
Abstract
Salinity severely affects plant growth and development. Plants evolved various mechanisms to cope up stress both at molecular and cellular levels. Halophytes have developed better mechanism to alleviate the salt stress than glycophytes, and therefore, it is advantageous to study the role of different genes from halophytes. Salicornia brachiata is an extreme halophyte, which grows luxuriantly in the salty marshes in the coastal areas. Earlier, we have isolated SbASR-1 (abscisic acid stress ripening-1) gene from S. brachiata using cDNA subtractive hybridisation library. ASR-1 genes are abscisic acid (ABA) responsive, whose expression level increases under abiotic stresses, injury, during fruit ripening and in pollen grains. The SbASR-1 transcript showed up-regulation under salt stress conditions. The SbASR-1 protein contains 202 amino acids of 21.01-kDa molecular mass and has 79 amino acid long signatures of ABA/WDS gene family. It has a maximum identity (73 %) with Solanum chilense ASR-1 protein. The SbASR-1 has a large number of disorder-promoting amino acids, which make it an intrinsically disordered protein. The SbASR-1 gene was over-expressed under CaMV 35S promoter in tobacco plant to study its physiological functions under salt stress. T0 transgenic tobacco seeds showed better germination and seedling growth as compared to wild type (Wt) in a salt stress condition. In the leaf tissues of transgenic lines, Na+ and proline contents were significantly lower, as compared to Wt plant, under salt treatment, suggesting that transgenic plants are better adapted to salt stress.





Similar content being viewed by others


References
Agarwal PK, Gupta K, Jha B (2010a) Molecular characterization of the Salicornia brachiata SbMAPKK gene and its expression by abiotic stress. Mol Biol Rep 37:981–986
Agarwal P, Agarwal PK, Joshi AJ, Sopory SK, Reddy MK (2010b) Overexpression of PgDREB2A transcription factor enhances abiotic stress tolerance and activates downstream stress-responsive genes. Mol Biol Rep 37:1125–1135
Amaya I, Botella MA, de la Calle M, Medina MI, Heredia A, Bressan RA, Hasegawa PM, Quesada MA, Valpuesta V (1999) Improved germination under osmotic stress of tobacco plants overexpressing a cell wall peroxidase. FEBS Lett 457:80–84
Amitai-Zeigerson H, Scolnik PA, Bar-Zvi D (1995) Tomato Asr1 mRNA and protein are transiently expressed following salt stress, osmotic stress and treatment with abscisic acid. Plant Sci 110:205–213
Arnon DI (1949) Copper enzymes in isolated chloroplasts polyphenol oxidase in Beta vulgaris. Plant Physiol 24:1–15
Ashraf M, Foolad MR (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot 59:206–216
Ashraf M, Harris PJ (2004) Potential biochemical indicators of salinity tolerance in plants. Plant Sci 166:3–16
Bates LS, Waldern R, Teare ID (1973) Rapid determination of free proline for water stress studies. Plant Soil 39:205–207
Bubner B, Gase K, Baldwin IT (2004) Two-fold differences are the detection limit for determining transgene copy numbers in plants by real-time PCR. BMC Biotechnol 4:14–24
Cakir B, Agasse A, Gaillard C, Saumonneau A, Delrot S, Atanassova R (2003) A grape ASR protein involved in sugar and abscisic acid signaling. Plant Cell 15:2165–2180
Carrari F, Fernie AR, Iusem ND (2004) Heard it through the grapevine? ABA and sugar cross-talk: the ASR story. Trends Plant Sci 9:57–59
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
Dai J-R, Liu B, Feng D-R, H-y L, Y-m H, K-b Q, Wang H-B, Wang J-F (2011) MpAsr encodes an intrinsically unstructured protein and enhances osmotic tolerance in transgenic Arabidopsis. Plant Cell Rep 30:1219–1230
Das M, Chauhan H, Chhibbar A, Mohd Q, Haq R, Khurana P (2011) High-efficiency transformation and selective tolerance against biotic and abiotic stress in mulberry, Morus indica cv. K2, by constitutive and inducible expression of tobacco osmotin. Transgenic Res 20:231–246
de Jonge HR, Hogema B, Tilly BC (2000) Protein N-myristoylation: critical role in apoptosis and salt tolerance. Sci. STKE 2000:pe1-4
Farazi TA, Waksman G, Gordon JI (2001) The biology and enzymology of protein N-myristoylation. J Biol Chem 276:39501–39504
Fink AL (2005) Natively unfolded proteins. Curr Opin Struct Biol 15:35–41
Frankel N, Nunes-Nesi A, Balbo I, Mazuch J, Centeno D, Iusem ND, Fernie AR, Carrari F (2007) ci21A/Asr1 expression influences glucose accumulation in potato tubers. Plant Mol Biol 63:719–730
Giri B, Kapoor R, Mukherji KG (2007) Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza, Glomus fasciculatum, may be partly related to elevated K+/Na+ ratios in root and shoot tissues. Microb Ecol 54:753–760
Goldgur Y, Rom S, Ghirlando R, Shkolnik D, Shadrin N, Konrad Z, Bar-Zvi D (2007) Desiccation and zinc binding induce transition of tomato abscisic acid stress ripening 1, a water stress- and salt stress-regulated plant-specific protein, from unfolded to folded state. Plant Physiol 143:617–628
Guo L, Yu Y, Xia X, Yin W (2010) Identification and functional characterisation of the promoter of the calcium sensor gene CBL1 from the xerophyte Ammopiptanthus mongolicus. BMC Plant Biol 10:18–33
Gupta K, Agarwal PK, Reddy MK, Jha B (2010) SbDREB2A, an A-2 type DREB transcription factor from extreme halophyte Salicornia brachiata confers abiotic stress tolerance in Escherichia coli. Plant Cell Rep 29:1131–1137
Horsch RB, Fry JE, Hoffmann NL, Eichholtz D, Rogers SG, Farley RT (1985) A simple and general method for transferring genes into plants. Science 227:1229–1231
Husaini AM, Abdin MZ (2008) Development of transgenic strawberry (Fragaria × ananassa Duch.) plants tolerant to salt stress. Plant Sci 174:446–455
Ishitani M, Liu J, Halfter U, Kim C-S, Shi W, Zhu J-K (2000) SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding. Plant Cell 12:1667–1677
Iusem ND, Bartholomew DM, Hitz WD, Scolnik PA (1993) Tomato (Lycopersicon esculentum) transcript induced by water deficit and ripening. Plant Physiol 102:1353–1354
Jha A, Joshi M, Yadav NS, Agarwal PK, Jha B (2011a) Cloning and characterization of the Salicornia brachiata Na+/H+ antiporter gene SbNHX1 and its expression by abiotic stress. Mol Biol Rep 38:1965–1973
Jha B, Agarwal PK, Reddy PS, Lal S, Sopory SK, Reddy MK (2009) Identification of salt-induced genes from Salicornia brachiata, an extreme halophyte through expressed sequence tags analysis. Genes Genet Syst 84:111–120
Jha B, Sharma A, Mishra A (2011b) Expression of SbGSTU (tau class glutathione S-transferase) gene isolated from Salicornia brachiata in tobacco for salt tolerance. Mol Biol Rep 38:4823–4832
Kalifa Y, Perlson E, Gilad A, Konrad Z, Scolnik PA, Bar-Zvi D (2004) Over-expression of the water and salt stress-regulated Asr1 gene confers an increased salt tolerance. Plant Cell Environ 27:1459–1468
Konrad Z, Bar-Zvi D (2008) Synergism between the chaperone-like activity of the stress regulated ASR1 protein and the osmolyte glycine-betaine. Planta 227:1213–1219
Koiwa H, Li F, McCully MG, Mendoza I, Koizumi N, Manabe Y, Nakagawa Y, Zhu J, Rus A, Pardo JM, Bressan RA, Hasegawa PM (2003) The STT3a subunit isoform of the Arabidopsis oligosaccharyltransferase controls adaptive responses to salt/osmotic stress. Plant Cell 15:2273–2284
Lal S, Gulyani V, Khurana P (2008) Overexpression of HVA1 gene from barley generates tolerance to salinity and water stress in transgenic mulberry (Morus indica). Transgenic Res 17:651–663
Leigh RA, Ahmad N, Wyn Jones RG (1981) Assessment of glycine betaine and proline compartmentation by analysis isolated beet vacuoles. Planta 153:34–41
Levitt J (1980) Responses of plants to environmental stress: water, radiation, salt and other stresses. Academic Press, New York
Liu H-Y, Dai J-R, Feng D-R, Liu B, Wang H-B, Wang J-F (2010) Characterization of a novel plantain Asr gene, MpAsr, that is regulated in response to infection of Fusarium oxysporum f. sp. cubense and abiotic stresses. J Integr Plant Biol 52:315–323
Lutts S, Kinet JM, Bouharmont J (1996) NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann Bot 78:389–398
Mahajan S, Tuteja N (2005) Cold, salinity and drought stresses: an overview. Arch Biochem Biophys 444:139–158
Maskin L, Frankel N, Gudesblat G, Demergasso MJ, Pietrasanta LI, Iusem ND (2007) Dimerization and DNA-binding of ASR1, a small hydrophilic protein abundant in plant tissues suffering from water loss. Biochem Biophys Res Commun 352:831–835
Maskin L, Gudesblat GE, Moreno JE, Carrari FO, Frankel N, Sambade A, Rossi MM, Iusem ND (2001) Differential expression of the members of Asr gene family in tomato (Lycopersicon esculentum). Plant Sci 161:739–746
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Parkhi V, Kumar V, Sunilkumar G, Campbell LM, Singh NK, Rathore KS (2009) Expression of apoplastically secreted tobacco osmotin in cotton confers drought tolerance. Mol Breed 23:625–639
Prilusky J, Feder CE, Zeev-Ben-Mordehai T, Rydberg E, Man O, Beckman JS, Silman I, Sussman JL (2005) FoldIndex: a simple tool to predict whether a given protein sequence is intrinsically unfolded. Bioinformatics 21:3435–3438
Rom S, Gilad A, Kalifa Y, Konrad Z, Karpasas MM, Goldgur Y, Bar-Zvi D (2006) Mapping the DNA- and zinc-binding domains of ASR1 (abscisic acid stress ripening), an abiotic-stress regulated plant specific protein. Biochimie 88:621–628
Sairam RK (1994) Effects of homobrassinolide application on plant metabolism and grain yield under irrigated and moisture-stress conditions of two wheat varieties. Plant Growth Regul 14:173–181
Shepherd CT, Lauter ANM, Scott MP (2009) Determination of transgene copy number by real-time quantitative PCR. In: Scott MP (ed) Methods in molecular biology: transgenic maize. Humana, New York
Shukla PS, Agarwal PK, Jha B (2011) Improved salinity tolerance of Arachis hypogaea (L.) by the interaction of halotolerant plant-growth-promoting rhizobacteria. J Plant Growth Regul. doi:10.1007/s00344-011-9231-y
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739
Töpfer R, Matzeit V, Gronenborn B, Schell J, Steinbiss HH (1987) A set of plant expression vectors for transcriptional and translational fusions. Nucleic Acids Res 15:5890
Uversky VN, Dunker AK (2010) Understanding protein non-folding. Biochim Biophys Acta 1804:1231–1264
Uversky VN, Gillespie JR, Fink AL (2000) Why are “natively unfolded” proteins unstructured under physiologic conditions? Proteins Struct Funct Genet 41:415–427
Virlouvet L, Jacquemot M-P, Gerentes D, Corti H, Bouton S, Gilard F, Valot B, Trouverie J, Tcherkez G, Falque M, Damerval C, Rogowsky P, Perez P, Noctor G, Zivy M, Coursol S (2011) The ZmASR1 protein influences branched-chain amino acid biosynthesis and maintains kernel yield in maize under water-limited conditions. Plant Physiol 157:917–936
Wang HJ, Hsu CM, Jauh GY, Wang CS (2005) A lily pollen ASR protein localizes to both cytoplasm and nuclei requiring a nuclear localization signal. Physiol Plant 23:314–320
Wang C, Jing C, Mao X, Chang X, Li A (2011) TaABC1, a member of the activity of bc1 complex protein kinase family from common wheat, confers enhanced tolerance to abiotic stresses in Arabidopsis. J Exp Bot 62:1299–1311
Yang CY, Chen YC, Jauh GY, Wang CS (2005) A lily ASR protein involves abscisic acid signaling and confers drought and salt resistance in Arabidopsis. Plant Physiol 139:836–846
Zhang HX, Blumwald E (2001) Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat Biotechnol 19:765–768
Zhu JK (2002) Salt and drought stress signal transduction in plants. Annu Rev Plant Biol 53:247–273
Acknowledgments
The authors are thankful to the Council of Scientific and Industrial Research, New Delhi (NWP-020), for the financial assistance. VT is thankful to CSIR for the Junior Research Fellowship.
Author information
Authors and Affiliations
Corresponding authors
Electronic Supplementary Materials
Below is the link to the electronic supplementary material.
Supplementary Fig. 1
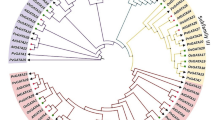
The phylogenetic tress was prepared using protein accession number, viz. AAB96681 (Oryza sativa), ACI15208 (Salicornia brachiata), ACZ60128 (Musa acuminate), AAA82741 (Citrus maxima), BAI94530 (Prunus mume), AAL26889 (Prunus persica), AAB97140 (Prunus armeniaca), AAP46155 (Hevea brasiliensis), AAA34137 (Solanum lycopersicum), AAY97998 (Solanum lycopersicum), AAZ93634 (Vitis vinifera), AAA21866 (Zea mays), AAB64185 (Solanum lycopersicum), AAM51877 (Lilium longiflorum), AAD00254 (Solanum tuberosum), AAL27560 (Cucumis melo), XP_002524296 (Ricinus communis), AAY97997 (Solanum cheesmaniae), AAY98001 (S. chilense), AAY98002 (Solanum corneliomuelleri), AAY98000 (Lycopersicon peruvianum), AAY97999 (Solanum habrochaites), AAR23420 (Ginkgo biloba), ABC86744 (Vitis pseudoreticulata), ACL68147 (Musa acuminata), AAB02692 (Pinus taeda) and BT114797 (Picea glauca). AAB96796 (Solanum lycopersicum ER5) was used as an out-group to make the root of the tree. The evolutionary history of ASR-1 amino acid sequences was inferred by using the maximum likelihood method based on the JTT matrix-based model using MEGA version 5 software. The bootstrap consensus tree inferred from 1,000 replicates is taken to represent the evolutionary history of the taxa analysed. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 28 amino acid sequences. All ambiguous positions were removed for each sequence pair. There were a total of 222 positions in the final data set (JPEG 70 kb)
Supplementary Fig. 2
Multiple alignment of ASR-1 amino acid sequences was carried out using the sequences as in Supplementary Fig. 1 by online ClustalW program. Conserved WDS/ABA domain annotated by query against InterPro is underlined. Other conserved domains are shaded in different colours and shown by double-sided solid arrows. A N-terminal His-rich domain is highlighted with yellow colour but absent in SbASR-1 and underlined. B Zn2+-dependent DNA-binding domain of tomato ASR-1 (Rom et al. 2006). C Hydrophobic tomato ASR-1 sequence hindering DNA binding (Rom et al. 2006). D and E C-terminal-conserved NLS sequence found in two K-rich amino acid clusters separated by non-conserved 11 amino acid residues in lily LLA23 (Wang et al. 2005) while by 16 amino acid residues in SbASR-1 (DOC 43 kb)
Supplementary Fig. 3
Secondary structure of SbASR-1 predicted by ExPasy tools shows five coils and four helix loops (JPEG 111 kb)
Supplementary Table 1
Comparative analysis of SbASR-1 and other glycophytic ASR-1 proteins showed higher percentage of disorder-promoting amino acid residues and N-myristoylation site (DOC 44.0 kb)
Supplementary Table 2
Prediction of N-glycosylation sites, protein kinase C phosphorylation sites, tyrosine kinase phosphorylation sites and N-myristoylation sites by PROSCAN.BASE server (JPEG 67 kb)
Rights and permissions
About this article
Cite this article
Jha, B., Lal, S., Tiwari, V. et al. The SbASR-1 Gene Cloned from an Extreme Halophyte Salicornia brachiata Enhances Salt Tolerance in Transgenic Tobacco. Mar Biotechnol 14, 782–792 (2012). https://doi.org/10.1007/s10126-012-9442-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10126-012-9442-7