
Abstract
Proline hydroxylation is an important phenomenon of a living cell. Prolyl-4-hydroxylases (P4H) responsible for this process have been characterized from animals, and one of its forms, HIF-P4H, is regarded as an oxygen sensor. In plants, P4H has been partially characterized from few species, and one of the Arabidopsis P4H (AtP4H1) has been shown to hydroxylate proline-rich peptides in vitro. In order to study its function in planta, we have overexpressed AtP4H1 in Arabidopsis. The AtP4H1oexp plants showed hypoxia-in-normoxia phenotype with strict requirement for carbon source for its growth, increased root hair, absence of trichome, and reduction in seed size. Genome-wide expression analyses suggest that expression of several genes related to hypoxia as well as plant growth and development are upregulated in AtP4H1oexp lines. Based on our studies on AtP4H1oexp lines, we speculate a direct role of AtP4H1 in hypoxia stress and in different stages of plant growth and development.







Similar content being viewed by others


References
Baxter-Burrell A, Chang R, Springer P, Bailey-Serres J (2003) Gene and enhancer trap transposable elements reveal oxygen deprivation- regulated genes and their complex pattern of expression in Arabidopsis. Ann Bot 91:129–141
Berra E, Ginouves A, Pouyssegur J (2006) The hypoxia-inducible-factor hydroxylases bring fresh air into hypoxia signaling. EMBO rep 7:41–45
Bolwell GP, Robbins MP, Dixon RA (1985) Elicitor-induced prolyl hydroxylase from French bean (Phaseolus vulgaris). Localization, purification and properties. Biochem J 229:693–699
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Ann Biochem 72:248–254
Bruick RK, McKnight SL (2001) A conserved family of prolyl 4-hydroxylases that modify HIF. Science 294:1337–1340
Cechin AL, Sinigaglia M, Lemke N, Echeverrigaray S, Cabrera OG, Pereira GA, Mombach JC (2008) Cupin: a candidate molecular structure for the Nep1-like protein family. BMC Plant Biol 30:8–50
Clough SJ, Bent AF (1998) Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735–743
Epstein AC, Gleadle JM, Mc Neil LA, Hewitson KS, O’Rourke J, Mole DR, Mukherji M, Metzen E, Wilson MI, Dhanda A, Tian YM, Masson N, Hamilton DL, Jaakkola P, Barstead R, Hodgkin J, Maxwell PH, Pugh CW, Schofield CJ, Ratcliffe PJ (2001) C.elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 107:43–54
Eriksson M, Myllyharju J, Tu H, Hellman M, Kivirikko KI (1999) Evidence for 4-hydroxyproline in viral proteins. Characterization of a viral prolyl 4-hydroxylase and its peptide substrates. J Biol Chem 274:22131–22134
Estevez JM, Kieliszewski MJ, Khitrov N, Somerville C (2006) Characterisation of synthetic hydroxyproline-rich proteoglycans with arabinogalactan protein and extension motifs in Arabidopsis. Plant Physiol 142:458–470
Garcia-Hernandez ER, Lopez GIC (2005) Structural Cell wall proteins from five pollen species and their relationship with boron. Braz J Plant Physiol 17:375–381
Geigenberger P (2003) Response of plant metabolism to too little oxygen. Curr Opin Plant Biol 6:247–256
Hewitson KS, McNeill LA, Riordan MV, Tian Y, Bullock AN, Welford RW, Elkins JM, Oldham NJ, Bhattacharya S, Gleadle JM, Ratcliffe PJ, Pugh CW, Schofield CJ (2002) Hypoxia-inducible Factor (HIF) Asparagine Hydroxylase Is Identical to Factor Inhibiting HIF (FIH) and Is Related to the Cupin Structural Family. J Biol Chem 277:26351–26355
Hieta R, Myllyharju J (2002) Cloning and characterization of a low molecular weight prolyl 4-hydroxylase from Arabidopsis thaliana. Effective hydroxylation of proline-rich, collagen-like, and hypoxia-inducible transcription factor alpha-like peptides. J Biol Chem 277:23965–23971
Hughes BT, Espenshade PJ (2008) Oxygen-regulated degradation of fission yeast SREBP by Ofd1, a prolyl hydroxylase family member. EMBO J 27:1491–1501
Hulskamp M (2004) Plant trichomes: a model for cell differentiation. Nat Rev Mol Cell Biol 5:471–480
Ivan M, Haberberger T, Gervasi DC, Michelson KS, Günzler V, Kondo K, Yang H, Sorokina I, Conaway RC, Conaway JW, Kaelin WG (2002) Biochemical purification and pharmacological inhibition of a mammalian prolyl hydroxylase acting on hypoxia-inducible factor. Proc Natl Acad Sci USA 99:13459–13464
Kaska DD, Myllylä R, Günzler V, Gibor A, Kivirikko KI (1988) Prolyl 4-hydroxylase from Volvox carteri. A low-Mr enzyme antigenically related to the α subunit of the vertebrate enzyme. Biochem J 256:257–263
Kivirikko KI, Myllylä R, Pihlajaniemi T (1992) Hydroxylation of proline and lysine residues in collagens and other animal and plant proteins. In: Harding JJ, Crabbe MJC (eds) Post-translational modifications of proteins. CRC, Boca Raton, pp 1–51
Klok EJ, Wilson IW, Wilson D, Chapman SC, Ewing RM, Somerville SC, Peacock JW, Dolferus R, Dennis ES (2002) Expression Profile Analysis of the low oxygen response in Arabidopsis root cultures. Plant Cell 14:2481–2494
Li C, Wong WH (2001) Model-based analysis of oligonucleotide arrays: Expression index computation and outlier detection. Proc Natl Acad Sci USA 98:31–36
Liu F, VanToai T, Moy LP, Bock G, Linford LD, Quackenbush J (2005) Global transcription profiling reveals comprehensive insights into hypoxic response in Arabidopsis. Plant Physiol 137: 1115–1129
Loreti E, Poggi A, Novi G, Alpi A, Perata P (2005) A Genome-Wide Analysis of the Effects of Sucrose on Gene Expression in Arabidopsis Seedlings under Anoxia. Plant Physiol 137:1130–1138
Myllyharju J (2003) Prolyl 4-hydroxylases, the key enzymes of collagen biosynthesis. Matrix Biol 22:15–24
Qi HH, Ongusaha PP, Myllyharju J, Cheng D, Pakkanen O, Shi Y, Lee SW, Peng J, Shi Y (2008) Prolyl 4-hydroxylation regulates Argonaute 2 stability. Nature 455(7211):421–424 doi:10.1038/nature07186
Ringli C, Bigler L, Kuhn BM, Leiber RM, Diet A, Santelia D, Frey B, Pollmann S, Kleina M (2008) The modified flavonol glycosylation profile in the Arabidopsis rol1 mutants results in alterations in plant growth and cell shape formation. Plant Cell 20(6):1470–1481 doi:10.1105/tpc.107.053249
Schultz CJ, Jhonson KL, Currie G, Bacic A (2000) The classical arabinogalactan protein gene family of Arabidopsis. Plant Cell 12:1751–1768
Tanaka M, Shibata H, Uchida T (1980) A new prolyl hydroxylase acting on poly-L-proline, from suspension cultured cells of Vinca rosea. Biochim Biophys Acta 616:188–198
Thimm O, Bläsing O, Gibon Y, Nagel A, Meyer S, Krüger P, Selbig J, Müller LA, Rhee SY, Stitt M (2004) MAPMAN: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J 37:914–939
Tiainen P, Myllyharju J, Koivunen P (2005) Characterization of a second Arabidopsis thaliana prolyl 4-hydroxylase with distinct substrate specificity. J Biol Chem 280:1142–1148
Vlad F, Spano T, Vlad D, Daher FB, Ouelhadj A, Kalatizis P (2007) Arabidopsis prolyl 4- hydroxylase are differentially expressed in response to hypoxia, anoxia and mechanical wounding. Physiol Plant 130:471–483
Wojtaszek P, Smith CG, Bolwell GP (1999) Ultrastructural localisation and further biochemical characterisation of prolyl 4-hydroxylase from Phaseolus vulgaris: comparative analysis. Int J Biochem Cell Biol 31:463–477
Yuasa K, Toyooka K, Fukuda H, Matsuoka K (2005) Membrane-anchored prolyl hydroxylase with an export signal from the endoplasmic reticulum. Plant J 41:81–94
Zimmermann P, Hirsch-Hoffmann M, Henning L, Gruissem W (2004) GENEVESTIGATOR. Arabidopsis Microarray Database and Analysis Toolbox. Plant Physiol 136:2621–2632
Acknowledgement
Authors are thankful to Director, National Botanical Research Institute, Lucknow for providing necessary facilities for carrying out this work. The authors thankfully acknowledge the financial support provided by Council of Scientific and Industrial Research, Government of India.
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Fig. S1
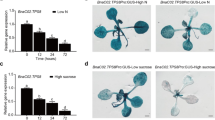
Expression of OsP4H1 in response to hypoxia. Three-week-old rice (Oryza sativa L., Indica cultivar IR64) seedlings were connected to continuous nitrogen supply and samples were collected after 30 min. Numbers above the panel correspond to time of samples collection during nitrogen supply in hours. Expression of ubiquitin genes were used as internal controls for semi-quantitative RT-PCR analysis (PDF 5 kb)
Supplementary Fig. S2
a–c Expression of anaerobic induced genes in AtP4H1oexp lines. Expression of hypoxia responsive genes by qRT-PCR in 2-week-old wild-type and AtP4H1oexp line seedlings (a) Adh alcohol dehydrogenase; Pdc1 pyruvate decarboxylase1; Pdc2 pyruvate decarboxylase2; and Sus1 sucrose synthase. in planta ADH activity staining in wild-type and AtP4H1–3oexp line. Wild-type plants were connected to nitrogen supply for 30 min before staining. Minimum of 20 plants from WT and AtP4H1–3oexp lines were taken for staining and representative pictures are shown in the figure. In gel staining of the ADH activity of the above samples (c). Data are expressed as means ± SD of at least three independent replicates (PDF 50 kb)
Supplementary Fig. S3
The normalization of arrays using three independent methods, i.e., probe logarithmic intensity error (PLIER), robust multiarray average (RMA), and GCRMA using ArrayAssist software (Stratagene, La Jolla, CA, USA). To identify statistically significant differentially expressed genes, a combined criterion of twofold or more change and t test p < 0.05 was adopted. Only common probe sets which were present in three different analyses were considered significant (PDF 32 kb)
Supplementary Fig. S4
Pathways affected by AtP4H1 overexpression as analyzed using MapMan software. Total microarray data was utilized to generate the pathways. Genes for which expression has been affected in AtP4H1 overexpressing lines are shown in pathway. Glycolysis, TCA, and ETC (a) and sugar metabolism (b) pathways are shown in the figure. Of the total genes involved in the pathway up- and downregulated genes are shown by red and blue boxes. White boxes represent for which expression was unaffected in the AtP4H1 overexpressing lines (PDF 71 kb)
Supplementary Fig. S5
Expression profiles of anaerobic genes (a) and trichome-related genes (b) in AtP4H1oexp lines. The color scale (representing one signal log ratio) is shown at the bottom. Differentially expressed genes between our data and as shown by Loreti et al. (2005) were utilized to construct heat map for (a) using one signal log ratio (PDF 45 kb)
Supplementary Fig. S6
In silico analysis of AtP4H1 expression in wild-type plants. The expression profile in different tissue was analyzed using the Genevestigator Gene atlas tool. AtP4H1 expression is high in actively growing tissues, like callus, suspension cells, root elongation zone, and endodermis (PDF 23 kb)
Supplementary Fig. S7
In silico analysis of putative P4H target proteins. PPPPP was used as query to search AGI proteins using BLASTP program. A total of 100 hits were obtained of which GO functional characterization was carried out at TAIR (PDF 3 kb)
Table S1
List of primers used for RT-PCR and qRT-PCR analyses (DOC 34 kb)
Table S2
List of probe sets significantly downregulated twofold in AtP4H1 overexpressing lines (XLS 91 kb)
Table S3
List of probe sets significantly upregulated twofold in AtP4H1 overexpressing lines (XLS 124 kb)
Table S4
Differentially regulated genes associated with various functions (XLS 135 kb)
Table S5
List and fold change of up- and downregulated genes in AtP4H1oexp lines related to hypoxia. Differentially expressed genes between our data and as shown by Loreti et al. (2005) were utilized in the table (XLS 22 kb)
Table S6
List of differentially expressed genes in AtP4H1oexp lines involved in the trichome formation (DOC 26 kb)
Table S7
List of microarray experiments showing the maximum variation in AtP4H1 expression. The list was generated using the gene swinger tool on NASCArrays (http://affymetrix.arabidopsis.info/) (XLS 24 kb)
Table S8
Putative targets of AtP4H1 (XLS 45 kb)
Rights and permissions
About this article
Cite this article
Asif, M.H., Trivedi, P.K., Misra, P. et al. Prolyl-4-hydroxylase (AtP4H1) mediates and mimics low oxygen response in Arabidopsis thaliana . Funct Integr Genomics 9, 525–535 (2009). https://doi.org/10.1007/s10142-009-0118-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10142-009-0118-y