
Abstract
The phenotypic differentiation of relic P. sylvestris in southern Europe and southwestern Asia was verified using thirty-two populations sampled from the Iberian Peninsula, Massif Central, Balkan Peninsula, Crimea and Anatolia. Twenty-one morphological and anatomical needle traits and 18 cone morphological characteristics were examined to describe the population diversity and differentiation. The needle characters were not correlated to those of cone. The differences between regions were significant based on 12 needle and 9 cone characteristics, suggesting spatial isolation. The differentiation between the Iberian and Anatolian populations was the highest, which indicates the isolation by distance. The high level of morphological differentiation was also found among Iberian populations, supporting the already known complex history of the species in that region. Populations within other regions were differentiated at lower levels; however, the West Anatolian populations differed morphologically from the eastern ones. The described pattern of morphological differentiation supports the idea of the long-lasting existence of P. sylvestris in the south-European and Anatolian mountain regions. To conserve this variation, seed transfer between regions in the forest economy should be restricted.
Similar content being viewed by others

Introduction
The southern regions of Europe, the Iberian, Apennine and Balkan Peninsulas, together with the Crimea, the Anatolian mountains and the Caucasus are generally accepted as refugial areas, where most tree species survived the cold periods of the Pleistocene (e.g., Hewitt 1996; Willis et al. 1998; Willis and van Andel 2004; Médail and Diadema 2009; Soto et al. 2010). The populations of trees, P. sylvestris among others, which currently exist in refugial areas, have a relic nature and have maintained part of their species variation, which was not greatly disturbed by the influence of the Pleistocene glaciations, representing till now the unique Tertiary gene pool (e.g., Mirov 1967; Staszkiewicz 1993; Sinclair et al. 1999; Soranzo et al. 2000; Petit et al. 2003; Prus-Glowacki et al. 2003, 2012; Cheddadi et al. 2006; Pyhäjärvi et al. 2007). The persistence of the species in “small refugia” closer to the ice sheet can additionally complement the contemporary variation and diversity observed (Björkman et al. 2002; Bhagwat and Willis 2008; Binney et al. 2009). The relic character of Mediterranean populations of P. sylvestris and their isolation from each other and from the boreal portion of the area of distribution resulted in the description of several taxa and their treatment as independent species, subspecies or varieties (Gaussen 1960; Mirov 1967). The isolation of the southernmost populations of P. sylvestris and occurrence in distinct plant communities in particular regions was also the basis for describing several “climatypes” (Svoboda 1953), which were sometimes treated as different taxa (e.g., Molotkov and Patlaj 1991). The subspecies P. sylvestris subsp. sylvestris from Western Europe and P. sylvestris subsp. hamata (Steven) Fomin from the Crimea and Anatolia were distinguished. The latter subspecies has cone scales with convex, hooked umbos, bluish green needles with 4–8 resin ducts and a relatively small percentage of fibrous sclerenchyma cells around the vascular bundles (Molotkov and Patlaj 1991). Within P. sylvestris subsp. sylvestris, several varieties were described on the Iberian Peninsula (Gaussen 1960). The taxonomic subdivision of P. sylvestris, however, was discontinued in the last decades (do Amaral Franco 1986; Christensen 1997; Farjon 2010). The systematic status of the most of taxa distinguished within P. sylvestris has not been tested using molecular markers so far. However, some of them differ from all the other (Prus-Glowacki et al. 2003, 2012).
The greater genetic distinctiveness of the Iberian and Anatolian populations in comparison to the populations from central and northern Europe has been reported several times (Tobolski and Hanover 1971; Mejnartowicz 1979; Prus-Głowacki and Stephan 1994; Prus-Glowacki et al. 2003; Semiz et al. 2007). Pyhäjärvi et al. (2007) have lately shown significant differences between Turkish and Spanish Scots pine populations in both nuclear and mitochondrial DNA. Conversely, a low level of genetic differentiation (F ST = 0.014) among several populations from Turkey was shown on the basis of isoenzymes (Turna 2003; Bilgen and Kaya 2007). The degree of population differentiation in Bulgaria revealed by cpDNA was not very high (F ST = 0.048) (Naydenov et al. 2005). Prus-Glowacki et al. (2012) described values of inter-population variation for the Iberian Peninsula (G ST = 0.039), Scotland (G ST = 0.050) and the Balkan Peninsula (G ST = 0.058) against the continuous range of the species, where the value was lower (G ST = 0.029). In addition, differentiation among Italian populations tested using nuclear SSR markers revealed F ST = 0.058 (Belletti et al. 2012) or even F ST = 0.08 (Scalfi et al. 2009). The greater level of genetic differences between populations of P. sylvestris on the Iberian Peninsula is frequently treated as the effect of permanent, impermeable barriers to the transport of seeds and pollen (Sinclair et al. 1999; Soranzo et al. 2000; Prus-Glowacki et al. 2003, 2012). The barriers for effective gene flow within the Iberian, Apennine and Balkan Peninsulas may be the result of site and/or climate conditions that were unfavorable for pines during the glacial and interglacial periods of the Pleistocene. In particular, this could be the case in the mountains and in extremely dry areas, such as the wide valleys of rivers, like the Ebro in the Iberian Peninsula (Alía et al. 2001; Dzialuk et al. 2009) and the Po on the Apennine Peninsula (Labra et al. 2006).
Local gene flow via long distance pollen and/or seed dispersal could be more effective during the glacial periods of the Pleistocene than at present, at least in some regions (Cheddadi et al. 2006; Naydenov et al. 2007). The Black Sea, whose level during glacial periods of the Pleistocene was much lower, did not function as such a strong barrier to plant migration as at the present time (Yena et al. 2005). Therefore, greater gene flow from the core Scots pine populations to the Balkan and Anatolian P. sylvestris stands can be expected in comparison to the Iberian stands, which are separated from the north by the Pyrenees. Taking into account the taxonomic status of populations in particular regions (Molotkov and Patlaj 1991), the present geographic range of P. sylvestris (Boratyński 1991) and its Pleistocene history (Mirov 1967), we hypothesized that: (1) isolation of populations in Mediterranean refugial areas caused not only genetic differences, but was also a reason for differentiation at the phenotypic level and, consequently, there is high genetic and morphological differentiation between distant refugial areas, (2) there is a low level of phenotypic differentiation among the presently isolated populations historically derived from the same refugial area, lower than among groups of populations representing particular refugia, (3) the differences among Anatolian populations should be at a lower level than those among Iberian ones, (4) the differences between Anatolian and Crimean populations, representing subsp. hamata, should be high, when compared to all the other, of subsp. sylvestris. This study aimed to verify these hypotheses by determining the level of differentiation between natural populations of P. sylvestris basing on biometric data from needles and cones.
Materials and methods
Investigated area
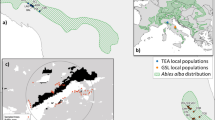
The populations of P. sylvestris included in the investigation were located in the mountains of Europe and Asia close to the Mediterranean and/or sub-Mediterranean regions, on or close to the refugial areas of the Tertiary floras (Médail and Diadema 2009). The material was sampled from populations of presumed natural origin (Coode and Cullen 1965; Horvat et al. 1974; do Amaral Franco 1986; Boratynski et al. 1992), in old stands, dispersed far from the southern limit of the main, boreal geographic range of Scots pine (Fig. 1; Table 1). Following published data on the genetic and morphological differentiation of P. sylvestris (Sinclair et al. 1999; Soranzo et al. 2000; Turna 2003; Labra et al. 2006; Bilgen and Kaya 2007; Pyhäjärvi et al. 2007, 2008; Naydenov et al. 2005, 2007; Prus-Glowacki et al. 2003, Prus-Glowacki et al. 2012; Turna and Güney 2009; Belletti et al. 2012), material was gathered from the Iberian Peninsula (IP), the Massif Central (MC), the Balkan Peninsula (BAL), Anatolia (AN) and the Crimea (CR). These five regions were treated as separate refugial areas of P. sylvestris in the subsequent comparisons.
Material and measurement procedures
The study comprised material from subsp. sylvestris var. nevadensis Christ, var. pyrenaica Svob. and var. iberica Svob. from the Iberian Peninsula, var. aquitana Schott from the Massif Central, var. illyrica Svob. and var. rhodopaea Svob. from the Balkan Peninsula and subsp. hamata var. subalpina Fomin from Crimea and N Anatolia (Molotkov and Patlaj 1991). In each stand, ca. 30 old, cone-bearing individuals separated from each other by 30–40 m were sampled. Ten two-year-old brachyblasts with needles with no visible insect and/or fungi damage, from well-illuminated, south-facing parts of the tree crown about 2- to 5-m above ground level were collected from each tree. After collection, the lengths of the two-year-old needles were measured and the plant material was then preserved in 70 % ethanol and stored at 20 °C until analysis (Boratynska et al. 2005; Boratyńska and Boratyński 2007). In total, 32 populations were sampled, from which 951 individuals were represented in the needle comparisons. 50 cones were collected as a population sample from 21 populations and conserved in a dry state for analysis. Each cone was picked up from the ground below the crown of a different old tree.
Semi-durable preparations from the central part of five two-year-old needles from each individual were made by a cross section. 21 characteristics of needles and 10 of cones (Tables 2, 3) were measured and/or evaluated. The needle anatomical traits were determined under a Nikon SMZ800 binocular and a JenaMed2 light microscope (Carl Zeiss, Jena) (Boratyńska and Bobowicz 2001). The cone traits were measured using an electronic caliper (Marcysiak 2005, 2006; Marcysiak and Boratyński 2007). Additional characteristics were assessed as ratios of measured characters describing the shapes of needle, cones or parts of these organs.
Statistical analyses
The Shapiro–Wilk’s test was used to verify the data distribution and the Brown–Forsythe test to assess the homoscedasticity of the data variance prior to multivariate comparisons. The Pearson’s correlation coefficient was analyzed between pairs of characters to detect possible redundant variables (Zar 1999; Sokal and Rohlf 2003). The data were standardized using STATISTICA (StatSoft) to avoid a possible influence of their different types on the result of the analyses.
Minimal and maximal values of characteristics were determined and arithmetical means, standard deviation and variation coefficients were calculated and analyzed for every population and for the IP, MC, BAL, AN and CR regions. Analysis of discrimination was used to detect groupings of samples separately for data from needles and from cones (Watała 2002; Sokal and Rohlf 2003). Similarly, agglomeration according to Ward’s method on the shortest Euclidean distances calculated for the synthetic and simple characteristics was used to describe the relationships among populations and to determine groups of populations (Sokal and Rohlf 2003).
Tukey’s test was used to assess the significance of differences in particular characteristics between regions for traits with normal distribution. The Kruskal–Wallis test was performed for traits with biased distribution to attest the statistical significance of the differences. The percentage data were arcsined before statistical analyses (Watała 2002). A hierarchical analysis of variance was performed for particular characteristics to determine the percentage of total variation and the statistical significance between groups of populations representing the five regions (Sokal and Rohlf 2003). STATISTICA 9.0 for Windows (StatSoft) and JMP (SAS Institute Inc.) software was used for the mathematical calculations.
The Mantel test for isolation by distance was performed by comparison of Euclidean and Mahalanobis’ distances to the geographic distances between populations (Mantel 1967), as implemented by GenAlEx (Peakall and Smouse 2006). The geographic distances were retrieved from geographic coordinates using Map Info 9.5 (Pitney Bowes). The most important barriers between populations were detected by the analysis of Euclidean and Mahalanobis’ distances as calculated by BARRIER 2.2 software (Manni et al. 2004; Dzialuk et al. 2009).
Results
Character verification
Most of the analyzed characteristics revealed normal or close to normal distribution. The variance in the raw data was not homoscedastic, but after standardization became more homogeneous. Both the normality and homoscedasticity of the transformed data allowed the use of multivariate analyses and parametric tests. The dimensional needle characters were generally dependent on each other; the cone and cone scale characteristics also correlated at a statistically significant level. In spite of this, no single trait was excluded in the step-wise discrimination analyses as highly redundant. For this reason, all the characteristics of the needles and cones were included in the multivariate analyses.
Correlations between needle and cone characteristics were generally found to be low; however, some of them were statistically significant. The numbers of stomata and resin canals correlated negatively, while the presence of cells with thin walls around resin canals and fiber-like cells between vascular bundles correlated positively with cone and cone scale sizes (Table 4). All these dependencies were weak.
Needle characteristics
The average values of characteristics (Table 2) differentiated in most instances at statistically significant levels between at least two of the compared regions, most frequently between IP and another (Table 5). The greatest number of traits differing at a statistically significant level was found between IP and AN, the lowest between BAL, CR and AN. The latter three regions appeared to be the most similar (Table 5).
On the plane between the first two discriminatory variables, the centroids of the populations were split into two groups by the first variable (U 1), which was responsible for more than 34 % of the variation. Populations from AN, the BAL and CR were shown to be separated from those representing IP. Populations from MC fell between these two groups (Fig. 2a). SL, SRC, VBS and TE were primarily responsible for this division. The second variable (U 2), which was responsible for more than 16 % of the total variation, differentiated the populations based on the IP and MC. Populations from the other regions revealed lower levels of differentiation (Fig. 2a).


Differentiation of Pinus sylvestris populations (acronyms as in Table 1) detected in the needle characters: a discrimination analysis among Iberian Peninsula (IP), Massif Central (MC), Balkan Peninsula (BAL), Anatolia (AN) and Crimea (CR), b agglomeration on the shortest Euclidean distances according to Ward’s method
Analysis of agglomeration on the shortest Euclidean distances calculated from the needle characters confirmed the results of the discrimination analysis and defined two main groups of populations, the first including the populations from the Iberian Peninsula and Massif Central, the second those from the Balkans, Crimea and Anatolia (Fig. 2b).
The hierarchical analysis of variance showed that 12 of the 21 needle characteristics differed among the groups of populations from IP, MC, BAL, CR and AN at a statistically significant level (Table 6). These characteristics revealed a greater percentage of the variance estimated between regions than between populations within a single region, which confirmed our hypothesis.
The correlations of Euclidean and Mahalanobis distances between populations, calculated from the needle characteristics, to geographic distances were high and statistically significant, with r 2 = 0.47, p < 0.001 and r 2 = 0.36, p < 0.001, respectively. The barriers constructed on the Euclidean and Mahalanobis’ distances confirmed high-level differences between the populations of P. sylvestris from the Iberian Peninsula and Massif Central. All the other compared populations appeared much more uniform (Fig. 3a, b).

Barriers between populations detected using Barrier 2.2 on the needle a, b and cone c, d characters and Euclidean a, d and Mahalanobis’ b, c distances between populations
Cone characteristics
The average values of cone characteristics (Table 3) differentiated at statistically significant levels between at least two or three of the compared regions (Table 7). Only the CVX/CCA ratio appeared completely stable throughout all of the regions compared. The greatest number of cone traits differentiated between IP and AN, and then between IP and BAL, while the most similar with respect to cone characteristics were IP and MC, which differed only in characteristics connected with CNS (Table 7).
Discrimination analysis based on the measured and/or evaluated characters did not reveal a clear pattern of population groupings (data not shown); however, using the proportions CL/CD, CL/CSN, LA/WA, LA/TA, CVX/CCA, CD/CSN, DAU/TA and CD/CDM (see Table 3) gave more interpretable results. On the plane between the first two discriminatory variables responsible for more than 61 % of the total variation, the centroids of the populations formed one dispersed group, but in the case of IB and MC they were slightly separated from BAL, CR and AN by the second variable (U 2) (Fig. 4a). The most separated population was ES_12. The particular regions, however, were also separated on the fields between the subsequent discriminatory variables (Fig. 5). Nevertheless, the high level of differences between the populations within the IP and AN regions should be stressed, especially by the first discriminatory variable U 1 (Figs. 4a, 5). The discriminatory variable U 1 was determined mostly by LA/WA, DAU/TA and CD/CDM, U 2 by CD/CSN and CL/CSN, U 3 by LA/WA, CD/CDM and CL/CD, U 4 by CD/CDM and CL/CSN, U 5 by CL/CD and CL/CSN and U 6 by DAU/TA. Finally, the most discriminating between regions appeared to be LA/WA, DAU/TA, CD/CDM, CD/CSN, CL/CSN and CL/CD. The Euclidean and Mahalanobis’ distances were statistically significant between all possible pairs of populations (data not shown). The population agglomeration on the shortest Euclidean distances retrieved from the cone characteristics did not show a clear geographic pattern. A high level of differentiation among Iberian and Massif Central populations was found, which were dispersed among Balkan, Crimean and Anatolian populations (Fig. 4b). Similar intermingled dispersion of the populations was observed on the agglomeration on the Mahalanobis’ distances (data not shown).

Differentiation of Pinus sylvestris populations (acronyms as in Table 1) detected in the cone characters: a discrimination analysis among Iberian Peninsula (IP), Massif Central (MC), Balkan Peninsula (BAL), Anatolia (AN) and Crimea (CR), b agglomeration on the shortest Euclidean distances according to Ward’s method

Differentiation of Pinus sylvestris populations detected in the cone characters: discrimination analysis among Iberian Peninsula (IP), Massif Central (MC), Balkan Peninsula (BAL), Anatolia (AN) and Crimea (CR) among six first discriminatory variables
The hierarchical analysis of variance found 9 out of the 18 cone characteristics differed among the IP, MC, BAL, CR and AN population groups at a statistically significant level. TA, LA/TA and CD/CSN revealed a greater percentage of the estimated variance between regions than between populations within a single region, while the next six variances between regions were at similar or lower levels than those within regions (Table 8).
Mantel’s test on the Euclidean and Mahalanobis’ distances between populations calculated based on the cone characteristics revealed relatively small but statistically significant positive correlations to the geographic distances, r 2 = 0.23, p < 0.001 and r 2 = 0.28, p < 0.01, respectively. The barriers constructed on the Euclidean and Mahalanobis’ distances confirmed high levels of differentiation in the Iberian populations, but also allowed differences to be detected among Crimean, Anatolian and Balkan populations (Fig. 3c, d).
Discussion
Differentiation between regions
The multivariate analyses on the morphological and anatomical characteristics of the needles of P. sylvestris showed differences between the Iberian group of populations in relation to the populations from Anatolia, the Balkans and the Crimea. Populations from Massif Central had an intermediate nature, but were more similar to those from the East (Fig. 2a, b). This is generally congruent with the results of genetic differentiation (Pyhäjärvi et al. 2007), but it should be stressed that the needle characteristics indicated, first of all, the great differences in the Iberian populations of P. sylvestris when compared to the populations from the other studied regions.
The result of multivariate analysis using the cone characteristics did not appear as clear as in the case of the needles. However, the distribution of the compared populations between subsequent pairs of discriminatory variables allowed populations and/or their groups from five compared regions to be distinguished. The differences are probably blurred by the high levels of cone variation, generally higher than that of needles (Boratynska et al. 2005), and the high level of variation of the cone characteristics among populations from IP and AN determined by the first discriminatory variable (Fig. 4a). The slightly different patterns of population differentiation based on the needle and cone characters can be explained by the lack of correlation between them and, consequently, independent adaptation and evolution of cones and needles to the local environment in particular populations and regions. Indeed, only a few cone characteristics correlated to needle traits in our study and the significant correlations found were weak (Table 4).
Generally, the most important barrier divided the eastern and western populations, confirming the hypotheses regarding migration history (Petit et al. 2003; Prus-Glowacki et al. 2012). The IP and MC populations could contact during cold periods of the Pleistocene (Benito Garzón et al. 2007; Rubiales et al. 2010; Prus-Glowacki et al. 2012), similar to the case of P. uncinata (Dzialuk et al. 2009). The differences found between groups of populations supposedly representing P. sylvestris subsp. sylvestris (IP, MC, BAL) and P. sylvestris subsp. hamata (AN and CR), in light of our results, do not support the validity of these two taxa.
Characters differentiating between regions
The most differentiating needle characteristics in the discrimination analysis were the number of sclerenchymatic layers above phloem (SL), the number of stomata rows on the convex side of the needle (SRC), the presence of fibrous and semi-fibrous cells between the vascular bundles (VBF and VBS) and the thickness of the epidermal cells (TE). Interestingly, the mentioned characteristics quite clearly differentiate the Iberian populations from all the others (see Tables 2, 4). The other set of characteristics differentiating between regions revealed a hierarchical analysis of variance (Table 5). Among differentiating characteristics, we did not find the number of resin canals (RCC + RCF), reported as specific for subsp. hamata (Molotkov and Patlaj 1991). All the regions had RCC + RCF of about 9–10 except for the Iberian and Anatolian populations, which were characterized by lower values (Table 2). The low numbers of resin canals in needles of the Iberian population (var. pyrenaica and var. iberica) and from Massif Central (var. aquitana) was reported by Gaussen (1960). In conclusion, we shall state that a small number of resin canals are characteristic of the southernmost, marginal populations of P. sylvestris. The same also concerns silvery needles, which prevail not only in the Caucasus but also in the southernmost Iberian populations (Gaussen 1960; Boratyńska and Hinca 2003).
The numbers of resin canals in populations of P. sylvestris close to the Mediterranean appeared smaller than those found in Central European populations, where values of this character oscillated between (11) 12 and 15 (compare Table 2 with, e.g., data of Bobowicz and Radziejewska 1989, Table 1; Bobowicz et al. 1994, 1995, Table 1; Boratynska et al. 2008, Table 1). The number of resin canals also depends on the age of the trees and is significantly lower in the needles of seedlings and young individuals than in adult trees (Lin et al. 2001; Boratynska et al. 2008). Using needles from only adult or even very old trees, we avoided the influence of this source of variation, which, however, could be a source of the differences reported in the papers by other authors.
The small percentage of the sclerenchymatic cells in the tissue between vascular bundles of needles of P. sylvestris subsp. hamata, indicated by Molotkov and Patlaj (1991), the fibrous and semi-fibrous cells (VBF and VBS) in our results appeared similar in whole regions (about 70–80 %), but varying between populations; they were highly variable with standard deviations reaching even 70 % of the mean values of the characteristic. Only populations from the Iberian Peninsula had a smaller value of VBF + VBS (Tables 2, 4). The data for Mediterranean populations of P. sylvestris, however, had lower values for these characteristics than were found in populations from the boreal part of the species range (Boratyńska and Boratyński 2007, Figs. 6 and 7; Boratyńska et al. 2008, Table 1).
The differentiation of cones appeared multivariate, but the greatest differences between regions appeared in traits concerning the size of the cone (CL, CD, CDM and its proportions) and cone apophysis (LA, TA and DAU and its proportions). The latter characteristics are connected with hooked umbo, described by the characteristics TA and DAU. The highest values for these traits were found in the Anatolian, Crimean and Balkan populations. The high proportion of cone scales with thick apophysis can be interpreted as confirmation of subsp. hamata; however, this type of cone was not only detected in the Anatolia and Crimea where the subsp. hamata was described (Gaussen 1960; Molotkov and Patlaj 1991), but also in Balkan populations. Cones with thick, hooked umbos were also found in Iberian populations, but in lower percentages. This type of cone occurs as well in the natural populations of Massif Central (Staszkiewicz 1963, 1968).
The results of detailed study of P. sylvestris cones by Staszkiewicz (1961, 1963, 1968) made it possible to compare the values of cone traits in populations from different regions of the species’ geographic range, including the southernmost populations from our study. Generally, we have to conclude that cones from the southernmost populations of the species appeared larger than those from central and northern Europe and were composed of a greater number of scales (CSN). The scale apophyses (LA, WA) of most of the Balkan, Crimean and Anatolian populations frequently resembled the values described by Staszkiewicz (1968) for the subcarpatica type, while some from IB more resembled the scotica type. The higher values of cone characteristics found in our study may result from adaptation of the southernmost populations of P. sylvestris to higher temperatures.
Differentiation within regions
Nevertheless, the high level of differentiation in the Iberian populations found in the needle and cone characters indicate an adaptation to the local conditions and support the impression of possible long-lasting isolation in separate Iberian refugia (Rubiales et al. 2010). The high level of differences in Scots pine among Iberian Peninsula populations found in our study can be treated as confirmation of the earlier described patterns of the species variation. Differentiation of the Iberian populations has been described based on taxonomic status (e.g., Gaussen 1960; do Amaral Franco 1986; Klaus 1989), and has previously been found using needle characteristics (Pardos et al. 1990; Boratyńska and Hinca 2003; Jasinska et al. 2010) and cone characteristics (Staszkiewicz 1963, 1993; Marcysiak 2005). Similar differentiation was also described using isoenzymatic (Prus-Głowacki and Stephan 1994) and molecular markers (Prus-Glowacki et al. 2003, 2012; Robledo-Arnuncio et al. 2005). The populations from Sierra Nevada and Sierra de Baza were recognized as the most distinct of all the Iberian populations of P. sylvestris (do Amaral Franco 1986; Boratyński 1991; Prus-Glowacki et al. 2003). The distinctiveness of the population from Sierra de Baza was also confirmed in the present study, but only in regard to cone characteristics (Fig. 4).
Morphological differences were detected between the East and West Anatolian populations (Figs. 3, 5), which have not been biometrically documented till now. The pattern of morphological differentiation in the Anatolian populations of P. sylvestris is similar to those already described using isozyme variation (Bilgen and Kaya 2007). The isolation of the eastern from western Anatolian populations of P. sylvestris can be explained by the mountain ranges known as the “Anatolian diagonal”. This system connects the Pontic Mountains with the Anti-Taurus and allowed migration of species during the Pleistocene climate changes, as for example Cedrus libani (Davis et al. 1971; Ekim and Güner 1986; Dogan et al. 2010), but possibly also formed a barrier between the western and eastern populations of P. sylvestris. The morphological pattern of diversity in Anatolian populations of the species may also result from: (1) another origin source with the western populations from the Balkans and the eastern ones from the Caucasus, (2) different rates of evolution in the two regions following the same adaptation processes. Generally, the same processes of diversity formation have been suggested in the comments to genetic investigations on diverse types of molecular markers (e.g., Pyhäjärvi et al. 2007, 2008; Floran et al. 2011; Prus-Glowacki et al. 2003, 2012).
Conclusion
The phenotypic differentiation of isolated populations of P. sylvestris in southern Europe and southwestern Asia is high. The differences among populations depend on the geographic distances, which are similar to the genetic differentiation of the species. The highest level of differences was found between the Iberian and Anatolian populations, while populations representing three other tested regions are generally intermediate. The high level of population differentiation argues for the use of local sources of seeds in forest management, especially in Anatolia and on the Iberian Peninsula, to conserve their biodiversity. Our result does not support the validity of P. sylvestris subsp. sylvestris and P. sylvestris subsp. hamata.
References
Alía R, Moro-Serrano J, Notivol E (2001) Genetic variability of Scots pine (Pinus sylvestris) provenances in Spain: growth traits and survival. Silva Fen 35:27–38
Belletti P, Ferrazzini D, Piotti A, Monteleone I, Ducci F (2012) Genetic variation and divergence in Scots pine (Pinus sylvestris L.) within its natural range in Italy. Eur J For Res 131:1127–1138
Benito Garzón M, Sánchez de Dios R, Sáinz Ollero H (2007) Predictive modelling of tree species distribution on the Iberian Peninsula during Last Glacial Maximum and Mid-Holocene. Ecography 30:120–134
Bhagwat SA, Willis KJ (2008) Species persistence in northerly glacial refugia of Europe: a matter of chance or biogeographical traits? J Biogeogr 35:464–482
Bilgen BB, Kaya N (2007) Allozyme variations in six natural populations of Scots pine (Pinus sylvestris L.) in Turkey. Biologia 62:697–703
Binney HA, Willis KJ, Edwards ME, Bhagwat SA, Anderson PM, Andreev AA, Blaauw M, Damblon F, Haesaerts P, Kienast F, Kremenetski KV, Krivonogov SK, Lozhkin AV, McDonald GM, Novenko EY, Oksanen P, Sapelko TV, Valiranta M, Vazhenina L (2009) The distribution of Late Quaternary woody taxa in northern Eurasia: evidence from a new macrofossil database. Quat Sci Rev 28:2445–2464
Björkman L, Feurdean A, Cinthio K, Wohlfarth B, Possnert G (2002) Late glacial and early Holocene vegetation development in the Gutaiului Mountains, north-western Romania. Quat Sci Rev 8(9):1039–1059
Bobowicz AM, Radziejewska A (1989) The variability of Scots pine from Piekielna Góra as expressed by morphological and anatomical traits of needles. Acta Soc Bot Poloniae 58:375–384
Bobowicz AM, Korczyk AF, Breninek J (1994) Variability of Pinus sylvestris L. plus trees of eight Polish provenances in traits of needle morphology and anatomy. Biol Bull Poznań 31:33–49
Bobowicz AM, Pieczyńska B, Sufryd A, Breninek J, Korczyk AF (1995) Inter-provenance variability of half-sib Scots pine progenies in traits of needle morphology and anatomy. Biol Bull Poznań 32:43–57
Boratyńska K, Bobowicz AM (2001) Pinus uncinata Ramond taxonomy based on needle characters. Plant Syst Evol 227:183–194
Boratyńska K, Boratyński A (2007) Taxonomic differences among closely related pines Pinus sylvestris, P. mugo, P. uncinata, P. rotundata and P. uliginosa as revealed in needle sclerenchyma cells. Flora 202:555–569
Boratyńska K, Hinca M (2003) Morphological characteristic of Pinus sylvestris L. in the southernmost, isolated locality in the Sierra de Baza (S Spain) as expressed in the needle traits. Dendrobiology 50:3–9
Boratyńska K, Marcysiak K, Boratyński A (2005) Pinus mugo (Pinaceae) in the Abruzzi Mountains: high morphological variation in isolated populations. Bot J Linn Soc 147:309–316
Boratyńska K, Jasińska AK, Ciepłuch E (2008) Effect of tree age on needle morphology and anatomy of Pinus uliginosa and Pinus sylvestris—species-specific character separation during ontogenesis. Flora 203:617–626
Boratyński A (1991) Range of natural distribution. In: Giertych M, Mátyás C (eds) Genetics of Scots pine. Akadémiai Kiadó, Budapest, pp 19–30
Boratyński A, Browicz K, Zieliński J (1992) Chorology of trees and shrubs in Greece. Institute of Dendrology, Kórnik/Poznań
Cheddadi R, Vendramin G, Litt T, François L, Kageyama M, Lorentz S, Laurent JM, de Beaulieu JL, Sadori L, Jost A, Lunt D (2006) Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris. Glob Ecol Biogeogr 15:271–282
Christensen KI (1997) Pinus L. In: Strid A, Tan K (eds) Flora Hellenica 1. Koeltz Sc Books, Königstein, pp 4–9
Coode MJE, Cullen J (1965) Pinus L. In: Davis PH (ed) Flora of Turkey, 1. Edinburgh University Press, Edinburgh, pp 72–75
Davis PH, Harper PC, Hedge IC (1971) Plant life of South West Asia. The Botanical Society of Edinburgh, Edinburgh
do Amaral Franco J (1986) Pinus L. In: Castroviejo S, Laínz M, López González G, Montserrat P, Muñoz Garmendia F, Paiva J, Villar L (eds) Flora iberica, 1, Real Jardín Botánico, Madrid, pp 171–174
Dogan B, Duran A, Bagci Y, Dinc M, Martin E, Cetin O, Ozturk M (2010) Phylogenetic relationships among the taxa of the genus Johrenia D. C. (Apiaceae) from Turkey based on molecular method. Bangladesh J Plant Taxon 17(2):113–120
Dzialuk A, Muchewicz E, Boratyński A, Montserrat JM, Boratyńska K, Burczyk J (2009) Genetic variation of Pinus uncinata (Pinaceae) in the Pyrenees determined with cpSSR markers. Pl Syst Evol 277:197–205
Ekim T, Güner A (1986) The Anatolian diagonal: fact or fiction? Proc R Soc Edinburgh 89B:69–77
Farjon A (2010) A handbook of the World conifers. Brill, Leiden, pp 1–2
Floran V, Sestras RE, Garcís Gil MR (2011) Organelle genetic diversity and phylogeography of Scots pine (Pinus sylvestris L.). Not Bot Horti Agrobotnici Cluj Napoca 39:317–322
Gaussen H (1960) Les Gymnospermes actuelles et fossils. Trav Labor Forestal Touluse 4
Hewitt GM (1996) Some genetic consequences of ice ages, and their role in divergence and speciation. Biol J Linn Soc 58:247–276
Horvat I, Glavač V, Ellenberg H (1974) Vegetation Südosteuropas. Geobotanica selecta, 4th edn. G. Fischer, Stuttgart
Jalas J, Suominen J (1973) Atlas Florae Europaeae 2. In: The Committee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo, Helsinki
Jasińska AK, Wachowiak W, Muchewicz E, Boratyńska K, Montserrat JM, Boratyński A (2010) Cryptic hybrids between Pinus uncinata and P. sylvestris. Bot J Linn Soc 163:473–485
Klaus W (1989) Mediterranean pines and their history. Pl Syst Evol 162:133–163
Labra M, Grassi F, Sgorbati S, Ferrari C (2006) Distribution of genetic variability in southern populations of Scots pine (Pinus sylvestris L.) from the Alps to the Apennines. Flora 201:468–476
Lin J, Sampson DA, Ceulemans R (2001) The effect of crown position and tree age on resin-canal density in Scots pine (Pinus sylvestris L.) needles. Can J Bot 79:1257–1261
Manni F, Guerard E, Heyer E (2004) Geographic patterns of (genetic, morphologic, linguistic) variation: how barriers can be detected by using Monmonier’s algorithm. Hum Biol 76:173–190
Mantel N (1967) The detection of disease clustering and a generalized regression approach. Cancer Res 27:209–220
Marcysiak K (2005) Taxonomic position of Pinus sylvestris L. population from Sierra de Baza in Spain on the basis of cone characters. In: Prus-Głowacki W (ed) Variability and evolution—new perspectives. UAM, Poznań, Ser Biolog, vol 72, pp 391–398
Marcysiak K (2006) Scots pine (Pinaceae) from the Crimea compared to the species variation in Europe on the basis of cone traits. Phytol Balcan 12:203–208
Marcysiak K, Boratyński A (2007) Contribution to the taxonomy of Pinus uncinata (Pinaceae) based on cone characters. Pl Syst Evol 264:57–73
Médail F, Diadema K (2009) Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J Biogeogr 36:1333–1345
Mejnartowicz L (1979) Genetic variation in some isoenzyme loci in Scots pine (Pinus sylvestris) populations. Arbor Kórnickie 24:91–104
Mirov NT (1967) The genus Pinus. The Ronald Press Company, New York
Molotkov PI, Patlaj IN (1991) Systematic position within the genus Pinus and intraspecific taxonomy. In: Mátyás C (ed) Genetics of Scots pine. Akad Kiadó, Budapest, pp 31–40
Naydenov KD, Tremblay FM, Alexandrov A, Fenton NJ (2005) Structure of Pinus sylvestris L. populations in Bulgaria revealed by chloroplast microsatellites and terpenes analysis: provenance tests. Biochem Syst Ecol 33:1226–1245
Naydenov K, Senneville S, Beaulieu J, Tremblay F, Bousquet J (2007) Glacial vicariance in Eurasia: mitochondria DNA evidence from Scots pine for a complex heritage involving genetically distinct refugia at mid northern latitudes and in Asia Minor. BMC Evol Biol 7(233):1–12
Pardos JA, Lange W, Weißmann G (1990) Morphological and chemical aspects of Pinus sylvestris L. from Spain. Holzforschung 44(2):143–146
Peakall R, Smouse PE (2006) GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6:288–295
Petit RJ, Aguinagalde I, de Beaulieu JL, Bittkau C, Brewer S, Cheddadi R, Ennos R, Fineschi S, Grivet D, Lascoux M, Mohanty A, Müller-Starck G, Demesure-Musch B, Palmé A, Martín JP, Rendell S, Vendramin GG (2003) Glacial refugia: hotspots but not melting pots of genetic diversity. Science 300(5625):1563–1565
Prus-Głowacki W, Stephan BR (1994) Genetic variation of Pinus sylvestris from Spain in relation to other European populations. Silvae Genet 43:7–14
Prus-Głowacki W, Stephan BR, Bujas E, Alia R, Marciniak A (2003) Genetic differentiation of autochthonous populations of Pinus sylvestris (Pinaceae) from the Iberian Peninsula. Pl Syst Evol 239:55–66
Prus-Głowacki W, Urbaniak L, Bujas E, Curtu AL (2012) Genetic variation of isolated and peripheral populations of Pinus sylvestris L. from glacial refugia. Flora 207:150–158
Pyhäjärvi T, García-Gil MR, Knürr T, Mikkonen M, Wachowiak W, Savolinem O (2007) Demographic history has influenced nucleotide diversity in European Pinus sylvestris populations. Genetics 177:1713–1724
Pyhäjärvi T, Salmela MJ, Savolinem O (2008) Colonization routes of Pinus sylvestris inferred from distribution of mitochondrial DNA variation. Tree Genet Genome 4:247–254
Robledo-Arnuncio JJ, Collada C, Aliá R, Gil L (2005) Genetic structure of montane isolates of Pinus sylvestris L. in a Mediterranean refugial area. J Biogeogr 32:595–605
Rubiales JM, García-Amorena I, Hernández L, Génova M, Martínez F, Gómez Manzaneque F, Morla C (2010) Late Quaternary dynamics of pinewoods in the Iberian Mountains. Rev Palaeobot Palynol 162:476–491
Scalfi M, Piotti A, Rossi M, Piovani P (2009) Genetic variability of Italian southern Scots pine (Pinus sylvestris L.) populations: the rear edge of the range. Eur J For Res 128:377–386
Semiz G, Heijari J, Isik K, Holopainen JK (2007) Variation in needles terpenoids among Pinus sylvestris L. (Pinaceae) provenances from Turkey. Bioch Syst Ecol 35:652–661
Sinclair WT, Morman JD, Ennos RA (1999) The postglacial history of Scots pine (Pinus sylvestris L.) in Western Europe: evidence from mitochondrial DNA variation. Mol Ecol 8:83–88
Sokal RR, Rohlf FJ (2003) Biometry. The principles and practice of statistics in biological research, 3rd edn. Freeman and Co., New York
Soranzo N, Alia R, Provan J, Powell W (2000) Patterns of variation at a mitochondrial sequence-tagged-site locus provides new insights into the postglacial history of European Pinus sylvestris populations. Mol Ecol 9:1205–1211
Soto A, Robledo-Arnuncio JJ, González-Martínez SC, Smouse PE, Alía R (2010) Climatic niche and neutral genetic diversity of the six Iberian pine species: a retrospective and prospective view. Mol Ecol 19:1346–1409
Staszkiewicz J (1961) Variation in recent and fossil cones of Pinus sylvestris L. Fragm Flor Geobot 7:97–160
Staszkiewicz J (1963) Recherches biométriques sur la variabilité des cônes du Pin sylvestre (Pinus sylvestris L.) du Massif central en France. Fragm Flor Geobot 9:175–187
Staszkiewicz J (1968) Investigations on Pinus sylvestris L. from South-eastern Europe and from Caucasus and its relation to the pine from other territories of Europe based on morphological variability of cones. Fragm Flor Geobot 14:259–315 [in Polish with English summary]
Staszkiewicz J (1993) Pine in the history of the forests. From Tertiary to Holocene. In: Białobok S, Boratyński A, Bugała W (eds) Biology of Scots pine. Sorus, Poznań – Kórnik, pp 11–17
Svoboda P (1953) Forest trees and its’ stands. Státní Zemĕdĕlskĕ Nakladatelství, Praha [in Slovak]
Tobolski JJ, Hanover JW (1971) Genetic variation in the monoterpenes of Scotch pine. For Sci 17:293–299
Turna I (2003) Variation of some morphological and electrophoretic characters of 11 populations of Scots pine in Turkey. Israel J Pl Sci 51:223–230
Turna I, Güney D (2009) Altitudinal variation of some morphological characters of Scots pine (Pinus sylvestris L.) in Turkey. Afr J Biotechnol 8:202–208
Watała C (2002) Biostatistics—utilisation of the statistic methods in the scientific biological investigations. α-medica Press, Bielsko-Biała [In Polish]
Willis KJ, van Andel TH (2004) Trees or no trees? The environments of central and eastern Europe during the Last Glaciation. Quat Sci Rev 23:2369–2387
Willis KJ, Bennet KD, Birks JB (1998) The late Quaternary dynamics of Pine in Europe. In: Richardson DM (ed) Ecology and biogeography of Pinus. Cambridge University Press, Cambridge
Yena A, Yena A, Yena V (2005) ‘Stankiewicz pine’ in Crimea: some new taxonomical, chorological and paleo-landscape considerations. Dendrobiol 53:63–69
Zar JH (1999) Biostatistical analysis. Prentice-Hall, New Jersey
Acknowledgments
The research was financially supported by The Polish Ministry of Science (Contr. No. NN303 360535) and partly by Institute of Dendrology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Jasińska, A.K., Boratyńska, K., Dering, M. et al. Distance between south-European and south-west Asiatic refugial areas involved morphological differentiation: Pinus sylvestris case study. Plant Syst Evol 300, 1487–1502 (2014). https://doi.org/10.1007/s00606-013-0976-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00606-013-0976-6