
Abstract
Sooty Shearwater (Puffinus griseus) is the most common Procellariiform seabird along the south-eastern South American coast. In recent years the wintering population off California has declined noticeably. This decline has been confirmed on the breeding grounds in New Zealand. In Chile, knowledge of the population is limited. Investigations on Isla Guafo were carried out during two seasons (03/04 and 04/05), beginning an ongoing monitoring and evaluation of the breeding population of this species in southern South America. On Isla Guafo we estimate a population of about 4 million birds that nest above 150 m above sea level (m.a.s.l.) under a forest without understory. Sooty Shearwaters on Isla Guafo prefer north and western slopes that we hypothesise protect them from the predominantly southerly winds. Population of the species from New Zealand and Australia are compared with the Isla Guafo population and the state of its conservation is discussed.
Similar content being viewed by others

Introduction
The Sooty Shearwater (Puffinus griseus) (Procellaridae) is a medium-sized seabird that breeds in burrows on islands in southern South America, Australia and New Zealand. It is the most common Procellariiform seabird in south-eastern South American and New Zealand. It nests between October and May, and migrates to the northern hemisphere during the austral winter (Watson et al. 1971; Warham et al. 1982; Briggs and Chu 1986; Cooper et al. 1991; Spear and Ainley 1999).
Significant declines in the breeding population of this species have recently been reported (Hamilton et al. 1997; Scofield 2001). Similar declines have been recorded during their migration (Scofield and Christie 2002) and on the wintering grounds (Veit et al. 1996; Oedekoven et al. 2001; Hyrenbach and Veit 2003). This species is abundant in the coastal waters surrounding the Chilean Fjords and offshore along in the Cape Horn current during the breeding season and is found throughout the Chilean-Peruvian (Humboldt) current during the post-nuptial migratory period (March–July) (Spear and Ainley 1999). Nevertheless, the numerical and conservation status of these populations is not yet known, in contrast to what is known about populations in New Zealand (Richdale 1963; Hamilton et al. 1997; Jones 2000; Cruz et al. 2001; Uhlman 2003).
Indeed the state of knowledge about seabird populations in southern Chilean (42°–57° S) is precarious. Zoogeographically it is a complex area that can be divided into 2 zones: (1) the fiords and channels and (2) the outer coastal line (Venegas and Sielfeld 1979). The area is divided by the 10°C mean annual isotherm and lies within the temperate and cold sub-zones in Sub-Antarctic seabird life zones. It is also included in the Magellanic district of the Atlantic Province for the Sub-Antarctic region (Watson et al. 1971).
Some basic survey of seabirds have been carried out in this area (Brown et al. 1975; Devillers and Terschuren 1978) but in general, studies on natural or population biology of seabirds between 42° and 51° S are scarce. Although there are several National Parks that have been established for over three decades and cover large areas of terrestrial habitat this is still rather isolated region. The island of Chiloé is of special zoogeographical interest, and is strongly populated. The development of aquaculture affects coastal waters as well as those in the fiords or estuaries (Schlatter 1984).
The objective of this study is to review the state of knowledge of Sooty Shearwaters world wide and update it with new information for the South American populations based on a study of the large Isla Guafo population, which is only the third documented colony in South American waters (Scofield 2001; Lawton et al. 2006).
Methods
Area of study
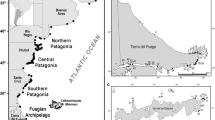
We surveyed Isla Guafo between December and March during 2003/2004 and 2004/2005. Isla Guafo is in Chile’s region X at 43°61′ S; 74°75′ W, of the south-west corner of Isla Grande de Chiloé at the mouth of Golfo de Corcovado (Fig. 1). The colony begins above 150 m above sea level (m.a.s.l.) and burrows are located along the steep slopes formed by numerous mountain ranges. These slopes predominate in the area and flat land is almost absent. Forest species such as Drimys winteri (canelo), Rhaphithamnus spinosus (arrayán macho) and Amomyrtus luma (luma) dominate, whilst shrubs are scarce and patchy, dominated by Myrceugenia planipes (pitrilla). Where shrubs dominate, the vegetation density is high preventing burrow building. The list of plant species and their abundance in colonies is shown in Table 1. Burrows are generally excavated in bare ground (defined here as soil with leave cover, no understory and under a canopy with > 80% tree cover).

Map indicating the geographic location of Guafo Island, Chiloé, Chile, highlighting the reference zones of this study
Population
A population estimate was made by calculating the density of occupied nesting burrows in 20,100 m2 quadrants randomly chosen in the hills between Punta Weather and Caleta Toro (Fig. 3). Not all burrows were active. To discriminate between active and inactive burrows we randomly selected and we marked up to 5 burrows (if possible) for each of the study quadrants to obtain proportion of quadrant occupation. At the entrance of each burrow, 2–3 toothpicks were loosely placed vertically in the entrance forming a barrier each evening and revisited each morning (Bibby et al. 2000). A burrow was considered occupied if all toothpicks were displaced at least once during the study period of at least 5 consecutive days (the minimum period calculated for shearwaters foraging at sea and returning back to the burrow; Warham 1996). In addition, the number of apparent attempts to build burrows (hereafter referred to as burrow attempts) in each quadrant was recorded. A burrow attempt was defined as an apparently freshly excavated cavity less than 20 cm long generally interrupted by arboreal roots. Rats and feral cats also occur on Guafo. Burrow attempts were attributed to Sooty Shearwaters if they left foot tracks, feathers or guano and if they left large amounts of freshly excavated soil. We believe that burrow attempts are probably made by young or inexperienced males learning to burrow and are loosely indicative of the overall activity of a colony (Warham 1990). It was considered that each burrow attempt was undertaken by a different bird. Approximately 7 ha was surveyed observing presence and burrow attempts in the entire zone, although the quadrate work was limited to 3.2 ha due to the rough island geography. Based on the observations of entrance and exit of birds from the burrows, in the areas of Caleta Toro, Caleta Samuel and NE of Punta Norte (Fig. 1) we estimate the colony is distributed all over the island above 150 m.a.s.l. The total area above the 150 m.a.s.l was calculated by means of the Instituto Geográfico Militar (IGM) chart and AutoCADTM 2000 software.
Morphometry
Eight morphometric measurements (using vernier callipers) and mass (using Pesola scales) were taken from 113 individual shearwater captured on the surface at night using netting and in burrows. The measurements taken were: mass, bill (from the tip to the beginning of the feathers), total length of the head (from the tip of the beak to the back of the skull), bill depth (at the base of the bill), bill width (at base), tarsus (metatarsus’s length), wing (from the edge of the carpal plexus to the tip of the longest primary feather) and body circumference (at the proximal edge of the wing). The weight, wing and body circumference plus the other measurements were taken with an accuracy of 10 g, 1 mm, 0.1 mm, respectively. We compared measurements made during the study with data from the literature using correlation coefficients and mean t-tests, respectively.
Use of habitat
Habitat variables were measured using a variety of techniques. Chest height diameter (CHD) of tree trunks was measured using a tree caliper with 1 cm accuracy and trees categorised as thick (>30 cm), medium (20–30 cm), small (<20 cm). Site slope was measured using a SuntoTM clinometer with 1° accuracy. Three measurements were made within each quadrant and the average calculated. The aspect (orientation) was measured using a compass to the nearest degrees parallel to the slope. Multiple regression was use to correlate these variables with the number of occupied burrows and burrow attempts allowing us to evaluate the influence of these variables on site use. This also enabled us to estimate overall habitat use. Regression analysis between burrows and burrow attempts enabled us to examine the influence of birds actually occupying burrows to those that were infer are either immature or not currently breeding. The relationship between occupied burrows, burrow attempts was evaluated by means of a contingency table with orientation as a categorical variable using a non-parametric chi square test.
For a random selection of burrow entrances in each quadrate the maximum width, height and diagonal were measured with a tape to 1 mm accuracy. For each of these randomly chosen burrows the direction of orientation was measured to the nearest degree using a SilvaTM compass. These four variables were correlated to evaluate degree of protection against adverse weather events such as rain and wind using one way ANOVA with burrow orientation as a predictive variable.
To evaluate the degree of burrow aggregation, the distance between a burrow and the closest neighbour was measured using a measuring tape (1 cm accuracy).
Meteorological variables (rainfall, wind intensity and direction, and temperature) taken the Isla Guafo’s weather station in Punta Weather operated by the Armada de Chile were recorded between November 2004 and March 2005. Measurements were carried out every 3 h from midnight. The sequentially sampled No. was 286, 184, 248, 224 and 208, between November and December. The variables considered as dependent were: wind intensity (knots) and temperature (°C). Data were transformed (log10 x + 1) to obtain a normal and homocedastic distribution. It was not possible to carry out a variance analysis (ANOVA/MANOVA) of repeated measurements because the expected sphericity (Mauckly’s test) was violated in all the variables despite the Greenhouse-Geisser correction (Zar 1999). Consequently an ANOVA/MANOVA for each month was carried out separately. These variables were analysed with the wind direction as a factor (categorised as N, S, W and E) with one way MANOVA if at least two variables correlated significantly and with one way ANOVA when the variables were independent among them (Zar 1999). The a posteriori Tukey’s test HSD for unequal N were carried out depending on the case. Wind direction and rainfall (the sum for each month) also categorised as N, S, W and E were compared each month to evaluate their relationships using contingency tables (with chi square).
The statistical analysis was carried out following Zar’s methodology (1999) using the STATISTICA 6.0 software. Data from this study was complemented by a literature review of Sooty Shearwater biology in South American waters.
Results
General information
Sooty Shearwaters were first seen near the island at dusk (ca. 18:00 h) flying around the coastline in great flocks. The first individuals arrive at the breeding grounds were after sunset (ca. 21:00 h) with a peak in arrival on the colony between 22:30 h and 23:30 h. The second peak of activity takes place shortly before sunrise (ca. 06:00 h) whilst it is still mostly dark.
The date of egg laying was not ascertained by us, but eggs hatch during the first 10 days of January. Based on an estimate of 56 days incubation for this species (Richdale 1954) it is assumed on Isla Guafo that incubation begins in the first fortnight of November. Most non-breeding adults appear to leave the colony and the surrounding seas before the 20th of March and only chicks were found in burrows examined on 30th March. It is presumed that chicks leave the colony by the end of April and beginning of May as happens in New Zealand (Warham et al. 1982; Sagar and Horning 1998).
Population estimation
The density of burrow attempts and occupation of burrows is shown in Table 2. The burrow density is low and highly variable (0.050 ± 0.049 burrows/m2). However, the proportion of burrows occupied in Isla Guafo is high (88 ± 30.71%). The burrow attempt density is greater than the burrow density (0.089 ± 0.075 attempts/m2); and we speculate that the non-breeding and pre-breeding proportion of the population is larger than that of breeding birds. The large variation in the number of mean burrow attempts and mean numbers of occupied burrows is probably due to the low number of quadrants and the clustered distribution of burrow with tight clusters of up to 16 burrows within quadrants (Fig. 2).

Distribution of the number of occupied burrows (black bars) and burrow attempts (white bars) per quadrant (n = 20), in four intervals of density
The total estimated area above the 150 m.a.s.l. was 84.411 km2. The population of birds occupying burrows based on the densities in the 20 sampling quadrants was 3,980,000 ± 3,870,000 pairs that were breeding and/or couples starting to reproduce (Fig. 3).

Nesting Area considered for Sooty Shearwater in Guafo Island, Chile. The black areas represent the total surface above the 150 m.a.s.l. The arrow point out a white ellipse that represents the area with density quadrants used in this study
Morphometry
The morphometric measurements for Sooty Shearwaters on Isla Guafo are shown in Table 3. There were no significant differences between the morphometric variables considered among the individuals captured in burrows and those captured on surface therefore, the data was treated as a single unit (t weight = − 0.13, P = 0.894; t culmen = 0.519, P = 0.605; t head length = 1.675, P = 0.097; t peak depth = 0.465, P = 0.643; t peak width = 0.502, P = 0.617; t circumference = 1.039, P = 0.302; t tarsus = 0.328, P = 0.744; t wing = 0.259, P = 0.796, d.f. = 86). A comparison of means between our results and those reported by Richdale (1963) for Whero Island (46°55′ S), New Zealand was carried out. This is unfortunately the only other work showing standard deviations. T-tests of means show significant differences between weight (t = 63.03), culmen (t = 5.73), peak depth (t = 213.36), beak width (t = 334.94) and wing (t = 10.31); t (2)0.001 = 3.34, d.f. = 211. All the measures published by Richdale (1963) are greater than the measurement of Isla Guafo Sooty Shearwaters, except the culmen size. Correlation values for the morphometric variables of Sooty Shearwaters from Isla Guafo (Table 4) show weak but significant correlation coefficients among them.
Use of macrohabitat
No significant relationship between the number of occupied burrows and burrow attempts per quadrant were found (R 2 = 0.103, F = 2.056, d.f. = 1,18, P = 0.169) (Fig. 2). Similarly, no significant relationships was observed between the hill slope inclination and the number of burrows (R 2 = 0.007, F = 0.137, d.f. = 1,18, P = 0.715) and burrow attempts (R 2 = 0.024, F = 0.438, d.f. = 1,18, P = 0.516), although they tend to be located between 19° and 37° of inclination (Fig. 4). The number of occupied burrows and burrow attempts is greater in the areas oriented N and W (Fig. 5). There is no relationship between the chest height diameter of the trees and the number of burrows per quadrant (without tree categories) (R 2 = 0.04, F = 0.749, d.f. = 1,18, P = 0.398) and for each group of dimension (R 2 multiple = 0.173, F = 1.115, d.f. = 3,16, P = 0.372) and neither between the same variables and burrow attempts (R 2 = 0.00, F = 0.086, d.f. = 1,18, P = 0.772 and R 2 multiple = 0.237, F = 1.662, d.f. = 3,16, P = 0.215, respectively). The frequency of occurrence of occupied burrows and attempts are distributed independently of the number of trees in each category (Fig. 6).

Relationship between the island’s slope inclination and the frequency of occurrence of occupied burrows (white circles) and burrow attempts (black circles)

Occurrence–frequency of occupied burrows and burrow attempts according to exposure of slopes selected as macro-habitat

Distribution of the occurrence–frequencies of occupied burrows and attempts, and number of trees for each category at chest height diameter: large (white circle), medium (black circle), small (black square)
Use of microhabitat
The mean height, width and diagonal of burrow entrances was 13.5 ± 4.0, 23.6 ± 7.5 and 22.0 ± 5.8 cm, respectively (n = 78) and there is no correlation between the width and the height (R = 0.086, P = 0.455). However, as expected, we found correlation between width, height and the diagonal (R = 0.242, P = 0.03 for height; R = 0.689, P = 0.000, for width) because the diagonal is a measurement that is proportional to width and height. There are no significant differences between the burrow exposure and the variables height, width and diagonal (Table 5). There is a tendency of the birds to build their burrows with entrances oriented towards the N and W (65% of the cases) and this conforms to the observed pattern of higher density of occupied burrows with greater slope exposure (Figs. 5 and 7). The burrow aggregations, measured with the distance between burrows show that 38% of the burrows are less than 1.33 m apart and 81% of them were less than 3.9 m apart. This indicates that Sooty Shearwaters show a degree of self-cohesion for the selection and construction of burrows at certain sites (Fig. 8).

Percentage of occurrence for the burrow entrances orientation (n = 78). X-axis indicates the orientation groups: 315–45° (N), 45–135° (E), 135–225° (S), and 225–315° (W)

Distance to the nearest neighbour burrow expressed as % of occurrence (n = 78)
Climatic conditions
Monthly rainfalls totalled 53.5, 89.1, 80.9, 27.5, 121.5 mm, between November and March, respectively. Thus rainfall during the study was lowest in November and February and highest in March, mainly during northerly and westerlies winds (Fig. 9). From the total observations for November, January and February almost 50–70% were dominated by southerly winds followed by an alternation of winds from the northerly and westerly directions. During December and March the northerly winds dominated (Fig. 9).

Rainfall frequencies (mm), winds intensity (knots) and T° (°C) during November and December 2004/2005. The series correspond to N, W, S and E. The series of the other variables correspond to mean and SD. Different letters indicate significant differences (P < 0.05) according to Tukey’s test HSD for unequal N
There were significant differences between the variables: wind intensity and temperature related to wind direction during each month (Table 6). There was no significant difference in wind intensity in November and December, but was significantly different during the three remaining months (Fig. 9). During January and February, the greatest wind intensities came from the south, whereas in March came from the west and south. There were significant differences in the temperature between months and these were lower during southerly winds (Fig. 9).
Discussion
Life cycle
Data indicate that in general the lifecycle of Sooty Shearwater on Isla Guafo was very similar to that on colonies in New Zealand (Richdale 1954, 1963; Marchant and Higgins 1990; Everett and Pitman 1993; Warham 1996; Onley and Bartle 1999; Spear and Ainley 1999), although the date non-breeder depart from the colony in southern Chilean seems be earlier than that on Whero Island (New Zealand; Richdale (1963)). We suggest this may be due to the increase in rainfall from March onwards in this area (Fig. 9).
Population size
Our population estimate for Isla Guafo is significantly greater than that reported by Clark et al. (1984b) of 200,000 individuals. Although the density of burrows in the Isla Guafo colony is quite low (Figs. 2, 3 and 6; Table 2) compared with those in New Zealand (sometime < 2 burrows/m2; Warham 1990), the population estimate is huge due to the extensive area available for nesting. The heterogeneous nature of the distribution of occupied burrows has caused a large standard deviation and thus wide confidence intervals on our estimate of population size. We found 10% of quadrates had no burrows whilst 70% of the quadrants that had two or more occupied burrows. Furthermore neighbour distance distribution shows that burrows appear to be clustered (Fig. 8).
There is anecdotal evidence that other even larger islands in the southern Chilean Fjords sooty colonies contain populations of Sooty Shearwaters thus the population of this species in South America should be larger than the population reported by Robertson and Bell (1984) for New Zealand. Agreeing with the assumption of Scofield (2001) that Chile may contain the majority of the world’s population of Sooty Shearwaters. Indeed we find that if our estimate is accurate that this is in fact the largest seabird colony in the world. It is rivalled only by the population of Sooty Shearwaters on The Snares (New Zealand) of 2.75 million burrows (Warham and Wilson 1982).
Sooty Shearwaters occur along the coast of New Zealand in a large number of small colonies of between 2 and 2,100 birds on and in large colonies on surrounding islands (Hamilton et al. 1997). It is estimated that there are a total of 20 million pairs in the New Zealand region (Robertson and Bell 1984) Thus it is one of the most abundant seabird species in the Pacific Ocean (Marchant and Higgins 1990). There are also small colonies in southern Australia with approximately 2,000 birds in the largest (Lane and White 1983 and Brothers 1979 cited in Spear and Ainley 1999).
Relationship between populations
Our morphometric data support the finding of Baduini and Warheit (in preparation) that have found that the population of Isla Guafo is genetically different from the populations of New Zealand, Valparaiso (Chile) and Monterrey Bay (Californian Current). We suggest that this difference could explain the Bergman’s rule (Scholander 1955) for this species, between populations of Whero Island, New Zealand, and Isla Guafo, Chile.
Factors affected distribution
Activity of the non-breeders (as witnessed by burrowing activity late in the breeding season) appears not be correlated to the density of breeding burrows (Fig. 5) suggesting that non-breeders are not attract by the presence of breeding birds.
Sooty Shearwaters breeding on Guafo prefer steep hill-slopes (Fig. 3). We suggest this may be in response to better drainage due to the heavy fogs and to a lesser extent due to the occasional heavy rainfalls during summer. Landscapes with slopes offer advantages for nesting tubenosed birds (Schramm 1986; Brandt et al. 1995; Catry et al. 2003). The density of birds is not correlated to the presence of a dense forest upperstory. In some cases birds occupy areas with no forest-understory but with a leave covered floor. Alternatively rainfall might have little importance in choosing nesting sites since soils formation is dependent on drainage and soil stability (Warham and Wilson 1982; Jones 2000). Neither collapse nor burrow flooding were observed on the island thus reflecting stability and good drainage of soils.
On Guafo birds tend to locate their burrows on N and W slopes (Fig. 7). During the breeding season winds from the south are frequent and strong bringing dry conditions. During the same period, rainy is more frequent during northerlies and (to a lesser extent) during westerlies (Fig. 9). We suggest therefore that Sooty Shearwaters chose nest site based on there protection against prevalent winds. Exactly why Sooty Shearwaters would make such habitat choices is unclear. Lyver (2002) established that environment in burrows is comparatively uniform due to the isolation from the climatic effects on the surface. Our study suggests that winds influence of burrow location may require further study. The size of burrow entrances did not seem to have been a factor affecting the burrow protection from adverse climatic events as there was no relationship between burrow height width and diagonal in relation to burrow orientation (Table 5).
State of the knowledge for Sooty Shearwater in South America
In South America and particularly in Chile, there is limited information about the populations of this species (Reynolds 1935; Watson et al. 1971; Jehl 1973; Brown et al. 1975; Devillers and Terschuren 1978; Schlatter 1984; Clark et al. 1984a, b, Lawton et al. 2006; Scofield 2001). The first records of nesting without population estimation are from Wollaston and Bayly islands (Reynolds 1935; Watson et al. 1971). Marín (1984) reported 21 burrows with eggs or chicks in the small island Puñihuil, Chiloé, (41°55′ S) located north of Guafo Island. No evidences of nesting has been reported recently there (A. Simeone, personal communication) as a consequence of the increasing tourist activities. Brown et al. (1975) reported that this species is common in Chilean fiords and scarce in the Atlantic Ocean and Drake Passage. It is rarely seen south of the Antarctic Convergence on the South American quadrant as opposed to the New Zealand one (Watson et al. 1971). Jehl (1973) established that Sooty Shearwater is absent in the inner seas during the austral winter. If we compare the abundance distribution of the sooties between the Strait of Magallanes and the central-south zone of Chile (40° S), reported by Jehl (1973) (May), Brown et al. (1975) (March) and Devillers and Terschuren (1978) (January) it is possible to observe an increase in the amount of Sooty Shearwater records towards the north as the summer season moves into autumn-winter. This variation is the result of the migratory process of this species towards the Northern Hemisphere although a small fraction of the population (probably non-breeders) remains in austral waters during winter (Jehl 1973).
Scofield (2001) considers 300,000 birds breed in the Wollaston Islands. Clark et al. (1984b) and Schlatter (personal observations) presume the nesting in Isla Guamblin (45° S) close to and south of Isla Guafo. For the archipelago Diego Ramírez (56°31′ S, 68°44′ W) a population of several thousand pairs (Lawton et al. 2006) have been estimated. Their also occur on Isla Ildefonso (55°50′ S, 69°19′ W) thought no population estimation has been made (Kieran Lawton, personal communication). In the Argentinean Patagonia, there are colonies with about 10,000–20,000 pairs on Las Malvinas (50°59′ S, 61°26′ W) (Leandro Tamini, personal communication). Nesting has been reported on Staten Island east of Tierra del Fuego (54°44′ S, 63°52′ W), by Clark (1986) but more recently discarded by Schiavini et al. (1999). This island needs further survey to confirming Sooty Shearwaters breeding status.
The populations trends of the Sooty Shearwater populations in Chile is unknown. On the Wollaston Islands Scofield (2001) reported a series of introduced predators and sheep that would interact negatively with colonies of shearwaters on these islands. On Guafo Island, the presence of rodents in the colony was evident, during all sample season. Two feral cats was also observed. Most mortality probably mainly by the Crested Caracara Polyborus plancus. Thus Sooty Shearwater populations breeding in Chile are under most of the same population pressures than those breeding in New Zealand, where the introduction of predators is considered one of the main causes of the population decreases (Hamilton et al. 1997; Jones 2000; Scofield and Christie 2002; Valenzuela and Grau 2005 for Chile).
Observations suggest Sooty Shearwaters forage mainly inshore and as far as he continental shelf off Guafo. The prey consumed by Sooty in South American waters is still unknown although Cruz et al. (2001) described a wide range of prey for New Zealand sooty populations. The stomach content of two individuals from the Guafo showed fish remains and a small amount of squid.
Uhlman (2003) has summarised incidence of fisheries bycatch of Sooty Shearwater. The interaction of Sooty Shearwaters with fisheries along southern South America is poorly unknown. In Peru there are no records of captures (Jeffrey Mangel, personal communication) and in Argentina, although local fishermen report their capture, data is lacking (Leandro Tamini, personal communication). There are only occasional reports of drowned birds coming ashore south of Concepción (36°30′ S) and near Valdivia (39°40′ S) as a result of the interaction with fisheries using fence nets or artisan fisheries. A very low rate of bycatch (0.005 birds/1,000 hooks) is reported appears with in the Dissostichus eleginoides long-line fishery (Moreno et al. 2003).
The Sooty Shearwater has recently been upgraded to “Near Threatened” (IUCN 2006) due to the decrease in the abundance of the species in the California current system (Veit et al. 1996); a decline in populations and colonies in New Zealand (Hamilton et al. 1997). This potentially in the colonies of the austral Chilean archipelagos as a result of the introduction of predators and the interaction with fisheries. In addition, climatic phenomena like El Niño, the global warming, and their relation with the distribution of preys in the ocean, would significantly affect the survival of these birds (Veit et al. 1996; Lyver et al. 1999).
To establish the effect of fisheries on this species in Chile it is essential to evaluate the population size in South America. Some data indicate that trawling fisheries of austral hake (Merluccius australis) could be interacting negatively with the populations of this pelagic seabird predator during the reproductive period in Chile (Javier Arata, personal communication). Gillnets have been demonstrated to be dangerous for marine bird species such as the Humboldt penguin (Spheniscus humboldti) (Simeone et al. 1999) and pink-footed shearwater (P. creatopus) (Schlatter 2004) and thus Sooty Shearwaters may be vulnerable as well. As part of this study we have begun to evaluate the survival and fecundity of this species on Guafo Island as a way to complement further studies on the interaction of Sooty Shearwaters with factors like fisheries and introduced predators.
References
Araya B, Millie G (1986) Guía de Campo de las Aves de Chile; Séptima Edición. Editorial Universitaria, Santiago de Chile
Bibby CJ, Burgess ND, David AH, Mustoe SH (2000) Bird census techniques, 2nd edn. Academic Press, London, 385 pp
Brandt CA, Parrish JK, Hodges CN (1995) Predictive approaches to habitat quantification: dark-rumped Petrels on Haleakala, Maui. Auk 112:571–579
Briggs KT, Chu EW (1986) Sooty shearwaters off California: distribution, abundance and habitat use. Condor 88:355–364
Brown RGB, Cooke F, Kinnear PK, Mills EL (1975) Summer seabird distributions in Drake Passage, the Chilean fjords and off southern America. Ibis 117:339–356
Catry P, Campos A, Segurado P, Silva M, Strange I (2003) Population census and nesting habitat selection of thin-billed prion Pachyptila belcheri on New Island, Falkland Islands. Polar Biol 26:202–207
Clark R (1986) Aves de Tierra del Fuego y Cabo de Hornos, guía de campo. L.O.L.A. Buenos Aires, Argentina, 294 pp
Clark GS, Von Meyer AP, Nelson JW, Watt JN (1984a) Notes on the Sooty shearwater and other avifauna of the Chilean offshore island of Guafo. Notornis 31:225–231
Clark GS, Goodwin AJ, Von Meyer AP (1984b) Extension of known range of some seabirds on the costal of southern Chile. Notornis 31:320–324
Cooper J, Underhill LG, Avery G (1991) Primary moult and transequatorial migration of the sooty shearwater. Condor 93:724–730
Cruz JB, Lalas C, Jillett JB, Kitson JC, Lyver POB, Imber M, Newman JE, Moller H (2001) Prey spectrum of breeding sooty shearwater (Puffinus griseus) in New Zealand. New Zeal J Mar Fresh 35:817–829
Devillers P, Terschuren JA (1978) Midsummer seabird distribution in the Chilean fjords. Le Gerfaut 68:577–588
Everett WT, Pitman RL (1993) Status and conservation of shearwater of the North Pacific. In: Vermeer K, Briggs KT, Morgan KH, Siegel-Causey D (eds) The status, ecology, and conservation of marine birds of the North Pacific. Can. Wildl. Serv. Spec. Publ., Ottawa, pp 93–100
Falla RA, Sibson RB, Turbott EG (1979) The new guide to the birds of New Zealand. New Zealand Ornithological Society, Auckland, New Zealand, 247 pp
Hamilton SA, Moller H, Robertson CJR (1997) Distribution of Sooty shearwater (Puffinus griseus) breeding colonies along the Otago Coast, New Zealand, with indication of countrywide populations trend. Notornis 44:15–25
Harrison P (1983) Seabirds – an identification guide. Croom Helm, London, Sidney
Harrison P (1987) Seabirds of the world – a photographic guide. Christopher Helm Ltd., London
Hyrenbach KD, Veit RR (2003) Ocean warming and seabird communities of the southern California Current System (1987–98): response at multiple temporal scales. Deep-Sea Res Pt II 50:2537–2565
IUCN (2006) IUCN Red List of threatened species Categories. www.redlist.net
Jehl JR (1973) The distribution of marine birds in Chilean waters in winter. Auk 90:114–135
Jones C (2000) Sooty shearwater (Puffinus griseus) breeding colonies on mainland South Island, New Zealand: evidence of decline and predictors of persistence. New Zeal J Zool 27:327–334
Lawton K, Robertson G, Kirkwood R, Valencia J, Schlatter R, Smith D (2006) An estimate of population sizes of burrowing seabirds at the Diego Ramírez archipelago, Chile, using distance sampling and burrow-scoping. Polar Biol 29(3):229–238
Lyver POB (2002) Use of traditional knowledge by Rakiura Maori to guide sooty shearwater harvests. Wildlife Soc B 30:29–40
Lyver POB, Moller H, Thompson C (1999) Changes in the sooty shearwater Puffinus griseus chick production and harvest rate precede ENSO events. Mar Ecol Prog Ser 188:237–248
Marchant S, Higgin PJ (eds) (1990) Handbook of Australian, New Zealand, and Antarctic Birds, vol 1. Oxford University Press, Melbourne, 1400 pp
Marín M (1984) Breeding record for the Sooty Shearwater (Puffinus griseus) from Chiloé Island, Chile. Auk 101:192
Moreno CA, Hucke-Gaete R, Arata J (2003) Interacción de la pesquería del bacalao de profundidad con mamíferos y aves marinas. Informe final proyecto FIP No. 2001-31. República de Chile, Ministerio de Economía, Fomento y Reconstrucción, Subsecretaría de Pesca, Fondo de Investigación Pesquera,199 pp
Murphy RC (1936) Oceanic birds of South America, vol II. The American Museum of Natural History, New York
Narosky T, Yzurieta D (2003) Aves de Argentina y Uruguay: guía para la identificación: decimoquinta edición. Vazquez Mazzini editores, Buenos Aires Argentina, 364 pp
Oedekoven CS, Ainley DG, Spear LB (2001) Variable responses of seabirds to change in marine climate: California Current, 1985–1994. Mar Ecol Prog Ser 212:265–281
Onley D, Bartle S (1999) Identificación de las aves marinas de los Océanos del Sur. Te Papa Press, Wellington New Zealand, 83 pp
Peterson RT, Chalif EL (1989) Aves de México. Editorial Diana, México, 473 pp
Reynolds PW (1935) Notes of the birds of Cape Horn. Ibis 5:65–101
Richdale LE (1954) Duration of parental attentiveness in the sooty shearwater. Ibis 96:586–600
Richdale LE (1963) Biology of Sooty shearwater Puffinus griseus. P Zool Soc Lond 141:1–117
Robbins Ch, Bruun B, Zim H (1966) Birds of North America: a guide to field identification. Goldern Press, New York
Robertson CJR, Bell BD (1984) Seabird status and conservation in the New Zealand region. In: Croxall JP, Evans PGH, Schreiber RW (eds) Status and conservation of the world’s seabirds. Inst Counc for Bird Press. Tech. Pub. No. 2, England, pp 573–586
Sagar PM, Horning DS Jr (1998) Mass-related survival of fledging Sooty Shearwaters Puffinus griseus at the Snares, New Zealand. Ibis 140:329–339
Schiavini A, Frere E, Yorio P, Parera A (1999) Las aves marinas de la Isla de los Estados, Tierra del Fuego, Argentina: revisión histórica, estado poblacional y problemas de conservación. Ans Inst Pat Punta Arenas (Chile) 27:25–40
Schlatter R (1984) The status and conservation of seabirds in Chile. In: Croxall JP, Evans PGH, Schreiber RW (eds) Status and conservation of the world’s seabirds. Inst Counc for Bird Press. Tech. Pub. No. 2, England, pp 261–269
Schlatter R (2004) Fardelas en peligro. Cartas al director Diario El Sur de Concepción, Chile 28/01/04. elsur.cl
Scholander PF (1955) Evolution of climatic adaptation in homeotherms. Evolution 9:15–26
Schramm M (1986) Burrow densities and nest site preferences of petrels (Procellarian) at the Prince Edward Islands. Polar Biol 6:63–70
Scofield P (2001) Status of sooty shearwater, Puffinus griseus, in the Wollaston and Hermite groups of southern Chile and concerns for the conservation status of Tierra del Fuego Islands. Unpublished document, 15 pp
Scofield P, Christie D (2002) Beach patrol records indicate a substantial decline in sooty shearwater (Puffinus griseus) numbers. Notornis 49:158–165
Simeone A, Bernal M, Meza J (1999) Incidental mortality of Humboldt penguins Spheniscus humboldti in gill nets, central Chile. Mar Ornithol 27:157–161
Spear LB, Ainley DG (1999) Migration routes of sooty shearwater in the Pacific Ocean. Condor 101:205–218
Uhlman S (2003) Fisheries bycatch mortalities of sooty shearwaters (Puffinus griseus) and short-tailed shearwaters (P. tenuirostris). Doc. Science Internal Series 92. Department of Conservation, Wellington, 52 pp
Valenzuela JA, Grau JH (2005) Ocurrence of American mink on the Chonos Archipelago of southern Chile. Oryx 39:15
Veit RR, Pyle P, Mcgowan A (1996) Oceanic warming and long-term change in pelagic bird abundance within the California current system. Mar Ecol Prog Ser 139:11–18
Venegas C, Sielfeld W (1979) Antecedentes para la determinación de un nuevo distrito zoo-geográfico en el litoral exterior de Magallanes. Ans Inst Pat Punta Arenas (Chile) 10:201–208
Warham J (1990) The Petrels, their ecology and breeding systems. Academic Press Limited, United States, 440 pp
Warham J (1996) The behavior, population biology and physiology of the petrels. Academic Press, New York, 611 pp
Warham J, Wilson GJ (1982) The size of the Sooty shearwater population at the Snares islands; New Zealand. Notornis 29:23–30
Warham J, Wilson GJ, Keeley BR (1982) The annual cycle of Sooty shearwater Puffinus griseus at the Snares Islands, New Zealand. Notornis 29:269–292
Watson GE, Angle JP, Harper PC, Bridge MA, Schlatter RP, Tickell WLN, Boyd JC, Boyd MM (1971) Birds of the Antarctic and Subantarctic. Antarctic Map Folio Series 14. American geographical Society, NY
Zar JH (1999) Biostatistical analysis. Prentice Hall Inc., Upper Saddle River, NJ
Acknowledgements
The authors wish to thank the Dirección de Extensión, Universidad Austral de Chile (Valdivia, Chile) and The Claremont College (Claremont CA, USA) for partially funding this study and to the Armada de Chile for the logistic support. Thanks are given to Javier Arata, Daniela Guicking, Jeffrey Mangel, Henrik Moller, Kieran Lawton, Philip Lyver, Leandro Tamini, Alejandro Simeone, Paul Scofield and Jorge Valenzuela for providing information. We would also like to thank Paul Scofield for their values comments and reviews on this manuscript. To Cesar Barrales for his help in the field. Ronnie Reyes-Arriagada Ph.D. student of the Ph.D. in Science Programme, on Systematic and Ecology, UACH, is being supported by MECESUP Higher Education Programme (UCO 0214).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Reyes-Arriagada, R., Campos-Ellwanger, P., Schlatter, R.P. et al. Sooty Shearwater (Puffinus griseus) on Guafo Island: the largest seabird colony in the world?. Biodivers Conserv 16, 913–930 (2007). https://doi.org/10.1007/s10531-006-9087-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-006-9087-9