
Abstract
Sand dunes in China are widely re-vegetated with Mongolian pine (Pinus sylvestris var. mongolica). However, the effects of precipitation and planting density on population dynamics of this tree species are not well known. We established a system dynamics (SD) model for Mongolian pine to explore the effects of precipitation and initial planting density on population dynamics. This study applied the SD model in simulating the dynamics of a plant population with random environmental factors. The results in this study revealed that the SD model performed well in reflecting the dynamics of Mongolian pine population in the Horqin Sandy Land (ARE < 0.13, MARE = 0.048). Population size fluctuated in the range of 1150–1350 individuals ha−1 under mean annual precipitation of 500 mm and planting density of 10,000 individuals ha−1, suggesting that this range was the fittest density of Mongolian pine population at that precipitation level. Population size varied with precipitation, indicating that water supply played an important role in determining the dynamics of Mongolian pine population. Initial plantation density did not influence population size in scenarios with high precipitation but it influenced population size under low precipitation. This suggested that initial plantation density was more important under low than under high water supply used to support the development of Mongolian pine population. These results are important for formulating guiding principles for the management of Mongolian pine populations in the Horqin Sandy Land.
Similar content being viewed by others
Introduction
Mongolian pine (Pinus sylvestris var. mongolica) is a tree species important in afforestation efforts especially in sandy areas in China (Jiao 1989; Zhu et al. 2003; Zheng et al. 2012). Because it has high tolerance to drought and soil infertility, Mongolian pine plays an important role in fixing sandy land and reducing soil desertification (Zeng et al. 1996; Wang et al. 2012). The area of Mongolian pine plantation forests reached more than 30,000 ha in sandy lands (Kang et al. 2004). Mongolian pine populations in large areas of sandy lands require large amounts of water, satisfied mainly by natural precipitation. Precipitation could influence Mongolian pine in aspects of aboveground biomass accumulation and seed germination (Zhu et al. 2005; Wang et al. 2012). Therefore, precipitation plays a vital role in regulating the dynamics of Mongolian pine populations (Zhu et al. 2006).
The effects of precipitation on population dynamics of Mongolian pine are not well known, although the influence of precipitation on individual growth and species interactions has been reported (Fan et al. 2014; Farrer et al. 2014; Lendvay and Kalapos 2014). The main reason for insufficient information is that the precipitation is critical but its dynamics are random, causing uncertainty when predicting the dynamics of a population. The dynamics of a population can be described with population models, which play an important role in predicting how populations might respond to changes in the environment (Freckleton et al. 2011), and are therefore essential tools in ecology (May 1975; Ellis and Crone 2013).
A widely used model describing the dynamics of a single-species population is the logistic model, proposed by Verhulst (Rockwood 2006). In this model, the rate of population growth is predicted by dN/dt = rN(1 − N/K), where N is the population size, r is the intrinsic rate of natural increase, and K is the carrying capacity of the environment.
The logistic model bases on several assumptions. For example, carry capacity is a constant and population growth is not affected by the age distribution (Rockwood 2006). These assumptions have been gradually overcome in the following studies. Smith (1963) divided the requirement of resources into the reproductive and survival parts. Cui and Lawson (1982) revised the intrinsic growth rate with a function of gender, age, and resource limitation. Recently, more complex models are commonly employed, such as the Leslie and Lefkovitch’s matrix models, which consider age- or stage-structured populations, and integral projection models, which consider the effects of size on population growth and survival (Easterling et al. 2000; Ellner and Rees 2006; Freckleton et al. 2011).
The age- or structured population models are useful tools for basic research on the dynamics of Mongolian pine populations, whose life history is more than 150 years old (Jiao 1985). The age- or structured population models induce a matrix into the differential equations and make the calculation more complex compared with that in the logistic model. To simplify the calculation, a system dynamics (SD) model is used to simulate the dynamics of Mongolian pine population. The SD model is a useful tool to simulate staged models, as transition probability matrix is included in intrinsic modules and can be dealt with simple parameters (McKnight and Finkel 2013). The SD model is widely used in scenario simulation, especially in complex ecosystem. For example, the SD model is used to simulate the conversion process of nitrogen and to verify the threshold value of regional eco-security (Xu et al. 2008; Li and Cai 2010).
The Horqin Sandy Land (42°15′N–45°47′N, 119°15′E–123°45′E) is located in the northeastern part of China with the area of 51,000 km2 (Tang et al. 2014). It is one of the most important distribution zones of Mongolian pine (Kang et al. 2004). A climatic warming and drying trend in the Horqin Sandy Land was observed during the last 50 years (Sun et al. 2005). For example, the annual precipitation days decreased at a rate of 1.3 days × 10 a−1 (Qu et al. 2009). Therefore, the change in plant water supply is essential for predicting the balance between population size and water supply, and should be reflected in the SD model.
In addition to precipitation levels, initial plantation density influences plant population in aspects of individual biomass (Liu et al. 2013), seed production (Li et al. 2012), and biological traits (Rui et al. 2011). Therefore, the effects of initial density of Mongolian pine need to be considered in modeling efforts. Exploring the effects of precipitation and initial plant density is helpful for guiding afforestation programs of Mongolian pine in semi-arid lands.
The main aim of this study was to estimate the effects of precipitation and initial plantation density on population size of Mongolian pine with the use of system dynamics model. To that end, we validated the use of the model for our application, evaluated stability of the population size, and simulated populations of Mongolian pine under 13 combinations of precipitation and initial planting density. The results of this study can serve as a guide for afforestation efforts with Mongolian pine and provide an example of using an SD model to simulate plant population dynamics for an age-structured population with randomly distributed environmental factors.
Materials and methods
Building the system dynamics model
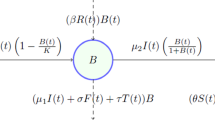
The system dynamics model used here included three parts: (1) a population growth model; (2) a water supply model; and (3) the transition probability matrix, i.e. the set of probabilities from one age stage to another age stage, age stage being young, half-mature, near-mature, mature and over-mature forest. This population growth model was based on a revised logistic model, i.e. the intrinsic rate in the logistic model was divided into birth and death rates (Zhou and Wang 2002). Birth and death rates are defined below. This model followed Eq. (1),
where N was the population size, D was death rate, i.e. the number of deaths individuals per 1 individual in a stage, TP was the transition probability (i.e. probability associated with a pine population at one age-stage shifting to another age-stage), i indicated stages from 1 to 5, B was birth rate, i.e. the function of natural birth rate and water supply limitation (see Eq. 2 and explanations below).
where r i was the natural birth rate in stage i, defined as the number of new seedlings per 1 individual within a stage, N i was the population size in stage i, K i was the maximum number of individuals supported by water supply in stage i, i.e. K i = ws i /c i , ws i was water supply in stage i, and c i was water consumed per 1 individual in stage i.
The water supply for Mongolian pine is a proportion of the precipitation, as Mongolian pine competes for water supply with other plants in sandy lands. The allocation proportion of water supply between Mongolian pine and other plants was observed in Horqin Sandy Land (Zeng and Jiang 1995). Therefore, the water supply for Mongolian pine as the product of precipitation (p) and the allocation proportion (ap), i.e. ws = p × ap. Precipitation followed Gaussian distribution with two parameters, i.e., expectation (E) and variation (V). The transition probability was the uniform distribution and probability density function as follows,
where T represented the duration of a stage. In addition, we used the ‘limit index’ to assign 1 − N i /K i in Eq. 2.
Parameter estimation
The data used to estimate the parameters and to calibrate the model were obtained in the Zhanggutai region (42°35′N–42°47′N, 122°23′E–122°40′E, altitude 226.5 m), located in the southern part of the Horqin Sandy Land, where Mongolian pine was planted from the 1950s to present (Jiao 2001). This region features typical semi-arid climatic conditions (Zheng et al. 2012). We obtained 61 years of annual precipitation data (1953–2013) from the China Meteorological Data Sharing Service System (http://data.cma.cn) to estimate the E and V of the precipitation distribution. The expectation of precipitation distribution is the mean annual precipitation in this region, i.e. 500 mm.
The five stages of Mongolian pine populations were defined based on age ranges of the forest, i.e. young: 1–20 years, half-mature: 21–30 years, near-mature: 31–40 years, mature: 41–60 years, and over-mature forest: 61–150 years old (Kang 2001). Water supply for the young forest and for the other stages was 30.58 and 51.03 % of the precipitation respectively, as reported in a previous study (Zeng and Jiang 1995). Water consumed by an individual in 1 year was 1166, 2684, 4493, 4493, and 4493 kg for the five stages, respectively (Zeng and Jiang 1995). Data for water consumed by an individual in mature and over mature forests were collected in the near-mature forest. The natural birth and death rates of Mongolian pine in different stages were calculated following the life table of Picea mongolica populations (Zheng et al. 1997), except for the death rate in the young forest, which was calculated from actual measurements (Yi et al. 2006). Picea mongolica is a species in the same family as Mongolian pine. The normal initial density was 1 m × 1 m in vertical spacing, or 10,000 individuals ha−1 (Alamusa and Jiang 2008). The values of parameters inputted in the SD model are showed in Table 1.
A SD simulation model
The model was formulated and simulated using a professional SD software package ‘Ventana Simulation Environment Personal Learning Edition (Vensim PLE)’. The simulation was run for a period of 1000 years, and the modeling step was 1 year. This model consisted of five sub-systems, corresponding to the five forest stages, i.e. young (stage 1), half-mature (stage 2), near-mature (stage 3), mature (stage 4), and over-mature forest (stage 5) (Fig. 1).

A stock-flow diagram representing the simulated system
Model validation
We used unit-consistency and quantitative tests to validate this model. The unit-consistency test, which checks for agreement among units, was automatically completed in the Vensim package. The quantitative tests analyzed whether the results of model simulation were consistent with real population dynamics. The real population dynamics were reflected by population size, determined with data collected in plots (Jiao 1985). Absolute percentage error (APE) and mean absolute percentage error (MAPE) were employed for model performance validation (Wei et al. 2012).
where Y t ′ and Y t represent simulated and observed results, t is time unit, n is the number of age-stages (n = 5).
Sensitivity analysis
We selected initial population density and mean annual precipitation for sensitivity analysis. We used a method in which the value of one parameter was changed while the values of the other parameters were kept constant. The sensitivity index (S yi ) was calculated using the following Eqs. (6):
where Y t and X t were population size and parameter value at simulated run time t (t = 1000 year), respectively. dY t and dX t were the values for a change of population size and parameter at time t. The mean sensitivity degree index (S) was calculated following Eq. (7), where n was the number of age-stages at time t (n = 5) (Wei et al. 2012).
Scenario analysis
Four types of precipitation years were considered, including an extreme drought (25 % of mean annual precipitation (MAP), A1), drought (50 % of MAP, A2), normal (MAP, A3), and high flow year (125 % of MAP, A4). Further, three levels of initial plantation density were considered, including low (50 % of normal initial density, B1), medium (100 %, B2), and high (150 %, B3). The combination of four types of precipitation years and three types of initial population size formed 12 scenarios. To avoid possible unstable states of the final population size caused by the combined effects of extreme drought and high initial plantation density, we conducted an additional scenario using a smaller initial density (1000 individuals ha−1, B4) and extreme drought. Therefore, we had a total of 13 scenarios in this model.
Results
Model validation and sensitivity analysis
ARE ranged from 0 to 0.13 and MARE was 0.048. This indicated that this model was satisfactory for simulating the dynamics of population size (Fig. 2). Mean annual precipitation varied ±10 % which caused variation in outputs of 73.3 % (positive, S = 0.733) to 99.2 % (negative, S = 0.992). Initial plantation density varied ±10 % which caused variation in outputs of 3.57 % (positive, S = 0.0357) to 3.63 % (negative, S = 0.0363).

Calibration results with ARE and MARE
Stability of population size
Population size rapidly changed in the beginning of the simulation. Over time (i.e. 150 year in general), population size fluctuated periodically (Fig. 3). In scenarios with low precipitation (A1 and A2), high initial density (B1, B2) may have driven the population into an unstable status (Fig. 3). Density of Mongolian pine in the different stages followed the trend: stage 5 > stage 1 > stage 4 > stage 2 > stage 3 (Fig. 4). Density of Mongolian pine in stage1 fluctuated periodically, while that in other stages was relative stable (Fig. 4). The descriptive statistics of Figs. 3 and 4 were shown in Tables 2 and 3 respectively.

Simulation results of the dynamics of Mongolian pine populations. Final population size under 125, 100, 50, and 25 % of the mean annual precipitation is shown in (a), (b), (c), and (d), respectively. B1 initial plantation density of 15,000 individuals ha−1, B2 10,000 individuals ha−1, B3 5000 individuals ha−1, B4 1000 individuals ha−1

Simulation results of the dynamics of Mongolian pine age stages. Stages from 1 to 5 indicate young, half-mature, near-mature, mature, and over-mature forest, respectively. a A4B1. b A3B2. c A2B3. d A1B4. Other scenarios are not shown
Effects of precipitation and initial density on population size
The final population size, despite some fluctuation, increased with the increase in precipitation (Fig. 3). The final population size fluctuated in the range between 1400 and 1800 individuals ha−1 under the precipitation level of 125 % of the mean annual (625 mm). Meanwhile, ranges in which the final population size fluctuated under other precipitation scenarios were 1150–1350, 600–700, and 310–340 individuals ha−1 under mean annual, 50 % of the mean annual, and 25 % of the mean annual precipitation, respectively (Fig. 3).
Trends in population dynamics for the three levels of initial plantation density overlapped under 125 and 100 % of mean annual precipitation (Fig. 3). Population size at the low level of initial plantation density (5000 individuals ha−1) persisted under 50 % of mean annual precipitation; however, population size at high and middle levels of initial plantation density (10,000 and 15,000 individuals ha−1) did not persist at that precipitation level. Populations at the three levels of initial plantation density did not persist under 25 % of mean annual precipitation (Fig. 3).
Discussion
Trends in population size for the three levels of initial plantation density overlapped in those scenarios that had high precipitation levels of 125 and 100 % of mean annual, indicating that initial plantation density did not influence the final population size in scenarios with high precipitation. Moreover, in scenarios with low precipitation (A1 and A2), high initial density may have driven the population into an unstable status, suggesting that initial plantation density was important in situations of low water supply in developing Mongolian pine. This was consistent with previous studies, in which initial plantation density of sand-stabilizing vegetation in sandy lands played a vital role for plant survival in conditions of low water supply (Xu and Li 2002). Water supply was the most important resource regulating plant growth in arid and semi-arid lands (Klausmeier 1999). High initial plantation density under low water levels resulted in intra- and inter-specific competition, which might decrease the survival of plants (Xu and Li 2002). Therefore, Mongolian pine with high initial plantation density in scenarios with low precipitation could not reach a stable status.
Population size fluctuated in the range between 1150–1350 individuals ha−1 under mean annual precipitation of 500 mm; this was consistent with the study of Wu et al. (2003), who reported that the population size of Mongolian pine was 1220 individuals ha−1 when it reached a stable status. Population size of 1220 individuals ha−1 is in the range simulated with the model in this study, validating its use. More importantly, the results in this study revealed that the number of individuals of Mongolian pine fluctuated periodically to maintain stability of the population; this is the first report of such fluctuation in Mongolian pine population. A possible explanation is the delayed density dependence, i.e., negative population feedback mechanisms bring the population to carrying capacity with a time lag. In this study, the carrying capacity of Mongolian pine is mainly controlled by precipitation.
As expected, precipitation played a vital role in regulating final population size, and the results showed that population size increased with the increase in precipitation, indicating that higher precipitation levels can support larger Mongolian pine populations. Moreover, the fluctuation in precipitation amounts increased the uncertainty associated with environmental change, but could not enhance the capacity to support the Mongolian pine population.
We applied a system dynamics model to simulate the dynamics of a plant population. A system dynamics approach may also be useful for other biological systems, extending the range of application of this approach. Moreover, in this model, age structures were established, an important but often ignored aspect in the basic research in plant ecology (Chu and Adler 2014). The results of this study, in which combinations of different levels of precipitation and initial plantation density were analyzed with the system dynamics model, may be used in establishing helpful guidelines for plantations of Mongolian pine in dry environments.
Limitations to this approach exist and may be overcome in subsequent research. First, the natural birth and death rates in Mongolian pine populations need a further study. We obtained values for natural birth and death rates in Mongolian pine for this study from the life tables of Picea mongolica populations, as Mongolian pine belonged to the same family. However, uncertainties regarding the applied rates exist and deviations of natural birth and death rates between the two populations may lead to a decrease in the accuracy of modeling results. Second, the death rate should be considered as a function of the population size not as a density-independent constant; this is potentially an essential factor in the death rates of plant populations (Schmickl and Karsai 2010). Third, the allocation of water supply for the Mongolian pine and for herbaceous species present in our ecosystem should be a function of the interaction between the two, as the allocation of water supply between two or more species is known to result from inter-specific competition (Reyer et al. 2010; Xu et al. 2010, 2011).
References
Alamusa, Jiang DM (2008) Characteristics of soil water consumption of two typical shrub and arbor in Horqin Sandy Land. Acta Ecol Sin 28:1981–1990
Chu C, Adler PB (2014) When should plant population models include age structure? J Ecol 102:531–543
Cui QW, Lawson G (1982) A new model of single-species population by extending of the logistic and exponential equations with several limiting conditions. Acta Ecol Sin 2:403–415
Easterling MR, Ellner SP, Dixon PM (2000) Size-specific sensitivity: applying a new structured population model. Ecology 81:694–708
Ellis MM, Crone EE (2013) The role of transient dynamics in stochastic population growth for nine perennial plants. Ecology 94:1681–1686
Ellner SP, Rees M (2006) Integral projection models for species with complex demography. Am Nat 167:410–428
Fan LL, Tang LS, Wu LF, Ma J, Li Y (2014) The limited role of snow water in the growth and development of ephemeral plants in a cold desert. J Veg Sci 25:681–690
Farrer EC, Ashton IW, Knape J, Suding KN (2014) Separating direct and indirect effects of global change: a population dynamics modeling approach using readily available field data. Glob Change Biol 20:1238–1250
Freckleton RP, Sutherland WJ, Watkinson AR, Queenborough SA (2011) Density-structured models for plant population dynamics. Am Nat 177:1–17
Jiao SR (1985) Water balance of Pinus sylvestris var. mongolica afforestion in Zhanggutai Sand Land. Liaoning Forest Sci Tech 12:21–27
Jiao SR (1989) Structure and function of Mongolian pine plantation for sand fixation in Zhanggutai. Liaoning Science and Technology Press, Liaoning
Jiao SR (2001) Report on the causes of the early decline of Pinus sylvestris var. mongolica shelter belt and its preventative and control measures in Zhanggutai of Liaoning Province. Sci Silv Sin 37:131–138
Kang XG (2001) Forest resource management. Chinese Forestry Press, Beijing
Kang HZ, Zhu JJ, Li ZH, Xu ML (2004) Natural distribution of Pinus sylvestris var. mongolica on sandy land and its cultivation as an exotic species. Chin J Ecol 23(5):134–139
Klausmeier CA (1999) Regular and irregular patterns in semiarid vegetation. Science 284:1826–1828
Lendvay B, Kalapos T (2014) Population dynamics of the climate-sensitive endangered perennial Ferula sadleriana Ledeb. (Apiaceae). Plant Spec Biol 29:138–151
Li H, Cai YL (2010) The threshold of eco-security based on system dynamics and application: a case study of Chongming Island. Acta Ecol Sin 30:3654–3664
Li L, Li N, Sheng JD, Wang H (2012) Effects of nitrogen fertilizer and planting density on Alfalfa growth and seed yield. Acta Ecol Sin 20:54–62
Liu JX, Zhang T, Xu XY (2013) Effects of planting density on dynamics growth and biomass of Vetiver. Hubei Agric Sci 52:4126–4129
May RM (1975) Biological populations obeying difference equations-stable points, stable cycles, and chaos. J Theor Biol 51:511–524
McKnight US, Finkel M (2013) A system dynamics model for the screening-level long-term assessment of human health risks at contaminated sites. Environ Model Softw 40:35–50
Qu CP, Guan DX, Wang AZ, Jin CJ, Yuan FH, Wu JB, Ni P (2009) Characteristics of climate change in Horqin Sandy Land in past 56 years. Chin J Ecol 28:2326–2332
Reyer C, Lasch P, Mohren GMJ, Sterck FJ (2010) Inter-specific competition in mixed forests of Douglas-fir (Pseudotsuga menziesii) and common beech (Fagus sylvatica) under climate change—a model-based analysis. Ann For Sci 67:805
Rockwood LL (2006) Introduction to population ecology. Wiley-Blackwell, Hoboken
Rui WY, Li SQ, Lu YS, Zhang JL, Xu YM, Zhou JL, Ren JX, Zhou FM (2011) Growth and competition characteristics of Robinia pseudoacacia forest in the coastal trunk shelterbelt of northern Jiangsu Province. J Northwest For Univ 26:26–30
Schmickl T, Karsai I (2010) The interplay of sex ratio, male success and density-independent mortality affects population dynamics. Ecol Model 221:1089–1097
Smith FE (1963) Population dynamics in Daphnia magna and a new model for population growth. Ecology 44:651
Sun FH, Yang SY, Chen PS (2005) Climatic warming-drying trend in northeastern China during the last 44 years and its effects. Chin J Ecol 24:715–755
Tang Y, Jiang DM, Lv XT (2014) Effects of exclosure management on elm (Ulmus pumila) recruitment in Horqin Sandy Land, northeastern China. Arid Land Res Manag 28:109–117
Wang F, Letort V, Lu Q, Bai X, Guo Y, de Reffye P, Li BG (2012) A functional and structural Mongolian scots pine (Pinus sylvestris var. mongolica) model integrating architecture, biomass and effects of precipitation. PloS One 7(8):e43531. doi:10.1371/journal.pone.0043531
Wei S, Yang H, Song J, Abbaspour KC, Xu Z (2012) System dynamics simulation model for assessing socio-economic impacts of different levels of environmental flow allocation in the Weihe River Basin, China. Eur J Oper Res 221:248–262
Wu CR, Wang JH, Liu SZ, Man DQ, Jin HX, Yan ZZ (2003) Reasonable irrigation of Mongolica pine in sandylands. Prot For Sci Tech 1:59–61
Xu CL, Li ZZ (2002) Population regulation and dynamical simulation of annual plant Eragrostis poaeoides in desert region. Acta Bot Boreal Occident Sin 22:135–140
Xu W, Yang JP, Wang M, Chen WY, Ge CS (2008) Using Vensim to establish nitrogen dynamic simulation model and its validation in paddy field. J Zhejiang Univ 34:649–654
Xu B, Deng X, Zhang S, Shan L (2010) Seedling biomass partition and water use efficiency of switchgrass and milkvetch in monocultures and mixtures in response to various water availabilities. Environ Manag 46:599–609
Xu BC, Xu WZ, Huang J, Shan L, Li FM (2011) Biomass allocation, relative competitive ability and water use efficiency of two dominant species in semiarid Loess Plateau under water stress. Plant Sci 181:644–651
Yi XY, Zhao HL, Cui JH, Li YQ, Zuo XA, Zhuo H (2006) Growth of small area Pinus sylvestris var. mongolica artificial forest under different densities in Horqin Sandy Land, North of China. Acta Ecol Sin 26:1200–1206
Zeng DH, Jiang FQ (1995) The reasonable density of Mongolian pine population in the view of water balance. Prot For Sci Tech 13:4–7
Zeng DH, Pei TF, Fan ZP, Jiang FQ, Zhu JJ (1996) Simulation of canopy interception by Mongolian pine. Chin J Appl Ecol 7:134–138
Zheng YR, Zhang XS, Xu WD (1997) Model forcast of population dynamics of spruce on sandy land. Acta Phytoecol Sin 21:130–137
Zheng X, Zhu JJ, Yan QL, Song LN (2012) Effects of land use changes on the groundwater table and the decline of Pinus sylvestris var. mongolica plantations in southern Horqin Sandy Land, Northeast China. Agric Water Manag 109:94–106
Zhou SR, Wang G (2002) Problems of the size and number of nature reserves. J Gansu Agric Univ 21:193–196
Zhu JJ, Fan ZP, Zeng DH, Qiang FQ, Matsuzaki T (2003) Comparison of stand structure and growth between plantation and natural forests of Pinus sylvestris var. mongolica on sandy land. J For Res 14:103–111
Zhu JJ, Li ZH, Kang HZ, Fan YZ (2005) Effects of polyethylene glycol (PEG)-simulated drought stress on Pinus sylvestris var. mongolica seed germination on sandy land. Chin J Appl Ecol 16:801–804
Zhu JJ, Kang HZ, Tan H, Xu ML (2006) Effects of drought stresses induced by polyethylene glycol on germination of Pinus sylvestris var. mongolica seeds from natural and plantation forests on sandy land. J For Res 11:319–328
Acknowledgments
This work was supported by the National Basic Research Program of China (2013CB429905); and the National Natural Science Foundation of China (41201052). We thank Climatic Data Center, National Meteorological Information Center, China Meteorological Administration for providing climate data. We are grateful to two anonymous reviewers whose comments contributed to improve this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Tang, Y., Li, X. Simulating effects of precipitation and initial planting density on population size of Mongolian pine in the Horqin Sandy Land, China. Agroforest Syst 92, 1–9 (2018). https://doi.org/10.1007/s10457-016-0004-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-016-0004-2