD, Kaveri SV. Immunomodulation of autoimmune and inflammatory diseases with intravenous immune globulin. N Engl J Med. 2001;345(10):747–55.
Bayry J, Dasgupta S, Misra N, Ephrem A, Van Huyen JP, Delignat S, et al. Intravenous immunoglobulin in autoimmune disorders: an insight into the immunoregulatory mechanisms. Int Immunopharmacol. 2006;6(4):528–34.
Siberil S, Elluru S, Graff-Dubois S, Negi VS, Delignat S, Mouthon L, et al. Intravenous immunoglobulins in autoimmune and inflammatory diseases: a mechanistic perspective. Ann N Y Acad Sci. 2007;1110:497–506.
Liumbruno GM, D’Alessandro A, Rea F, Piccinini V, Catalano L, Calizzani G, et al. The role of antenatal immunoprophylaxis in the prevention of maternal-foetal anti-Rh(D) alloimmunisation. Blood Transf. 2010;8(1):8–16.
Glezen WP. Effect of maternal antibodies on the infant immune response. Vaccine. 2003;21(24):3389–92.
Celis E, Chang TW, Stricker L, Tiebout RF, Zeijlemaker WP. Role for monoclonal hepatitis B antibody in hepatitis B immunisation programmes. Lancet. 1985;1(8439):1219.
Celis E, Kato I, Miller RW, Chang TW. Regulation of the human immune response to HBsAg: effects of antibodies and antigen conformation in the stimulation of helper T cells by HBsAg. Hepatology. 1985;5(5):744–51.
Inostroza J, Villanueva S, Mason K, Leiva LE, Sorensen RU. Effects of absorption with pneumococcal type 22 F polysaccharide on maternal, cord blood, and infant immunoglobulin G antipneumococcal polysaccharide antibodies. Clin Diagn Lab Immunol. 2005;12(6):722–6.
Celis E, Abraham KG, Miller RW. Modulation of the immunological response to hepatitis B virus by antibodies. Hepatology. 1987;7(3):563–8.
Sorensen R, Leiva L, Javier F, Sacerdote D, Bradford N, Butler B, et al. Influence of age on the response to Streptococcus pneumoniae vaccine in patients with recurrent infections and normal immunoglobulin concentrations. J Allergy Clin Immunol. 1998;102(2):215–21.
Chang HS, Sack DA. Development of a novel in vitro assay (ALS assay) for evaluation of vaccine-induced antibody secretion from circulating mucosal lymphocytes. Clin Diagn Lab Immunol. 2001;8(3):482–8.
Rijkers GT, Mosier DE. Pneumococcal polysaccharides induce antibody formation by human B lymphocytes in vitro. J Immunol. 1985;135(1):1–4.
Kumpel BM. On the mechanism of tolerance to the Rh D antigen mediated by passive anti-D (Rh D prophylaxis). Immunol Lett. 2002;82(1–2):67–73.
Brinc D, Le-Tien H, Crow AR, Siragam V, Freedman J, Lazarus AH. Transfusion of IgG-opsonized foreign red blood cells mediates reduction of antigen-specific B cell priming in a murine model. J Immunol. 2008;181(2):948–53.
Clutterbuck EA, Salt P, Oh S, Marchant A, Beverley P, Pollard AJ. The kinetics and phenotype of the human B-cell response following immunization with a heptavalent pneumococcal-CRM conjugate vaccine. Immunology. 2006;119(3):328–37.
Rand KH, Gibbs K, Derendorf H, Graham-Pole J. Pharmacokinetics of intravenous immunoglobulin (Gammagard) in bone marrow transplant patients. J Clin Pharmacol. 1991;31(12):1151–4.
Elluru SR, Vani J, Delignat S, Bloch MF, Lacroix-Desmazes S, Kazatchkine MD, et al. Modulation of human dendritic cell maturation and function by natural IgG antibodies. Autoimmun Rev. 2008;7(6):487–90.
Vani J, Elluru S, Negi VS, Lacroix-Desmazes S, Kazatchkine MD, Bayary J, et al. Role of natural antibodies in immune homeostasis: IVIg perspective. Autoimmun Rev. 2008;7(6):440–4.
Rossi F, Sultan Y, Kazatchkine MD. Anti-idiotypes against autoantibodies and alloantibodies to VIII:C (anti-haemophilic factor) are present in therapeutic polyspecific normal immunoglobulins. Clin Exp Immunol. 1988;74(2):311–6.
Schwab I, Nimmerjahn F. Intravenous immunoglobulin therapy: how does IgG modulate the immune system? www.naturecom/reviews/immunol. 2013;13(March):176–88.
Proulx DP, Aubin E, Lemieux R, Bazin R. Spontaneous internalization of IVIg in activated B cells. Immunol Lett. 2009;124(1):18–26.
Proulx P, Aubin E, Lemieux R, Bazin R. Inhibition of B cell-mediated antigen presentation by intravenous immunoglobulins (IVIg). Clin Immunol. 2010;135(3):422–9.
Dussault N, Ducas E, Racine C, Jacques A, Pare I, Cote S, et al. Immunomodulation of human B cells following treatment with intravenous immunoglobulins involves increased phosphorylation of extracellular signal-regulated kinases 1 and 2. Int Immunol. 2008;20(11):1369–79.
Ekerfelt C, Ernerudh J, Jenmalm MC. Detection of spontaneous and antigen-induced human interleukin-4 responses in vitro: comparison of ELISPOT, a novel ELISA and real-time RT-PCR. J Immunol Methods. 2002;260(1–2):55–67.
Acknowledgments
This study was supported by a grant from Talecris Biotherapeutics, Inc., Research Triangle Park, NC.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Leiva, L.E., Monjure, H. & Sorensen, R.U. Modulatory Role of Intravenous Gammaglobulin (IgIV) on the in vitro Antibody Response to a Pneumococcal Polysaccharide Antigen. J Clin Immunol 35, 206–212 (2015). https://doi.org/10.1007/s10875-014-0120-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10875-014-0120-6