
Abstract
Recently, there has been considerable interest in the idea that mutational robustness enhances the propensity for future adaptations, i.e. evolvability, if evolution proceeds over a neutral network that extends far throughout a fitness landscape. While the genetic neutral network (NN-G) model may have important implications to our understanding of evolution, little has been done to integrate these theoretical developments with empirical evidence that heritable phenotypes can also originate and become fixated as a result of changes in the environment. In this brief commentary, I reconsider the role of environmental change in the adaptation of species and ask whether positive robustness-evolvability relationships might exist not only for genetic but also environmental buffering. In particular, I ask whether the insensitivity of species fitness towards variability in its environment can have a positive influence on the likelihood of future environment-induced adaptations (i.e. ecological opportunities) in a manner analogous to that proposed by the NN-G model. After outlining scenarios where such a counter-intuitive relationship appears plausible, I comment on the merits of evolutionary theories that can integrate complementary pathways to adaptation under static and time-variant environments. I also speculate on some of the features that such a theory might have.
Similar content being viewed by others


Text box: glossary
Cryptic genetic variation (CGV)
CGV is a population property that involves the following features: (1) in its native environment a population maintains high levels of genetic diversity but exhibits relatively few trait differences, i.e. genetic differences are cryptic and (2) when exposed to new environmental conditions (or systematically exposed to new alleles), the population displays new heritable phenotypic variability.
Neutral network
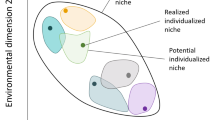
A neutral network is defined as a connected graph of nodes with equal fitness. One can consider it a connected set of external and internal conditions within which a species has the same fitness. Notice that connectedness implies that each node within the network can be reached by every other without changing the species fitness along the path of arcs. Illustrations of genetic and environmental neutral networks are given in Fig. 1.

a Nodes represent genotypes and connections between nodes indicate that two genotypes differ by only a single mutation. The connected graph of black nodes illustrates a genetic neutral network (NN-G). The neutral network implies that for a particular environment, none of these genotypes are selectively distinguishable. Square grey nodes represent genotypes that are mutationally accessible from the NN-G but that have phenotypic and selective differences when compared to members of NN-G. As observed in gene regulatory network simulations (Ciliberti et al. 2007) and abstract genome:proteome models (Whitacre and Bender 2010), it is assumed that the phenotypes associated with grey nodes will change depending on their position in genotype space. Thus, as NN-G grows and extends throughout larger regions of genotype space, it is assumed that the number of unique phenotypes accessible from NN-G will also grow (illustrated in bottom figure). b Nodes represent environmental patches and connections between nodes indicate that two patches are physically connected so that a population can move directly from one patch to the other. The connected graph of black nodes illustrates an environmental neutral network (NN-E). The neutral network implies that a population can move to different patches without consequences to survival or reproductive success. While this does not preclude the possibility that different patches in NN-E are associated with mildly distinct trait distributions, it is assumed that these distinctions do not result in changes to selection that have evolutionary consequences. As with the NN-G pathway, positive associations may exist between the number of accessible environments in which species fitness is robust (NN-E) and the number of accessible environments that induce heritable (and sometimes selectively relevant) phenotypic differences in a population (illustrated in bottom figure)
Genetic Neutral Network (NN-G)
NN-G is a neutral network in which the class of condition changes is restricted to single gene mutations. Such networks represent fitness-neutral regions within a classic fitness landscape.
Environmental neutral network (NN-E)
NN-E is a neutral network in which the class of condition changes is restricted to changes in the environment experienced by an organism during development.
Evolvability
Evolvability refers to the propensity of a species to discover heritable and beneficial phenotypes. Evolvability requires access to distinct heritable traits and it requires that some heritable differences can be transformed into beneficial innovations during development within a particular environment. The first requirement—the ability to access heritable phenotypic variation—is an important precondition and often used proxy for evolvability.
Robustness
Robustness describes the insensitivity of some functionality or measured biological trait to a set of distinct conditions. This article focuses primarily on the robustness of high level traits that influence survival and fecundity.
Mutational robustness
This refers to the extent that species fitness is robust towards genetic mutations.
Introduction
The genetic neutral network hypothesis for adaptation
There are a growing number of studies reporting evidence of a positive relationship between mutational robustness and evolvability at microevolutionary scales (Aldana et al. 2007; Bloom et al. 2006; Babajide et al. 1997; van Nimwegen and Crutchfield 2000; Ciliberti et al. 2007; Wagner 2008; Whitacre and Bender 2010) with possible repercussions to our understanding of macroevolution and speciation (Gavrilets 1997) (for reviews see (Wagner 2008; Wagner 2008). The hypothesis put forth is that networks of fitness neutral genotypes result in mutational robustness and reduced accessibility of heritable phenotypes over short timescales. With little genetic variation expressed as phenotypic variation, natural selection has few immediate options for modifying traits. On the other hand, genetic drift over neutral/buffered mutations can also provide mutational access to many distinct heritable phenotypes that are reached from directly off the genetic neutral network over longer periods of time. This leads to the seemingly paradoxical conclusion (resolved through a separation of timescales) that the suppression of heritable phenotypic variation can ultimately increase the accessibility of distinct heritable phenotypes (Wagner 2008).
Because access to distinct heritable phenotypes is a prerequisite for adaptation, mutational robustness has thus been described as a potential facilitator of evolvability, i.e. the propensity for a species to adapt. This genetic neutral network (NN-G) hypothesis has been supported by recent computer models of biological systems (Aldana et al. 2007; Bloom et al. 2006; Babajide et al. 1997; van Nimwegen and Crutchfield 2000; Ciliberti et al. 2007; Wagner 2008; Whitacre and Bender 2010) and appears consistent with the available data related to biomolecular evolution (Bloom et al. 2006; Wagner 2008). The model clearly relies on a gene-centric view of evolution where novel alleles are encountered and occasionally result in novel and selectively relevant phenotypes.
Environment-induced adaptation
While the genetic basis of heredity is not disputed, the introduction of novel alleles is not the only way that heritable phenotypes originate (Palmer 2004; Gibson and Dworkin 2004; Schlichting 2008; Waddington 1953; Waddington 1957; Schmalhausen and Dobzhansky 1949; West-Eberhard 2005; West-Eberhard 2003; Barrett and Schluter 2008). Because the phenotype is the result of self-environment organization, those conditions provided by an organism’s genetic background, environmental background, and their interaction during development, will determine the qualitative character, quantitative attributes, and the timing of expressed traits. Changes in environmental inheritance due to movement (e.g. migration, seed dispersal), external perturbation [e.g. geologic cycles, ecological regime shifts, epigenetic inheritance (Anway et al. 2005)], and various forms of local environment shaping [e.g. niche construction (Odling-Smee et al. 1996; Day et al. 2003), sexual selection (West-Eberhard 2003), behavioural and genetic coevolution (Agrawal 2001), parental inheritance (Uller 2008; Jablonka et al. 1995), cultural inheritance (Dawkins 1983; Dennett 1995)] can expose the conditional plasticity of a trait, i.e. phenotypic plasticity. The role that the environment plays in the origination of new traits and its consequences to adaption have been discussed at length in (West-Eberhard 2003; Pfennig et al. 2010; Sultan 2007) [but also see (Agrawal 2001; Schlichting and Pigliucci 1998; Newman 1994)].
At the population level, the conditional exposure of trait variation is often discussed as a phenomena known as cryptic genetic variation (CGV) or “hidden reaction norms” or “genetic charge” (Le Rouzic and Carlborg 2008) [for reviews see (Gibson and Dworkin 2004; Schlichting 2008; McGuigan and Sgrò 2009)]. CGV describes heritable phenotypic variation that is hidden under “normal conditions” but that is released in the presence of novel conditions. Studies of CGV have found evidence that novel conditions can come both in the form of novel alleles and novel environments, and that the phenotypic consequences can be remarkably similar between these perturbation classes.
For circumstances where the environment induces a new adaptive trait, the inheritance of this trait is generally expected to depend on subsequent genetic modifications. While the persistence of a change to the environment, exposure to similarly modified environments, and trans-generational carry-over effects after a short-lived environmental change (Jablonka et al. 1995) can, in principle, induce and maintain novel traits, it is genetic assimilation (Waddington 1953; Waddington 1957; Schmalhausen and Dobzhansky 1949) and genetic accommodation (West-Eberhard 2005; West-Eberhard 2003) that act to preserve the inheritance of phenotypic adaptations in the face of further environmental modifications (West-Eberhard 2003; Pigliucci et al. 2006).
At present, it is not clear whether the origination of heritable phenotypes by genetic or environmental novelty has been more relevant to evolution (McGuigan and Sgrò 2009) however, the clearest available evidence suggests that these pathways are equally common (Palmer 2004).West-Eberhard (2003) and Schlichting and Pigliucci (1998) argue that environment-induced phenotypic variation is a more likely source of heritable change, at least partly due to evidence that environment-induced trait variation is more common (Houle et al. 1996; Gibson 2008), and potentially orders of magnitude more common (Lynch 1988), in populations and species compared to gene-induced trait variation.
For an individual species, the pathway—genetic or environment-induced phenotypic novelty—to adaptation most often observed will likely depend on several factors such as properties related to the genetic architecture (e.g. sign epistasis, modularity), environment complexity, rates of genetic and environmental change, effective population size, etc. My aim in this commentary is not to argue for the dominance of one pathway over another. Instead I entertain a view of evolution where these pathways are conditionally relevant and at times complementary and within this setting I consider whether recent advances in understanding the NN-G pathway could provide new insights into the alternative environment-induced pathway.
One of the most intriguing insights from NN-G is the idea that mutational robustness—the observation of attenuated phenotypic effects from mutations—can actually increase access to distinct heritable phenotypes (Wagner 2008). Here I consider whether such counter-intuitive relationships between robustness and evolvability are a more general feature of biological evolution. More precisely, I consider whether the insensitivity (robustness) of fitness toward environmental variability might in some cases positively influence the likelihood that a species or deme is exposed to environments with heritable phenotypic consequences, as illustrated in Fig. 1.
Robustness and adaptation within complex environments
To understand this proposal, consider a spatially distributed, heterogeneous environment that can be represented by a connected network of environmental patches. Each patch represents an unique physical environment and movement from one patch to another occurs over a timescale of generations. When the fitness of a species or deme is robust across a connected set of environmental patches (i.e. each with similar stabilizing selection), then by definition, the deme is able to migrate over this neutral sub-network (NN-E) without consequences to reproductive success or survival. Assuming the environment is heterogeneous and structured, different regions of the NN-E will connect to somewhat unique environmental patches that are not members of NN-E and each of these may have distinct phenotypic consequences that are differentially expressed across a genetically diverse population, i.e. the conditional release of CGV. On occasion, these environments may reveal new ecological opportunities for some individuals and lead to changes in selection. In particular, if some of the plastic responses are adaptive this will increase the likelihood that alleles contributing to this adaptive response will become fixated in the population. These plastic traits will then be expressed each time individuals revisit similar environments and would likely be subject to modification (e.g. in timing, environmental cues) through genetic accommodation, e.g. (West-Eberhard 2005; Suzuki and Nijhout 2006). On the other hand, subsequent passive/buffered genetic evolution as well as selection for polygenic traits may alternatively lead to genetic assimilation (Waddington 1953; Waddington 1957; Schmalhausen and Dobzhansky 1949) whereby trait changes tend to remain even after the deme relocates to other patches. For either outcome, it is important to emphasize that genetic changes are not initiating the exposure of phenotypic novelty even though they play a role in creating it and making it heritable to future generations (West-Eberhard 2005).
So long as the size of NN-E is positively associated with the number of accessible patches that have unique phenotypic consequences, the relationship between robustness and evolvability would appear to be compatible with environment-induced adaptations. In other words, the insensitivity of fitness to environmental variability (increase in NN-E) would have a positive influence on the accessibility of environments that invoke distinct and heritable phenotypic consequences, thereby increasing the long-term likelihood for adaptive change.
The required correlation between NN-E size and the variety of patches accessible from NN-E depends on several conditions, some of which do not appear to be under the control of a species. For instance, one prerequisite for accessing ecological opportunities is the existence of a heterogeneous and structurally complex environment. This requirement may however be more easily satisfied than it would first appear. For instance, a recent review of empirical data and simulation studies has concluded that physical environments commonly display complex patterns that emerge from several types of scale-dependent feedback between organisms and their environment and moreover that this pattern formation does not require initial heterogeneity within the physical environment (Rietkerk and van de Koppel 2008).
Any relationship between NN-E and evolvability also depends upon certain topological properties in the NN-E which might be difficult to achieve in networks constrained to low dimensional spaces such as those implied in the environmental patch example (Gavrilets 1997; Reidys et al. 1997). However, this limitation probably arises more from the use of an abstract network model than from actual limitations of ecosystems embodied within a physical environment. In particular, properties such as multi-scaling (Rietkerk and van de Koppel 2008; Ruokolainen et al. 2009), time-variance (e.g. in material flux and organizational structure), and niche construction can result in a “nested hierarchy of patch mosaics” (Wu and Loucks 1995) with multiplicative effects on the number of distinct environmental conditions that can be experienced despite the fact that organisms are confined to a low-dimensional physical space.
While the NN-E pathway just described has a close correspondence to the NN-G model, NN-E phenomena might also arise at other timescales relative to the reproductive cycle and by means other than migration. For instance, multi-cellular eukaryotes are regularly exposed to different environmental conditions where they must execute different functions that contribute to their survival. Increased flexibility in addressing such varied conditions may in some cases directly contribute to the total number of unique accessible features within their local environment. For instance, increased habitat range (Feduccia 1999), foraging range, predator avoidance, flexibility in the materials used for nest building, and in the resources that can be consumed and metabolized (Jernvall et al. 1996) each constitute examples where the robustness of a high-level trait (e.g. fitness) has been increased, each afford a multitude of further environmental conditions that can be experienced, and each of these novel environmental conditions has the potential to induce previously cryptic trait differences in a deme. In exceptionally rare cases, morphological adaptations occur that ‘unknowingly’ facilitate robustness to an astronomical number of environmental conditions and these can lead to the exposure of many ecological opportunities that unfold over space and time, e.g. the adaptive radiations that proceeded after evolution of the fourth cusp in the mammalian tooth (Jernvall et al. 1996) and after evolution of flight in birds (Feduccia 1999).
More generally, I argue that it is robustness of high level traits to environmental variation that is important to the NN-E pathway and that different response classes for achieving this robustness could each potentially alter the propensity for future adaptations. Such response classes include environment shaping, environment tracking, adaptive phenotypic plasticity of lower-level traits, canalization, and various combinations of these. For example, shaping one’s environment can sometimes stabilize external environmental features that influence a wide range of biological processes, e.g. temperature regulation. The ability to create environmental conditions when needed instead of relying entirely on the environment has an obvious and direct influence on the range of viable habitats. West-Eberhard argues that one particular example of the NN-E pathway, environment simplification, is a common type of environment-shaping that provides exceptional ecological opportunities for specialization (West-Eberhard 2005; West-Eberhard 2003). While humans are the most remarkable users of environment shaping, the extent that this occurs in other species is also impressive (Laland and Sterelny 2006).
Discussion
From a theoretical perspective, the possibility of complementary pathways for exposing heritable phenotypic novelty is in some ways desirable as it permits/explains a broader range of conditions under which the adaptation of species can proceed, e.g. within predominantly static and simple as well as time-variant and complex environments. Conversely, if the findings in (Palmer 2004) reflect a more general trend whereby genetic and environment-induced adaptations have both substantially contributed to the evolution of species, then any theories of evolution that do not account for both pathways are either fragile to or reliant upon a dynamic environment. Here I have suggested a broad relationship may exist between robustness and evolvability, facilitating adaptation under different conditions. While speculative, this view of evolution has broad implications in theoretical biology that are worth briefly elaborating upon.
For instance, a recent study has found that mutational robustness has a strong positive relationship with the genomic complexity of a species (Sanjuan and Elena 2006). One important implication from that study is that the likelihood of adaptations through the NN-G pathway has been subject to change over the course of evolutionary history. More generally, and assuming adaptations predominantly arise through NN-G and NN-E pathways, it would follow that a species’ propensity to adapt could be indirectly modified by changes in mutational (or environmental) robustness, which itself may evolve over time either as a result of direct selection, indirect selection, or congruency (including passive/neutral evolution) with selection (de Visser et al. 2003). Based on current understanding of these alternatives, direct selection for mutational robustness appears likely to occur only for highly restrictive conditions while direct selection for robustness against environmental variations is believed to be a common feature of evolution in spatially and temporally-rich environments (de Visser et al. 2003; Meiklejohn and Hartl 2002; Newman 1994).
Based on these considerations, I speculate that a myopic selection for greater robustness towards variable environmental conditions (experienced during a lifetime) could also positively affect the level of exposure (over many generations) to future environmental conditions with selectable phenotypic consequences. This hints at a process by which natural evolution indirectly and unintentionally facilitates its own future evolvability; a requirement for open-ended evolution that is not easily arrived at based on the NN-G relationship between mutational robustness and adaptation.
As stated, this conjecture leaves questions remaining about the origins of the NN-G pathway. In one plausible explanation, Meiklejohn and Hartl argue that robustness towards mutational and environmental perturbation classes is typically congruent; an idea most easily appreciated at molecular and cellular levels due to the clear physical similarities that these perturbation classes have on gene expression (Meiklejohn and Hartl 2002) [also see (de Visser et al. 2003; Szollosi and Derenyi 2009; Ancel and Fontana 2000)]. If such congruency is a common feature of life, then mutational robustness and the NN-G pathway might have emerged as an important by-product of evolving systems selectively driven by survival and reproductive success within a variable environment. This implies that properties of the genotype:phenotype map (and similarly the smoothness/ruggedness and other properties of the fitness landscapes on which genetic evolution now proceeds) are neither a fortuitous accident nor a common ensemble property of biological systems as is sometimes suggested (Kauffman 1993) but instead are an indirect and unavoidable consequence of evolution in a complex environment.
While this proposed congruence in robustness appears to increase the likelihood of adaptations originating through the NN-G pathway, it could have a still stronger affect on the likelihood of NN-E associated adaptations. This would be the case if genetic neutrality is itself condition-dependent or in other words, if the neutrality of mutations generally depends on both the genetic and environmental background. Arguments supporting the presence of these cryptic GxE interactions are given in (Olivieri 2009). In this scenario, NN-GFootnote 1 helps to facilitate the accumulation of CGV in natural populations and in turn would elevate the diversity of phenotypic responses within each novel environment. In short, each of these pathways may positively reinforce the other.
What predictions might one expect based on this proposed model of evolution? In a recent study, an analysis of phylogenetic data has found that speciation rates are highly similar between closely related species yet vary greatly between distantly related species (Venditti et al. 2010). Interestingly, when integrating their findings with other evidence from the literature, the authors propose that these speciation events are best described by an environment-induced pathway that becomes heritable through genetic accommodation. If the hypothesis outlined here is correct, then one would expect a positive association between the speciation rates reported in (Venditti et al. 2010) and the robustness of those species to variability within their environments.
Limitations to this proposal
There are limitations to the ideas outlined in this commentary and even if the assumptions hold, the ideas remain incomplete. For instance, acquiring environmental robustness is not necessarily feasible in all biological contexts and is likely restricted to conditions where environmental novelties experienced during a lifecycle are moderate or the occurrence of these environmental novelties is infrequent and spatially differentiated. In circumstances where environments change both frequently and widely (i.e. across all individuals in all demes), the resulting exposure of CGV would be limited by the degree to which CGV has accumulated in addition to the population size and the number of offspring generated in a relatively short span of time and thus one would expect extinction to be a more likely outcome in such scenarios. Under these more volatile conditions, life could probably only persist by rapidly adapting to the environment using anti-robustness combined with high offspring counts. This scatter-gun strategy to adaptation is illustrated by viral evolution where environmental change is broad and antagonistic, viral gene sign epistasis is strongly negative (Sanjuan and Elena 2006), and large numbers of highly distinctive offspring are common. This would suggest that NN-G and NN-E pathways are two of possibly several pathways by which adaptation can be facilitated and, in light of the counter-example just given, might best reflect the evolution of complex species whose niches have become increasingly intertwined with an organism’s flexibility and robustness.
Evolutionary mechanics
Our commentary has also neglected important issues surrounding the underlying mechanics of how biological buffering can facilitate phenotypic novelty and how this novelty is transformed into adaptive innovations, e.g. during development. Recent evidence from computer simulations has indicated that only certain mechanisms that stabilize traits (e.g. degeneracy) will also provide access to distinct and heritable phenotypic variation (Whitacre and Bender 2010; Whitacre 2010a). Related studies have further suggested that the exposure of these novel phenotypes can be adaptive and can originate from either new alleles or new environments (Whitacre et al. 2010; Whitacre 2010b). While there is also interesting evidence of other biological processes that play a role in transforming random perturbations into adaptive innovations (Kirschner and Gerhart 1998), these ideas are not yet understood well enough to be integrated with the ideas presented in this commentary.
Entropic barriers to neutral evolution
As I only briefly summarize the NN-G pathway, I did not discuss contentious points surrounding the idea that adaptations can be facilitated by neutral evolution. One potential problem arises in the time that is required to discover adaptations by searching over a network. In particular, even if a neutral network provides access to many distinct heritable phenotypes, the time required to access these phenotypes and find one that is adaptive could be prohibitively long, resulting in so called entropic barriers to adaptation that take as long or longer to cross than fitness barriers, i.e. adaptation that is inhibited by the need to cross over multiple low-fitness genotypes (van Nimwegen and Crutchfield 2000). While more investigation of these limiting conditions is warranted, one recent simulation study of neutral evolution has indicated that entropic barriers do not necessarily arise when genetic neutrality is created through degeneracy. It has been suggested that this suppression of entropic barriers occurs due to the presence of multiple alternative paths to adaptation that are reachable from different regions of a neutral network (Whitacre et al. 2010).
Concluding remarks
The genetic neutral network (NN-G) model resolves a paradoxical relationship between mutational robustness and the evolvability of species, however, it does not aim to describe or account for differences in evolvability that may arise through the facilitation of ecological opportunities. Here I propose that two species may differ in their propensity for environment-induced adaptation based on difference in their robustness towards variability within their environments. I described an example of this robustness-evolvability relationship using a direct application of the neutral network model, i.e. where the insensitivity of fitness to distinct environments increases access to additional environments with unique and heritable phenotypic consequences.
While the conditions supporting an alternative NN-E pathway appear plausible for evolution in complex environments, my aim was not to propose an alternative model of evolutionary adaptation that is in perfect competition with neutral evolution theory. Instead I briefly consider some of the merits and consequences of a rich evolutionary process that allows for multiple complementary paths toward adaptation. While less elegant than the model I started with, I propose that this integration of contingent biological pathways may provide a more comprehensive understanding of evolution in the myriad contexts in which life is found.
Notes
While genetic diversity is supported by NN-G, it would also be constrained by various factors such as NN-G topology, population size, stochastic effects, mutation-selection balance, etc.
References
Agrawal AA (2001) Phenotypic plasticity in the interactions and evolution of species. Science 294:321–326
Aldana M et al (2007) Robustness and evolvability in genetic regulatory networks. J Theor Biol 245:433–448
Ancel LW, Fontana W (2000) )Plasticity, evolvability, and modularity in RNA. J Exp Zool 288:242–283
Anway M et al (2005) Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 308:1466
Babajide A et al (1997) Neutral networks in protein space: a computational study based on knowledge-based potentials of mean force. Fold Des 2:261–269
Barrett RDH, Schluter D (2008) Adaptation from standing genetic variation. Trends Ecol Evol 23:38–44
Bloom JD et al (2006) Protein stability promotes evolvability. Proc Nat Acad Sci 103:5869
Ciliberti S et al (2007) Innovation and robustness in complex regulatory gene networks. Proc Nat Acad Sci USA 104:13591–13596
Dawkins R (1983) Universal darwinism. In: Bendall DS (ed) Evolution from molecules to man. Cambridge University Press, UK, p 202
Day R et al (2003) Rethinking adaptation. Perspect Biol Med 46:80–95
de Visser J et al (2003) Perspective: evolution and detection of genetic robustness. Evolution 57:1959–1972
Dennett D (1995) Darwin’s dangerous idea. Simon and Schuster, NY
Feduccia A (1999) The origin and evolution of birds. Yale University Press, New Haven, Conn., USA
Gavrilets S (1997) Evolution and speciation on holey adaptive landscapes. Trends Ecol Evol 12:307–312
Gibson G (2008) The environmental contribution to gene expression profiles. Nat Rev Genet 9:575–581
Gibson G, Dworkin I (2004) Uncovering cryptic genetic variation. Nat Rev Genet 5:681–690
Houle D et al (1996) Comparing mutational variabilities. Genetics 143:1467
Jablonka E et al (1995) The adaptive advantage of phenotypic memory in changing environments. Philos Trans: Biol Sci 350:133–141
Jernvall J et al (1996) Molar tooth diversity, disparity, and ecology in cenozoic ungulate radiations. Science 274:1489
Kauffman SA (1993) The origins of order: self-organization and selection in evolution. Oxford University Press, USA
Kirschner M, Gerhart J (1998) Evolvability. Proc Nat Acad Sci USA 95:8420–8427
Laland K, Sterelny K (2006) Perspective: seven reasons (not) to neglect niche construction. Evolution 60(9):1751–1762
Le Rouzic A, Carlborg Ö (2008) Evolutionary potential of hidden genetic variation. Trends Ecol Evol 23:33–37
Lynch M (1988) The rate of polygenic mutation. Genet Res 51:137–148
McGuigan K, Sgrò CM (2009) Evolutionary consequences of cryptic genetic variation. Trends Ecol Evol 24:305–311
Meiklejohn CD, Hartl DL (2002) A single mode of canalization. Trends Ecol Evol 17:468–473
Newman SA (1994) Generic physical mechanisms of tissue morphogenesis: a common basis for development and evolution. J Evol Biol 7:467–488
Odling-Smee F et al (1996) Niche construction. Am Nat 147:641–648
Olivieri I (2009) Alternative mechanisms of range expansion are associated with different changes of evolutionary potential. Trends Ecol Evol 24:289–292
Palmer A (2004) Symmetry breaking and the evolution of development. Science 306:828
Pfennig DW, Wund MA, Snell-Rood EC, Cruickshank T, Schlichting CD, Moczek AP (2010) Phenotypic plasticity's impacts on diversification and speciation. Trends Ecol Evol 25(8):459–467
Pigliucci M et al (2006) Phenotypic plasticity and evolution by genetic assimilation. J Exp Biol 209:2362
Reidys C et al (1997) Generic properties of combinatory maps: neutral networks of RNA secondary structures. Bull Math Biol 59:339–397
Rietkerk M, van de Koppel J (2008) Regular pattern formation in real ecosystems. Trends Ecol Evol 23:169–175
Ruokolainen L et al (2009) Ecological and evolutionary dynamics under coloured environmental variation. Trends Ecol Evol 24:555–563
Sanjuan R, Elena SF (2006) Epistasis correlates to genomic complexity. Proc Nat Acad Sci USA 103:14402
Schlichting CD (2008) Hidden reaction norms, cryptic genetic variation, and evolvability. Ann NY Acad Sci 1133:187–203
Schlichting CD, Pigliucci M (1998) Phenotypic evolution: a reaction norm perspective. Sinauer Sunderland, MA
Schmalhausen I, Dobzhansky T (1949) Factors of evolution: the theory of stabilizing selection. Blakiston Co., Philadelphia
Sultan S (2007) Development in context: the timely emergence of eco-devo. Trends Ecol Evol 22:575–582
Suzuki Y, Nijhout H (2006) Evolution of a polyphenism by genetic accommodation. Science 311:650
Szollosi GJ, Derenyi I (2009) Congruent evolution of genetic and environmental robustness in micro-RNA. Mol Biol Evol 26:867
Uller T (2008) Developmental plasticity and the evolution of parental effects. Trends Ecol Evol 23:432–438
van Nimwegen E, Crutchfield JP (2000) Metastable evolutionary dynamics: crossing fitness barriers or escaping via neutral paths? Bull Math Biol 62:799–848
Venditti C et al (2010) Phylogenies reveal new interpretation of speciation and the red queen. Nature 463:349–352
Waddington CH (1953) Genetic assimilation of an acquired character. Evolution 7:118–126
Waddington CH (1957) The strategy of the genes: a discussion of some aspects of theoretical biology. Allen and Unwin, London
Wagner A (2008a) Robustness and evolvability: a paradox resolved. Proc R Soc Lond B Biol Sci 275:91–100
Wagner A (2008b) Neutralism and selectionism: a network-based reconciliation. Nat Rev Genet 9:965–974
West-Eberhard MJ (2003) Developmental plasticity and evolution. Oxford University Press, USA
West-Eberhard M (2005) Developmental plasticity and the origin of species differences. Proc Nat Acad Sci 102:6543
Whitacre JM (2010a) Degeneracy: a link between evolvability, robustness and complexity in biological systems. Theor Biol Med Model 7:6
Whitacre JM (2010b) Genetic and environment-induced innovation: complementary pathways to adaptive change that are facilitated by degeneracy in multi-agent systems. In Proceedings of the conference on artificial life XII, Odense, Denmark
Whitacre JM, Bender A (2010) Degeneracy: a design principle for achieving robustness and evolvability. J Theor Biol 263:143–153
Whitacre JM et al (2010) The role of degenerate robustness in the evolvability of multi-agent systems in dynamic environments. In 11th international conference on parallel problem solving from nature (PPSN 2010), Krakow, Poland
Wu J, Loucks OL (1995) From balance of nature to hierarchical patch dynamics: a paradigm shift in ecology. Q Rev Biol 70:439
Acknowledgments
I would like to thank G. Gibson, C. Schlichting, A. Harding, and the two anonymous reviewers for their insightful comments and suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Whitacre, J.M. Genetic and environment-induced pathways to innovation: on the possibility of a universal relationship between robustness and adaptation in complex biological systems. Evol Ecol 25, 965–975 (2011). https://doi.org/10.1007/s10682-011-9464-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10682-011-9464-z