
Abstract
The rabbit kidney cell line RK13 has been reported to be contaminated with noncytopathogenic (ncp) bovine viral diarrhea virus (BVDV). Persistent infection was confirmed by demonstrating the stability of virus titers (104.6±0.5 TCID50/ml) and BVDV positive cells (71.9 ± 3.12 %), over six successive passages. Based on the “exaltation of Newcastle disease virus” (END) and reverse plaque formation methods, two types of ncp viruses were isolated, END-phenomenon-positive and negative. Isolates, RK13/E+ and RK13/E−, demonstrated (1) differing levels of reproducibility in cell cultures, (2) similar antigenicity against BVDV antisera, (3) identical 5′-UTR region nucleotide sequences, (4) four amino acid differences throughout the genomic open reading frame, and (5) better growth ability in primary rabbit cells than other laboratory strains when inoculated in parallel at an MOI of 0.01. Overall, the BVDV population in RK13 cells consists of at least two different END characteristic quasispecies that are adapted to cultures of rabbit origin, giving rise to naturally attenuated BVDV strains that can be used in vaccine development.
Similar content being viewed by others


Introduction
Bovine viral diarrhea virus (BVDV) is an infectious agent of cattle, causing diarrhea and mucosal disease, resulting in economic losses to the livestock industry worldwide. Along with classical swine fever virus (CSFV) and border disease virus (BDV), BVDV belongs to the genus Pestivirus within the family Flaviviridae. All members of this family are enveloped viruses, with a single-stranded positive-sense RNA genome, encompassing one large open reading frame (ORF), flanked by 5′ and 3′ untranslated regions (UTRs) [34, 50].
Under in vitro culturing conditions, BVDV exhibits two biotypes that can be distinguished according to their lytic effect in permissive cell cultures: cytopathogenic (cp) and noncytopathogenic (ncp) [24, 50]. More than 90 % of BVDV field strains are ncp BVDVs, which can multiply in cell culture without inducing cytopathogenic effect (CPE) and therefore go undetected [24]. This feature facilitates adventitious virus contamination, posing serious risks to virology diagnostics, research, and the production of vaccines [7]. Several types of cell culture have been reported to be contaminated with BVDV through fetal bovine serum (FBS) [32, 37], which is commonly used for promoting cell growth [1, 32]. One such culture is the RK13 rabbit kidney cell line from the American Type Culture Collection (ATCC) [3, 18, 43].
Under in vitro conditions, BVDV’s ability to enter and replicate is restricted to cells of host origin [40, 41]. However, its pattern of replication varies, within various pestivirus host cell types [44]. For example, BVDV can replicate in cultures of cells originating from natural hosts, such as ruminants and swine, whereas it rarely occurs in non-natural hosts, such as rabbits and cats [3, 44]. Therefore, adaptation of BVDV strains to non-natural host cells can provide an opportunity for studying aspects of virus-host cell interactions that are not readily observed in cell cultures of bovine origin [12]. The ability of this virus to multiply in cells of heterologous origin may be indicative of a capacity for similar adaptation in vivo [33]. The exact biological and genetic characteristics of BVDV that persistently infects the RK13 cell line are unclear.
For identification of the biological features of BVDV, all experimental procedures were first established for CSFV and then applied to BVDV. It has been reported that marked CPE was induced by superinfection with the Miyadera strain of Newcastle disease virus (NDV) in primary swine testicle (ST) cells that had been previously infected with CSFV, while no CPE was induced in normal ST cells. This was referred to as the “exaltation of Newcastle disease virus” (END) phenomenon [27, 28]. In addition, an END method based on this phenomenon was developed for the titration of CSFV [27]. Subsequently, a virus that did not show the END phenomenon but showed interference against NDV and western equine encephalitis virus (WEEV) was found in a CSFV population [49]. Interference and reverse plaque formation (RPF) methods with vesicular stomatitis virus (VSV) based on this interference phenomenon were developed for the detection of CSFV that does not show the END phenomenon [13, 22, 49]. Inaba et al. [19] determined that BVDV shows the END phenomenon in primary bovine testicle (BT) cells in a manner similar to that of CSFV.
With respect to genetic characteristics, RNA viruses, including BVDV, lack proofreading activity [4, 25, 42], resulting in very high error rates during replication, leading to highly diverse viral populations termed quasispecies, as originally proposed by Eigen [9–11]. To date, no method for the detection and quantification of variants in a quasispecies has been developed, except for gene cloning and sequencing. Therefore, there is a need for other detection and titration approaches for small populations of variants in order to detect live variants in a quasispecies. The RPF method has been available for the detection and quantification of nearly identical live viral variants in major heterologous populations of BVDV. Itoh et al. [21] and Nakamura et al. [35] isolated a minor population of BVDV that does not show the END phenomenon but shows interference against VSV in the ncpBVDV laboratory strain, which constitutes a major population, by using the RPF method. Since then, ncpBVDV has been subdivided into two groups: END-positive (END+) virus, which shows the END phenomenon, and END-negative (END−) virus, which shows heterologous interference phenomena [35, 36].
Several BVDV adaptation studies, conducted in rabbits under in vitro and/or in vivo conditions with the purpose of producing an attenuated live vaccine have been reported previously [2, 6, 12]. Therefore, the main aim of this study was to isolate and characterize BVDV biotypes found in the RK13 cell line with different END characteristics, focusing on the following parameters: virus persistence, adaptation, in vitro amplification, infectivity, and genetic affinity, which may have implications for potential vaccine development. Identification of a naturally attenuated BVDV strain, whose pathogenicity was reduced through natural adaptation to cells of rabbit origin, would bypass the normal arduous and time-consuming manual attenuation procedure that is often required for virus vaccine development.
Materials and methods
Cells
The RK13 cell line, originating from rabbit kidney, was obtained from ATCC, USA. This cell line was at passage 89 when purchased from a distributor in Japan. The MDBK-SY cell line, cloned from the MDBK cell line, was provided by the Tochigi Prefectural Central Livestock Hygiene Service Center (Tochigi, Japan). These cells were grown as monolayers in Eagle’s minimum essential medium (MEM) (Nissui, Japan) supplemented with 2 mM glutamine, 0.295 % tryptose phosphate broth (TPB), 5 % inactivated fetal bovine serum (FBS) free of pestiviruses and BVDV antibodies, 1.125 mg/mL NaHCO3, and antibiotics (100 IU/mL penicillin and 100 μg/mL streptomycin). Primary bovine testicle (BT) and rabbit kidney (RK) cell cultures were prepared in MEM containing 10 % inactivated FBS, 1.125 mg/mL NaHCO3, 2 mM glutamine, and antibiotics.
Viruses
The virus that was isolated from the RK13 cell line was named the parent RK13 strain and used for the isolation of BVDV showing negativity for the END phenomenon, and also for the cloning of BVDV showing positivity for the END phenomenon. BVD viruses isolated from the parent RK13 strain in this experiment were named the RK13/E− strain and the RK13/E+ strain, respectively, and were used for the analysis of biological and genetic characteristics. NcpBVDVs (No.12/E+, No.12/E−, Nose/E+, Nose/E− and KZ91/E+) and cpBVDVs (Nose and KZ91CP strains) were used as controls for comparison. The No.12/E+, Nose/E+, and KZ91/E+ strains of ncpBVDV show the END phenomenon, while the RK13/E−, Nose/E−, and No.12/E− strains of ncpBVDV do not show the END phenomenon but do show the interference phenomenon. All BVDV strains were propagated for stock virus, using MDBK-SY cells. The Miyadera strain of NDV and the New Jersey serotype of VSV were used as the challenge viruses in the END and interference methods, respectively. NDV was propagated in the allantoic cavities of 9-day-old embryonated hens’ eggs, with allantoic fluid being collected and stored as stock virus. VSV was propagated using a Vero cell line.
END method
The END method was used for the titration and neutralization test of END-phenomenon-positive viruses, parent RK13 and No12/E+ strains, according to the original method [19, 27], with the exception of the cultured cells used. This method is applicable only for detection and titration of END-phenomenon-positive viruses. Briefly, after each tenfold dilution of BVDV, 0.05 mL of dilution was placed in 5 wells of a 96-well microplate, followed by addition of 0.15 mL of MDBK-SY cell suspensions (~2 × 103 cells) in each well. The plate was then incubated at 37 °C in a 5 % CO2 humid incubator for 5 days before aspiration of culture fluid from each well. Then, 0.1 mL of NDV suspension (107 TCID50/mL) was added to each well, and the plate was incubated again for 3 days at 37 °C in a 5 % CO2 humid incubator. Cells were examined for CPE under a light microscope and were considered BVDV positive when they showed a clear CPE. The virus titre was estimated based on the highest dilution of BVDV/E+ yielding CPE by NDV, which will be referred throughout the manuscript hereafter as the “limiting dilution”. The 50 % tissue culture infectious dose (TCID50) was calculated according to the method described by Kärber [30].
Interference method with VSV
The interference method with VSV was carried out according to Fukousho et al. [14] with the modification that MDBK-SY cells were used. Briefly, after each tenfold dilution of BVDV, 0.05 mL of dilution was placed in 5 wells of a 96-well microplate, followed by addition of 0.15 mL of MDBK-SY cell suspension to each well. After incubation for 5 days, 0.1 mL of VSV suspension (107 TCID50/mL) was added to each well, and cells were examined for inhibition of CPE after 2 days and were considered BVDV-positive when showing no CPE. The TCID50 was calculated according to the method described by Kärber [30]. This method was used for the titration and neutralization test of END-phenomenon-negative viruses, and also for the cloning of the RK13/E− strain.
Reverse plaque formation (RPF) method
The RPF method was carried out according to the original method [13] with some modifications. The stock virus of parent RK13 strain was serially diluted tenfold with MEM, and applied to 60-cm2 Petri dishes containing fully grown monolayers of MDBK-SY cells. Dishes were incubated for 90 min at 37 °C in a humid 5 % CO2 incubator for virus absorption. After 90 min, dishes were gently washed twice with MEM and covered with 5 mL of methylcellulose overlay medium (MEM with 5 % FBS and 1.5 % methylcellulose). After 5 days of incubation, the overlay medium was removed and washed out using warm MEM (39-41 °C). The infected cell monolayer was challenged with VSV (107 TCID50/mL) and incubated for 1 hour at 37 °C for absorption. The monolayer was washed twice with MEM and covered again with methylcellulose overlay medium with 3.5 % NaHCO3. Two days after incubation, the overlay medium was removed and washed out using warm MEM. The cells that formed reverse plaques were collected using a penicillin cup with 0.1 % trypsin for trypsinization. The collected sample was then suspended with MEM and centrifuged for 5 min at 1,000 rpm. The sedimented cells were resuspended in MEM containing antiserum against VSV and BVDV and incubated at 37 °C for 90 min. After neutralization with antisera, the cells were washed three times with MEM and frozen at −80 °C to release the intracellular viruses. The freeze-thaw process was repeated three times, and the collected suspensions were then centrifuged for 10 min at 2000 rpm. Supernatants were collected and serially diluted tenfold with MEM. The interference method with VSV [14] was carried out for virus cloning and confirmation of the existence of virus showing intrinsic interference.
Immunoperoxidase assay
A microplate immunoperoxidase test was carried out according to the instructions in the OIE manual [8]. Positive reactions of viral antigens with anti-BVDV NS3 monoclonal antibody (mAb) (JCU TropBio, Australia) were shown by the development of a red-brown color. Over six successive passages of RK13 cells, the proportion of positively infected cells was calculated by dividing the number of positive cells by the total cell number, as observed in ten independent foci (n.10).
Serum neutralization (SN) test
A serum cross-neutralization test was performed between RK13/E+, RK13/E−, Nose, and KZ91CP viruses and rabbit RK13/E+, Nose and KZ91 antisera. Briefly, serial twofold dilutions of inactivated antisera of RK13/E+, Nose, and KZ91NCP were made in 96-well microplates using MEM. Each dilution was mixed with an equal volume of 200 TCID50/0.1 mL of each virus: RK13/E+, RK13/E−, Nose, and KZ91CP. After 1 hour of incubation at 37 °C, 0.1 mL of MDBK-SY cell suspension was mixed with 0.1 mL of each dilution mixture per well. Microplates were then incubated for up to 5 days at 37 °C in a 5 % CO2 incubator. Fluids of RK13/E+ and RK13/E− infected cultures were removed from each well and subsequently challenged with NDV and VSV, respectively. The SN titer against ncp RK13 viruses was determined as the reciprocal of the highest serum dilution capable of inhibiting CPE by NDV, and exhibiting CPE by VSV. The SN titer against Nose and KZ91CP was determined as the reciprocal of the highest serum dilution capable of inhibiting CPE.
RNA extraction, RT-PCR, and sequencing
Total RNA was extracted from parent RK13, RK13/E+, and RK13/E− strains propagated in MDBK-SY cells, using a QIAamp MinElute Virus Spin Kit (QIAGEN, Germany). RT-PCR was carried out using an RNA PCR Kit (AMV) Version 3.0 (TaKaRa). The 5′-UTR of each BVDV RNA was amplified and sequenced using primers 324 and 326 [46, 51]. After amplification, the PCR products were resolved on a Tris-borate-EDTA (TBE)-2 % agarose gel by electrophoresis in TBE buffer. The PCR products were extracted from the gel using a QIAquick Gel Extraction Kit (QIAGEN, Germany). Nucleotide sequences were determined by direct sequencing of PCR products using a BigDye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems, USA) and an ABI PRISM 310 Genetic Analyzer according to the manufacturer’s instructions. All of the sequences were confirmed by dual-strand sequencing. The nucleotide sequences of 5′-UTR were proofread using GENETYX software (Genetyx Corp., Japan). Phylogenetic analysis was performed using the sequence spanning nucleotide positions 135 to 354 in the BVDV SD-1 genome [5], employing the neighbor-joining method as reported previously [52]. The elucidated sequences of RK13 cell line-origin viruses were compared to the reported type strains in the GenBank database and used for phylogenetic analysis.
Comparison of virus growth in cells of bovine and rabbit origin
Confluent monolayers of MDBK-SY and primary RK cells were inoculated in parallel with RK13/E+, RK13/E−, No.12/E+, and No.12/E− strains at a multiplicity of infection (MOI) of 0.01. After 90 min of absorption, cells were washed twice with medium and maintained in fresh medium at 37 °C in a humid incubator with 5 % CO2 Supernatants and cells were harvested at 24-hour intervals post-inoculation (pi) for up to 6 days. The virus titer was estimated by the END and interference methods for all supernatant and intracellular phases as described above.
Rabbit infection
Two 2-month-old Japanese white rabbits (1.84 to 1.89 kg) were housed in individual cages in an animal isolation room. Both animals were confirmed to be seronegative for BVDV before initial injection. One rabbit was injected intravenously with 1 mL of RK13/E+ virus (106.4TCID50/mL), whereas the other received 1 mL of RK13/E− virus (105.8TCID50/mL). Body temperature and clinical signs were observed daily. On day 6 post-injection, blood samples were collected and mixed with EDTA for virus isolation from white blood cells (WBC). After collecting blood samples, both rabbits were necropsied, and their visceral organs (spleen, mesenteric lymph nodes [MLN], kidney, liver, and pancreas) were collected for virus isolation. The rabbit infection experiment was performed in accordance with guidelines of the Nippon Veterinary and Life Science University Animal Care and Use Committee.
Results
Identification of a virus persistently infecting the RK13 cell line
When the RK13 cell line was obtained from an ATCC distributor in Japan, virus isolation was carried out directly, without cell passage, using RK13 cell line culture fluid. When the culture supernatant was applied to MDBK-SY cells, no CPE was observed for 7 days. The virus was detected by the END method and identified as BVDV by immune staining against BVDV-NS3 protein. Overall, the isolated virus was identified as having the characteristics of BVDV and of being END-phenomenon-positive, and was named the parent RK13 strain of BVDV.
Kinetics of BVDV replication in the persistently infected RK13 cell line
When the RK13 cell line was passaged 31 times (from the 89th to 120th passages) in 25-cm2 culture flasks, 0.5 mL of culture fluid was collected at 5 days after cell seeding at each passage and stored at −80 °C. After harvesting the culture fluid, cells were trypsinized and subsequently passaged to the next generation, at a dilution of 1:3. BVDV was detected in culture fluid at each passage by the END method. Virus was retained in culture fluid at a titer of 104.5±0.7 TCID50/mL over 31 subsequent passages (passages 89 to 120) of the RK13 cell line.
In addition, six consecutive passages (passages 89 to 94) were used to confirm the infection with BVDV by immune staining with an anti-NS3 monoclonal antibody (mAb). Virus was detected in the extracellular and intracellular fractions from the RK13 cell line at a titer of 104.6±0.5 TCID50/mL at each passage (Fig. 1). To determine the population of BVDV-positive cells during passage of the RK13 cell line, the proportion of positively infected cells was determined by immune staining with anti-NS3 mAb over six subsequent passages (89th to 94th passages). The average proportion of BVDV-positive cells was 71.9 ± 3.12 % through all six passages, as shown in Fig. 1.

Estimated titers of contaminant BVDV in supernatant and cellular fractions of the RK13 cell line and proportion of BVDV-positive cells. The x-axis indicates the passage number, while the left y-axis indicates virus titer, expressed as log10 TCID50/mL. The solid circle (●) and open circle (○) represent the viral titer in the supernatant and cellular fraction, respectively. The solid square (■) represents the proportion of NS3-positive cells expressed as a percentage, as determined by immunostaining. The right y-axis indicates the proportion of BVDV-positive cells, expressed as a percentage
Isolation of END− virus from the parent RK13 strain using the RPF method
As mentioned above, the parent RK13 strain isolated from the RK13 cell line showed the END phenomenon and could be titrated by the END method, but not by the interference method. The RPF method was used to examine whether a small number of viruses showing the interference phenomenon were mixed together in the virus stock of the parent RK13 strain showing the END phenomenon. Consequently, BVDV that does not show the END phenomenon was isolated from the parent RK13 strain virus stock. This virus showed the interference phenomenon against VSV, but no CPE or END phenomenon in MDBK-SY cells and primary BT cells. After four repeats of cloning by limiting dilution using the interference method with VSV, the stock virus was propagated in MDBK-SY cells, reaching a titer of up to 106 TCID50/mL. This virus was designated as the RK13/E− isolate of BVDV.
The major population of virus in the parent RK13 strain was cloned four times by the limiting dilution technique using the END method with MDBK-SY cells. The cloned virus showed the END phenomenon, but no CPE or any interference phenomenon with VSV. The virus titer reached up to 108TCID50/mL in MDBK-SY cells. This virus was named the RK13/E+ isolate of BVDV.
Biological and genetic characteristics of RK13/E+ and RK13/E− strains
Both RK13/E− and RK13/E+ strains showed the characteristics of BVDV, such as the END phenomenon (negative or positive) and the interference phenomenon (positive or negative) and were also identified as BVDV by immune staining using mAb against the BVDV NS3 protein. The results of the serum neutralization tests are shown in Table 1. Identical antigenicity profiles were observed with the RK13/E+ and RK13/E− strains; SN titers were 1,024 with homologous serum. However, RK13 cell line-origin viruses demonstrated a significant difference in antigenicity compared with other BVDV isolates, with a neutralizing Nose (BVDV-1) antiserum titer of 256, compared with a KZ91CP antiserum titer (BVDV-2) of <8.
The nucleotide sequences of the 5′-UTRs of parent RK13, RK13/E+ (accession number: JX419398), and RK13/E− (accession number: JX419397) strains were determined and aligned. Within a 250-base region of the 5′-UTR, the homology between all strains was 100 %. A preliminary phylogenetic tree was created using the nucleotide sequence of the RK13 cell line-origin strain and 16 BVDV previously reported strains from the GenBank database (Fig. 2). As a result, RK13 cell line-origin viruses were classified as BVDV genotype 1b, along with BVDV CP7 (accession number; AF220247) with high homology (96.7 %).

Phylogenetic analysis of the 5′-UTR of RK13 strains: Sequences of 12 strains of BVDV, 3 strains of CSFV, and 1 strain of BDV. The phylogenetic tree was constructed by the neighbor-joining method and bootstrap test (n = 500)
To analyze the genetic characteristics of RK13/E+ and RK13/E− strains, nucleotide sequences of the ORF were determined, in addition to partial sequences of the 5′- and 3′-UTRs. The lengths of the sequences were similar between the strains, with a length of 12,061 bp at positions 130-12,190 in the BVDV SD-1 strain (accession number M96751). There was a single large ORF spanning 11,694 nucleotides, which could encode a large protein of 3,898 amino acids. There was no subgenomic RNA, nor was there an insertion of a cellular RNA sequence or duplication of a viral sequence in the genome, which are characteristic of the genomes of cytopathogenic BVDV strains. There were differences at 12 nucleotide positions within the ORF between the RK13/E+ and RK13/E− strains, only four of which resulted in amino acid differences: His130Pro in Npro, Val760Ala in E2, Val2086Ala in NS3, and Lys3757Gln in NS5B (Table 2).
Comparison of virus growth in bovine- and rabbit-origin cells
The virus titers of collected samples from MDBK-SY are shown in Fig. 3A and B. The titers of RK13/E+ and No.12/E+ strains were higher than those of their respective E− counterparts, starting from 3 and 4 days pi (dpi), respectively. Under the same conditions, only the RK13/E+ and RK13/E− strains exhibited the ability to grow in primary RK cells, reaching titers of up to ≥104.3 and ≥103.8 TCID50/mL, respectively (Fig. 3C). Neither the No.12/E+ strain nor the No.12/E− strain was detectable in primary RK cells. Both RK13 cell line-origin viruses plateaued after 48 h, with virus maintenance gradually decreasing thereafter. Moreover, each RK13 cell line-origin virus exhibited a similar growth curve pattern in the supernatant and cellular fractions of MDBK-SY and primary RK cells.

Growth kinetics of BVDV/E+ and BVDV/E− viruses in MDBK-SY and RK cells. Titers of BVDV/E+ and BVDV/E− were estimated using MDBK-SY cells by the END and interference method, respectively. The lowest estimated BVDV titer by either method was 101.8 TCID50/mL. Extracellular and intracellular viruses were harvested at 0, 1, 2, 3, 4, 5, and 6 days pi. The x-axis represents the number of days of sample collection, whereas the y-axis represents the virus titer expressed as log10 TCID50/mL. (A) Growth curve of RK13/E+ and E− strains inoculated at an MOI of 0.01 onto MDBK-SY cell monolayers. The symbols ● and ○ represent RK13/E+ titers in the supernatant and cellular fraction, whereas ▼ and ∆ represent RK13/E− titers in the supernatant and cellular fraction, respectively. (B) Growth curve of No.12/E+ and E− strains inoculated at an MOI of 0.01 onto MDBK-SY cell monolayers. The symbols ● and ○ represent No.12/E+ titers in the supernatant and cellular fraction, whereas ▼ and ∆ represent No.12/E− titers in the supernatant and cellular fraction, respectively. (C) Growth curve of RK13/E+, /E−, No.12/E+ and /E− strains at an MOI of 0.01 in RK cell monolayers. RK13 isolates demonstrated growth, as represented by solid lines. The symbols ● and ○ represent RK13/E+ titers in the supernatant and cellular fractions, whereas ▼ and ∆ represent RK13/E− titers in supernatant and cellular fraction, respectively. Meanwhile, No.12/E+ and /E− strains (represented by crossed and dotted lines) were not detected by the conventional END and interference method, respectively. (D) Persistence of parent RK13, RK13/E+, RK13/E−, No.12/E+, and No.12/E− strains in RK cells over five passages. The x-axis represents the passage number in RK cells. The symbols ■, ●, and ○ represent the titers of parent Rk13, RK13/E+, and RK13/E− strains in the supernatant of infected RK cells, respectively. No.12/E+ and No.12/E− strains, represented by crossed and dotted lines, were not detected by conventional methods
Furthermore, primary RK cells infected with RK13 parent, RK13/E+, RK13/E−, No.12/E+, and No.12/E− strains underwent five subsequent passages, and culture fluids were titrated. Only cells infected with parent RK13, RK13/E+, and RK13/E− strains supported BVDV maintenance, while No.12/E+ and No.12/E− strains were undetectable (Fig. 3D).
Comparison of cell susceptibility to RK13 cell line-origin viruses and No.12 viruses
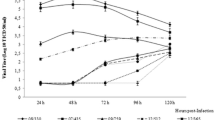
At a starting titer of 105 TCID50/mL, RK13/E+, RK13/E−, No.12/E+, and No.12/E− viruses were prepared in tenfold dilutions and inoculated, in parallel, onto monolayers of MDBK-SY, primary BT, and primary RK cells in 96-well plates. Virus titers were determined in a similar manner by the END and interference methods. Primary BT and RK cells exhibited the greatest and second greatest sensitivity for RK13/E+ and RK13/E− strains, respectively, exceeding that of MDBK-SY cells (Fig. 4). Similarly, primary BT cells were also the most sensitive to infection by No.12/E+ and No.12/E− strains, exceeding that of MDBK-SY cells. In contrast, primary RK cells showed a titer of up to 102.1 TCID50/mL for the No.12/E− strain, but nothing for the No.12/E+ strain (Fig. 4).

Susceptibility of MDBK-SY, BT and RK cells to BVDV (RK13/E+ and /E−, No.12/E+ and /E− viruses). Virus titers were determined by the END and interference method for BVDV/E+ and BVDV/E− viruses, respectively. The lowest estimated BVDV titer detection limit, by either method, was 101.8 TCID50/mL. The y-axis represents virus titer expressed as log10 TCID50/mL, whereas the x-axis represents cell culture types. The order of cell sensitivity for RK13 strains, from high to low, was BT, RK, and MDBK-SY cells. RK cells were not susceptible to No.12/E+, whereas low sensitivity (about 102.1TCID50/mL) was observed for No.12/E− virus
Recovery of virus from rabbits experimentally infected with RK13/E+ and RK13/E− strains
The detection of virus was demonstrated by RT-PCR and the END and interference methods from rabbit samples inoculated with the RK13/E+ and RK13/E− strains, respectively (Table 3). The RT-PCR results were positive for the presence of virus in all organs of rabbits inoculated with the RK13/E+ strain, and all organs except for WBC of rabbits inoculated with the RK13/E− strain. Alternately, viruses were detected by the END method in spleen, MLN, and liver, but not in kidney, pancreas, and WBC of samples infected with the RK13/E+ strain. The virus titer was approximately 102.1 TCID50/mL in all detected samples. Active viruses were undetectable in all rabbit samples except for MLN infected with the RK13/E− strain by the interference method.
Discussion
The RK13 cell line (ATCC) has been reported previously to be persistently infected with BVDV [3, 17, 18, 43]. In this study, we investigated BVDV quasispecies in the RK13 cell line by means of active virus isolation and analyzed their characteristics.
The BVDV isolated from the RK13 cell line showed the END phenomenon and could be titrated by the END method, but not by the interference method with VSV. This indicated that the virus persistently propagating in the RK13 cell line is the only virus showing the END phenomenon, but not the interference phenomenon. It was designated as the parental RK13 strain of BVDV. Unlike previous reports of BVDV contamination, which mostly focused on avoidance of contamination and reported detectable traces of virus RNA or defective particles [3], BVDV detected by the END method in the RK13 cell line appears to have developed the ability to grow in the RK13 cell line, which was demonstrated throughout 31 passages, and stability in terms of reproductive virus titer (104.5±0.7 TCID50/m) and proportion of positively infected cells (71.9 ± 3.12 %).
At first glance, the parent RK13 virus did not appear to contain any END− virus, while sharing the same END+ characteristics as END+ virus. However, END− virus was isolated from the parent RK13 virus by employing the RPF method, a unique tool for the detection of active quasispecies and isolation of a minor population of virus showing intrinsic interference [13] from the major population of virus showing the CPE or END phenomenon. This result indicates that the END− virus most likely exists at low levels within the parent RK13 virus stock. The END− and END+ viruses derived from parent RK13 virus were named the RK13/E− and RK13/E+ strain, respectively.
As shown in Table 1, an identical antigenicity profile was observed between RK13/E+ strains and RK13/E− strains. However, both strains demonstrated a significant difference in antigenicity compared with those of Nose (genotype 1) and KZ91CP (genotype 2). The antigenicities of Nose and KZ91CP strains were estimated as described previously [48].
The nucleotide sequences of the 5′-UTR in parent RK13, RK13/E+, and RK13/E− strains were determined, and phylogenetic analysis was performed. As a result, the nucleotide sequences of the 5′-UTRs of all of the strains were found to be completely identical (100 % homology), and all RK13 cell line-origin strains were classified as BVDV1b, along with the BVDV Osloss and CP7 strains (Fig. 2). Although there are clear biological differences between the RK13/E+ and RK13/E− isolates, these results suggest that each quasispecies might be derived from the same parental strain.
The conserved 5′-UTR of pestivirus has been widely used in studies in evolution, epidemiology, and taxonomy [15, 23, 46]. From a phylogenetic analysis of 34 strains, previously classified as BVDV1b, RK13 cell line-origin strains were subdivided into the same cluster as Argentinean and American field strains that were isolated in outbreaks [23, 53], and they were distinct from the Japanese isolates used in this study (data not shown).
Complete nucleotide sequences of the ORF region in RK13/E+ and RK13/E− strains were determined, and it was found that the genome constructions of both strains were consistent, with the characteristics of noncytopathogenic BVDV genomes. Although each strain had different biological characteristics, surprisingly, only four amino acid differences were discovered, located in Npro, E2, NS3, and NS5B (Table 2). Substitutions were located in (1) Npro, the BVDV autoprotease; (2) E2, an epitope region that plays a significant role in mediating receptor binding and virus entry, although both strains demonstrated similar growth ability in rabbit cells; (3) NS3, a highly conserved region in BVDV, responsible for helicase, protease and RNase activity; and (4) NS5B, the region encoding the RNA-dependent RNA polymerase. Of these, only the substitution in Npro is expected to have a potential effect with regard to interference of heterologous viruses. Several reports have shown that Npro can interfere with interferon-β (IFN-β) synthesis by inducing proteasomal degradation of INF regulatory factor 3 during CSFV and BVDV infection [34–42]. In the case of CSFV, END+ strains suppress the induction of IFN in cell cultures, whereas END− strains lose this ability, which may be attributed to Npro amino acid substitutions at positions 112 and 136 [45]. Therefore, one of the main differences between BVDV END+ and END− strains should be the latter strain’s ability to induce interferon in cell cultures. We speculate that the key difference between the RK13 END+ and END− strains, located in the Npro region, may be similar to those observed in CSFV strains, and further studies should therefore be carried out.
Since BVDV can naturally infect ruminants and swine, RK13 cells might have been contaminated with BVDV present in biological materials that were used when culturing the cells. Not only has a high rate of detection (57 %) of pestivirus genes in FBS samples been reported, but it has also been reported that infectious BVDV can be isolated from commercially available FBS [26]. Therefore, the results in this study support the hypotheses that BVDV in FBS most likely (1) contaminated a rabbit kidney cell line long before it was first reported [3], and (2) maintained persistent infections in different host cell lines for more than 20 years.
There appears to be evidence for the greater propagation ability of the RK13/E+ strain and the predominance of the /E+ quasispecies over its /E− counterpart. Inaba et al. [19, 20] previously reported that most BVDV isolates are END+ viruses. The isolated RK13/ E+ and RK13/E− strains, replicating in MDBK-SY bovine-origin cell cultures, demonstrated different growth curve patterns and virus titers (Fig. 3A). The RK13/E+ strain demonstrated greater growth ability than the RK13/E− virus, which was clearly evident after day 3 in a growth curve comparison, at an MOI of 0.01. Moreover, inoculation of RK13/E+ and RK13/E− strains, at a 100-fold higher MOI of 1, onto MDBK-SY and primary BT cells, in parallel, resulted in a ≥tenfold higher level of the RK13/E+ strain than RK13/E− strain after 1 dpi in all cells (data not shown). Additionally, serving as controls, the level of the No.12/E+ strain was also higher than that of the No.12/E− strain in terms of growth curves, which is yet another indicator of the greater amplification ability of the BVDV-END+ virus. All of these results support the predominance of the END+ characteristic observed for parent RK13 virus, which may be primarily due to the majority of RK13/E+ quasispecies in the virus population. They maintain different virus-cell interactions, demonstrating the ability to make the infected cell block homologous viruses only, or both homo- and heterologous viruses.
It is known that there is a significant difference in the growth abilities among BVDV in terms of their replication in various cell types [38]. The ability of BVDV to enter and replicate in vitro in cell cultures from different host species is relatively or strictly limited [40, 41, 44]. Only RK13 cell line-origin viruses, amongst the BVDV isolates examined in this study (No.12/E+ and No.12/E−, Nose/E+ and Nose/E−, and KZ91NCP/E+) demonstrated growth ability and reproducibility in primary RK cells, and this may be an indicator of their adaptability to rabbit cell culture in vitro. It is possible that this adaptation occurred and was acquired at the infection persistence stage during establishment of the RK13 cell line. Fernelius et al. [12] managed to obtain growth adaptability for BVDV (NADL and C24V) strains after more than 10 passages in a mixture of human and rabbit cells, but only after frequent passages in other cultures. However, it is also possible that, amongst the BVDV strains, such as No.12/E+, No.12/E−, Nose/E+, Nose/E−, and KZ91NCP/E+ strains, deemed unable to grow in rabbit-origin cultures, there might have been a very low titer. A slight difference (≤2-fold) between the RK13/E+ and RK13/E− strains was noted in the cell susceptibility test in MDBK-SY and primary RK cells. This would indicate that, although virus infectivity may be similar between the two strains, there may be other factors inhibiting or limiting the ability of RK13/E− virus to replicate to high levels.
To examine the infectivity of RK13 cell line-origin viruses in rabbits, the RK13/E+ and RK13/E− strains were each used to infect a rabbit. The RT-PCR results were positive for the presence of virus in all organ samples of the rabbit inoculated with the RK13/ E+ strain, and in all organ/tissue samples except for WBC of the rabbit inoculated with the RK13/E− strain. However, since only one rabbit was used for each strain, the overall result is preliminary. The results of this animal experiment have to be viewed with caution, and future studies with more animals are needed to confirm these preliminary results.
However, infectious viruses were detected by the END method in spleen, MLN, and liver, but not in kidney and pancreas, of the rabbit inoculated with the RK13/E+ strain. On the other hand, infectious viruses were not detected by the interference method, except in the MLN, of the rabbit inoculated with the RK13/E− strain. It has been confirmed that the RK13/E+ strain propagates well in rabbits, but the RK13/E− strain does not propagate as well as in cell culture.
In conclusion, RNA viruses have long been known to adapt by producing quasispecies, and this phenomenon has been utilized for decades in important biological applications such as the production of attenuated live-virus vaccines [16, 31]. Our results demonstrate that the BVDV quasispecies detected in the RK13 cell line is not composed of an individual variant with a single serological characteristic. Instead, the RK13 cell line contains RK13/E+ and RK13/E− strains, exhibiting different biological properties such as END and interference phenomena and virus-cell interactions but identical antigenicity and identical 5′-UTR nucleotide sequences. These viruses are able to replicate in vitro by transient and/or persistent infection of rabbit cell cultures, unlike other BVDVs that have been examined. Therefore, these quasispecies isolated from the RK13 cell line, especially the RK13/E− strain, may provide key insights and have potential future use for the process of generating a BVD-MD vaccine, perhaps in a similar manner to the GPE− strain, which has been shown to induce competent protection against classical swine fever [39, 47, 49]. Further studies on these virus quasispecies may also have benefit for elucidating the mechanism behind the diversity of BVDV and development of a live attenuated BVDV vaccine. In addition, The RPF method might be useful as a tool for the detection of an active minor population of viruses when researching quasispecies of not only pestiviruses but also other RNA viruses such as dengue virus [29] and other viruses in the genus Flavivirus.
References
Audet SA, Crim RL, Beeler J (2000) Evaluation of vaccines, interferons and cell substrates for Pestivirus contamination. Biologicals 28(1):41–46
Baker JA, York CJ, Gillespie JH, Mitchell CB (1954) Virus diarrhea of cattle. Am J Vet Res 15(57):525–531
Bolin SR, Ridpath JF, Black J, Macy M, Roblin R (1994) Survey of cell lines in the American Type Culture Collection for bovine viral diarrhea virus. J Virol Methods 48:211–221
Collins ME, Desport M, Brownlie J (1999) Bovine viral diarrhea virus quasispecies during persistent infection. Virology 259(1):85–98
Deng R, Brock KV (1992) Molecular cloning and nucleotide sequence of a pestivirus genome, noncytopathic bovine viral diarrhea virus strain SD-1. Virology (2):867–869
Deregt D, Jacobs RM, Carman PS, Tessaro SV (2004) Attenuation of a virulent type 2 bovine viral diarrhea virus. Vet Microbiol 100(3–4):151–161
Dezengrini R, Weiblen R, Flores EF (2006) Selection and characterization of canine, swine, and rabbit cell lines resistant to bovine viral diarrhea virus. J. Virol Methods 137(1):51–57
Drew T (2004) Bovine viral diarrhea manual of diagnostic tests and vaccines for terrestrial animals (mammals, birds and bees) vol 2, 5th edn. OIE, Paris pp 1051–1063
Eigen M (1971) Selforganization of matter and evolution of biological Macromolecules. Naturwissenschaften 58(10):465–523
Eigen M (1993) Viral quasispecies. Sci Am 269(1):42–49
Eigen M (1996) On the nature of virus quasispecies. Trends Microbiol 4(6):216–218
Fernelius AL, Lambert G, Hemness GJ (1969) Bovine viral diarrhea virus-host cell interactions: adaptation and growth of virus in cell lines. Am J Vet Res 30(9):1561–1572
Fukusho A, Ogawa N, Yamamoto H, Sawada M, Sazawa H (1976) Reverse plaque formation by hog cholera virus of the GPE− strain inducing heterologous interference. Infect Immun 14:332–336
Fukusho A, Ogawa N, Yamamoto H, Sawada M, Sazawa H (1975) Interference method with vesicular stomatitis virus for detection of GPE strain of hog cholera virus. Annu Rep Nat Vet Assay Lab 12:9–14
Harasawa R, Giangaspero M (1998) A novel method for pestivirus genotyping based on palindromic nucleotide substitutions in the 5′-untranslated region. J Virol Methods 70:225–230
Holland JJ, de la Torre JC, Clarke DK, Duarte E (1991) Quantitation of relative fitness and great adaptability of clonal populations of RNA viruses. J Virol 65:2960–2967
Horner GW (2004) Equine viral arteritis control scheme: a brief review with emphasis on laboratory aspects of the scheme in New Zealand. N Z Vet J 52(2):82–84
Horzineck MC (1981) Non-arthropod borne Togaviruses. Academic Press Inc., London
Inaba Y, Omori T, Kumagai T (1963) Detection and measurement of non-cytopathogenic strains of virus diarrhea of cattle by the END method. Arch Ges Virusforsh 13:425–429
Inaba Y, Tanaka Y, Kumagai T, Omori T, Ito H, Matumoto M (1968) Bovine diarrhea virus. II. END phenomenon: exaltation of Newcastle disease virus in bovine cells infected with bovine diarrhea virus. Jpn J Microbiol 12:35–49
Itoh O, Sugiyama M, Nakamura S, Sasaki H (1984) Characterization of non-pathogenic agent isolated from cytopathogenic bovine viral diarrhea-mucosal disease virus stock. Microbiol Immunol 28:1163–1167
Izawa H, Nagabayashi T, Soekawa M (1971) A mutant of hog cholera virus in the virus-carrier cell cultivated at a lower temperature. Zentralbl Veterinaermed B 18:197–204
Jones LR, Zandomeni R, Weber EL (2002) Quasispecies in the 5′ untranslated genomic region of bovine viral diarrhoea virus from a single individual. J Gen Virol. 83(P9):2161–2168
Kalaycioglu AT (2007) Bovine viral diarrhoea virus (BVDV) diversity and vaccination. Vet Q 29:60–67
Kiss I, Ros C, Kecskeméti S, Tanyi J, Klingeborn SB, Belák S (1999) Observations on the quasispecies composition of three animal pathogenic RNA viruses. Acta Vet Hung 47(4):471–480
Kozasa T, Aoki H, Nakajima N, Fukusho A, Ishomaru M, Nakamura S (2011) Methods to select suitable fetal bovine serum for use in quality control assays for the detection of adventitious viruses from biological product. Biologicals 39:242–248
Kumagai T, Shimizu T, Ikeda S, Matumoto M (1961) A new in vitro method (END) for detection and measurement of hog cholera virus and its antibody by means of effect of HC 179 virus on Newcastle disease virus in swine tissue culture. I Establishment of standard procedure. J Immunol 87:245–256
Kumagai T, Shimizu T, Matumoto M (1958) Detection of hog cholera virus by its effect on Newcastle disease virus in swine tissue culture. Science 128 (3320):366
Kurosu T (2011) Quasispecies of dengue virus. Trop Med Health 39(4 Suppl):29–36
Kärber G (1931) Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Arch exp Pathol Pharmakol 162:480–483
Lauring A, Andino R (2010) Quasispecies theory and the behavior of RNA viruses PLoS Pathogens. (Review);6(7):e1001005
Levings RL, Wessman SJ (1991) Bovine viral diarrhea virus contamination of nutrient serum, cell cultures and viral vaccines. Dev Biol Stand 75:177–181
Liess B, Moening V (1990) Ruminant pestivirus infection in pigs. Revue scientifique et Technique, Office International epizooties 9:151–161
Lindenbach BD, Thiel H-J, Rice CM (2007) Fields virology, In: Knipe DM, Howley PM (eds) Flaviviridae: the virus and their replication, vol 1, 5th edn, Lippincott William & Wilkins, Philadelphia, pp 1101–1151
Nakamura S, Fukusho A, Inoue Y, Sasaki H, Ogawa N (1993) Isolation of different non-cytopathogenic bovine viral diarrhea (BVDV) viruses from cytopathogenic BVD virus stocks using reverse plaque formation method. Vet Microbiol 38(1–2):173–179
Nakamura S, Shimazaki T, Sakamoto K, Fukusho A, Inoue Y, Ogawa N (1995) Enhanced replication of orbiviruses in bovine testicle cells infected with bovine viral diarrhoea virus. J Vet Med Sci 57(4):677–681
Nuttall PA, Luther PD, Stott EJ (1977) Viral contamination of bovine foetal serum and cell cultures. Nature 266(5605):835–837
Odeón AC, Leunda MR, Faverín C, Boynak N, Vena MM, Zabal O (2009) In vitro amplification of BVDV field strains isolated in Argentina: effect of cell line and culture conditions. Rev Argent Microbiol 41(2):79–85
Okaniwa A, Nakagawa M, Shimizu Y, Furuuchi S (1969) Lesions in swine inoculated with attenuated hog cholera viruses. Natl Inst Anim Health Q (Tokyo) 9(2):92–103
Paton DJ (2001) Host range, diversity and classification of pestiviruses. In: Proceedings of international Workshop organised by the EDQM, Paris, pp 13–16
Paton DJ (1995) Pestivirus diversity (review). J Comp Pathol 112:215–236
Peterhans E, Bachofen C, Stalder H, Schweizer M (2010) Cytopathic bovine viral diarrhea viruses (BVDV): emerging pestiviruses doomed to extinction. Vet Res 41(6):44
Potts BJ, Sawyer M, Shekarchi JC, Wismer T, Huddleston D (1989) Peroxidase labeled primary antibody method for detection of pestivirus contamination in cell cultures. J Virol Methods 26(1):119–124
Roehe PM, Edwards S (1994) Comparison of pestivirus multiplication in cells of different species. Res Vet Sci 57(2):210–214
Ruggli N, Summerfield A, Fiebach AR, Guzylack-Piriou L, Bauhofer O, Lamm CG, Waltersperger S, Matsuno K, Liu L, Gerber M, Choi KH, Hofmann MA, Sakoda Y, Tratschin JD (2009) Classical swine fever virus can remain virulent after specific elimination of the interferon regulatory factor 3-degrading function of Npro. J Virol 83:817–829
Sakoda Y, Ozawa S, Damrongwatanapokin S, Sato M, Ishikawa K, Fukusho A (1999) Genetic heterogeneity of porcine and ruminant pestiviruses mainly isolated in Japan. Vet Microbiol 65(1):75–86
Sasahara J, Kumagai T, Shimizu Y, Furuuchi S (1969) Field experiments of hog cholera live vaccine prepared in guinea-pig kidney cell culture. Natl Inst Anim Health Q (Tokyo) 9:83–91
Shimizu M, Satou K (1987) Frequency of persistent infection of cattle with bovine viral diarrhea-mucosal disease virus in epidemic areas. Jpn J Vet Sci 49:1045–1051
Shimizu Y, Furuuchi S, Kumagai T, Sasahara J (1970) A mutant of hog cholera virus inducing interference in swine testicle cultures. Am J Vet Res 31:1787–1794
Thiel H-J, Plagemann PGW, Moennig V (1996) Pestiviruses. In: Fields BN, Knipe DM, Howley PM (eds) Fields virology, vol 1, 3rd edn. Raven Press, New York, pp 1059–1073
Vilcěk S, Herring AJ, Nettleton PF, Lowings JP, Paton DJ (1994) Pestiviruses isolated from pigs, cattle and sheep can be allocated into at least three genogroups using polymerase chain reaction and restriction endonuclease analysis. Arch Virol 136:232–309
Yamamoto T, Kozasa T, Aoki H, Sekiguchi H, Morino S, Nakamura S (2008) Genomic analyses of bovine viral diarrhea viruses isolated from cattle imported into Japan between 1991 and 2005. Vet Microbiol 127(3–4):386–391
Yan L, Zhang S, Pace LW, Wilson FD, Wan H, Zhang M (2011) Combination of reverse transcription real-time polymerase chain reaction and antigen capture enzyme-linked immunosorbent assay for the detection of animals persistently infected with Bovine viral diarrhea virus. J Vet Diagn Invest 23(1):16–25
Acknowledgments
This work was partially supported by Japan International Cooperation Agency (JICA). The authors thank Dr. Peter Lee for his kind help with the preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Muhsen, M., Aoki, H., Ikeda, H. et al. Biological properties of bovine viral diarrhea virus quasispecies detected in the RK13 cell line. Arch Virol 158, 753–763 (2013). https://doi.org/10.1007/s00705-012-1538-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-012-1538-x