
Abstract
Desert ants, Cataglyphis fortis, perform large foraging excursions during which they continuously compute a home vector that allows them to return to the nest on the shortest way. This type of navigation, termed path integration, needs a compass system and an odometer. Ants use several cues to determine their walking direction, two of the most important ones being the sun position and the polarization pattern of the sky. We tested whether an information transfer is possible from one compass system to the other, which depend on different anatomical substrates. Since the sky’s polarization pattern is detected by UV-photoreceptors located in the dorsal rim area (DRA), we used an orange Perspex filter that eliminated the UV part of the spectrum to prevent the use of the polarization compass. The use of the sun compass could be excluded by appropriate screens. In the critical tests the ants had learned a nest-feeder direction with e.g. the sun compass only, and were later tested with the polarization compass, or vice versa. The results show that a transfer is possible in both directions.
Similar content being viewed by others
Introduction
Desert ants of the genus Cataglyphis are champions of path integration (Müller and Wehner 1988; Wehner and Srinivasan 2003; Ronacher 2008). This kind of navigation implies a vector computation of the distances travelled in certain directions, i.e., an integration of directional information and of the corresponding actual distance information. Cataglyphis ants infer the travelling distances by means of a step integrator (Wittlinger et al. 2006). The necessary directional information can be derived from various celestial and terrestrial cues: the sky’s polarization [=POL] pattern, the sun’s azimuth, the spectral composition and intensity distribution of skylight (Wehner and Rossel 1985; Fent 1986; Wehner (1994, 1997); Wehner and Labhart 2006), wind (Wehner and Duelli 1971; see also Wolf and Wehner 2000; Müller and Wehner 2007), and panoramic scenes including landmarks (Collett et al. 2001; Wehner 2003; Graham and Cheng 2009; Wystrach et al. 2011; Baddeley et al. 2012; Collett et al. 2013). Under certain conditions wind or the panorama can influence path directions when experimentally disconnected from the celestial cues (Müller and Wehner 2007; Graham and Cheng 2009). However, the two celestial compass systems––POL and sun compass––are the most relevant ones to infer the direction of a path segment (Wehner and Duelli 1971; Ronacher et al. 2006; Müller and Wehner 2007; Heß et al. 2009; Lebhardt et al. 2012). Therefore, in the present account we focus on these.
The sky’s polarization pattern is detected by specialized ommatidia located in the dorsal rim area (DRA) and also by the ocelli (Wehner 1982; Fent and Wehner 1985). The ommatidia of the DRA house only a single photoreceptor type, which in Cataglyphis is sensitive in the UV part of the spectrum (Duelli and Wehner 1973; Wehner (1989, 1994); Labhart 2000; Wehner and Labhart 2006). Under most conditions, the sun’s position is perceived by different parts of the eye. Thus, in the periphery both compass systems rely on different anatomical substrates (Wehner 1997).
Under natural conditions, the POL pattern is tightly linked to the sun’s position (Wehner 1997). In experiments that precluded the use of one of the two celestial compass systems, Cataglyphis ants nevertheless showed well-oriented homebound paths (Duelli and Wehner 1973; Wehner and Müller 2006; Lebhardt et al. 2012). Thus, sun and POL compass, while acting together under natural conditions, may each be sufficient for navigation in the absence of the other cue. If ants are deprived from using both sun and POL compass they may still infer their homing direction, though less accurately, from the sky’s spectral gradient (Wehner 1997).
Remarkably, interocular transfer is possible for the celestial compass cues: ants trained to a feeder bearing a monocular eye cap found their way home when the eye cap was transferred to the other eye (Wehner and Müller 1985). It is, however, still unknown if a transfer is possible from one compass system to the other: if a compass direction was learned exclusively with the POL compass, can the ant recall this direction on the basis of its sun compass alone––or vice versa? In a comprehensive account on the ant’s polarization and spectral compass, Wehner (1997) tackled this question and by presenting some first data concluded that such an inter-compass transfer is possible. However, as this hypothesis was based on a rather small sample size, we present here an extensive study, in which we apply a new approach using a linear POL filter that allowed us to manipulate the POL compass cues independent of the sun’s position (Lebhardt et al. 2012). To exclude the POL cues we used orange-coloured Perspex that did not allow UV light to pass through, thus silencing the POL compass (Wehner 1982, 1994). Our data provide clear evidence that directional information obtained via the POL compass can be recalled by the sun compass, and vice versa.
Materials and methods
General training and test procedure
Desert ants of the species Cataglyphis fortis (Forel 1902) were studied in a dry salt pan near the village Menzel Chaker (34°57′N, 10°24′E) in Tunisia during June and July 2013. This area is devoid of prominent landmarks and, therefore, provides optimal conditions to study navigation strategies based on the celestial cues.
The ants were trained to walk in U-shaped aluminium channels (7 cm width and height) from their nest to a 7.5 m distant feeding station containing biscuit crumbs (for details see Grah et al. 2005; Lebhardt et al. 2012). Earlier observations had shown that ants rely on their last excursion to determine the homing direction by path integration (Wehner et al. 2002). Therefore, we did not record how often individual ants visited the feeder, but started the testing after a continuous flow of ants shuttling between the feeder and the nest was established and most likely the majority of the ants had visited the feeder at least more than once before being tested. While running between the nest and the feeder, the ants were deprived from using either the POL compass or the sun compass (except for a distance of ~15 cm between the nest exit and the channel entrance). Individual ants were then captured at the feeder and released on a distant test field to perform their homebound run under a trolley that excluded the compass system that was available during training. The trolley was constructed following Duelli and Wehner (1973) and Fent (1986). It had a circular opening that allowed a view of the sky in a ~130° window (see Fig. 2a in Fent 1986). The opening was covered either with orange Perspex (to exclude the use of the POL compass) or with UV-transparent Perspex that allowed a view of the celestial POL pattern (see “Materials and methods”); the direct view of the sun could be prevented by a movable circular screen (diameter 28–33°). Around the trolley a dark curtain was fixed that shielded the ant from wind (for further details see Wehner 1982; Fent 1986; Wehner 1994). On the test field a grid with mesh width of 1 m was painted with white colour that enabled the observer to record the ant’s path. The trolley was moved by an assistant so that the ant was always in the centre of the circular opening.
Paradigm I: Training with sun compass information while excluding the POL compass cue
The POL compass can be knocked out by removing wavelengths below 410 nm (Duelli and Wehner 1973). We trained ants under an orange Perspex sheet attached above the training channel that filtered all wavelengths below 530 nm (Plexiglas® GS 2C04, Evonik Industries AG, Darmstadt—transmission is less than 0.01 % for wavelengths below 506 nm (Wehner and Müller 2006), see transmission curve e, in Fig. 43 in Wehner 1982). This filter also prevents the perception of spectral gradients, since only the long-wavelength photoreceptor is still functional. During the training, a nest-to-feeder orientation of the channel of 120° in the morning (AM) and of 240° in the afternoon (PM) was chosen to ensure that the ants had a direct view of the sun when shuttling in the training channel (Fig. 1a, 0° corresponds to north). In the critical test, the ants performed their homebound run under the experimental trolley that was equipped with a UV light-transmitting Perspex allowing view of the sky’s natural polarization pattern [Plexiglas® GS 2458, Evonik Industries AG, Darmstadt; the transmission was above 90 % down to 300 nm, see Online Resource 1 (Fig. S1)]. In these tests, the direct view of the sun was prevented by the movable screen. Different cohorts of animals performed two controls after the same type of training (see Table 1). In the first control animals could perform their homebound run on the test field with free sight of the sky. In the second control, the animals were tested under the trolley with only the sun compass available, i.e. in the same condition as during the training.

Scheme of the training channels. a Experimental paradigm I (training without POL information). Channels were aligned differently in the morning (AM) and afternoon (PM) experiments to allow a view of the sun, such that nest directions were at 300° and 60° respectively. b In the paradigms II and III a linear POL filter reduced the available POL information to a single e-vector direction. The nest directions were at 45° and 315°, respectively, and direct view of the sun was prevented by a 50 cm high screen located on the right or left side of the channel (continuous lines)
Paradigm II and III: Excluding the sun compass cue while providing POL compass information during training
During the training the direct view of the sun was prevented by thin wooden plates (50 cm height) erected along one side of the training channel. The best shadows were obtained when the channel was oriented perpendicular to the sun’s position, i.e. towards 225° (AM) and 135° (PM) (Fig. 1b). We further manipulated the polarization pattern visible to the ant by covering the training channels with a polarization filter that is transparent in the visible and the UV part of the spectrum (HN38 Polarisationsfolie linear, 0.3 mm; Fa. ITOS GmbH, Mainz, Germany, for the transmission curve see Heß et al. 2009). This filter provided uniform POL information along the entire training distance, consisting of a 60° broad overhead stripe with a single e-vector orientation (in the present account either orthogonal or parallel to the ant’s walking direction). Such POL compass information allows a good prediction of the respective homing directions when ants are later tested under open sky conditions (see Lebhardt et al. 2012). In the critical test the return run was performed under the trolley equipped with orange Perspex and with the sun visible to the ants. For the controls, the trolley was covered with UV-transparent Perspex allowing view of the sky’s natural polarization pattern while the direct sunlight was blocked by the movable screen (see Table 1).
Data evaluation and statistical analysis
We documented the trajectories of the homing ants and determined the homing directions at 2, 3 and 4 m from the release point. The data were then plotted either relative to the actual nest direction in case of the sun compass training or relative to the solar azimuth after training under the POL filter. The results are illustrated in circular diagrams and circular statistics were applied using the software package Oriana2 (Kovach Computing Services, Angelsey, Wales). The Rayleigh test was used to test against a random circular distribution and the One sample t test was used to determine whether the expected homing direction (either sun azimuth or nest direction) belonged to a confidence interval. We applied the Mardia–Watson–Wheeler test to test for differences between two distributions and used the Wilcoxon–Mann–Whitney test to compare the deviation from their respective means of two independent samples (see Batschelet 1981; Zar 1999). Each ant was tested only once.
Results
Recall with the POL compass after sun compass training?
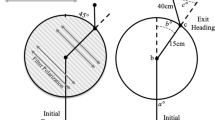
In the training paradigm I (see Table 1), ants were trained to visit a feeder in a channel covered with orange Perspex that eliminated POL cues. To allow view of the sun, the feeder was located at 120° in the morning (viewed from the nest) and at 240° in the afternoon, yielding homing directions of 300° and 60°, respectively (Fig. 1a). In the critical experiment, the ants were tested under a trolley that blocked a direct view of the sun but allowed UV light to pass through so that a large part of the sky’s POL pattern was available for the POL compass. The homing paths were on average oriented towards the nest (0° in Fig. 2a). There were significant deviations between the mean vectors in the morning and afternoon sessions (Mardia–Watson–Wheeler test: p < 0.001 for all distances; in the morning session, the 99 % confidence intervals include 0° for 3 and 4 m, while in the afternoon the confidence intervals do not include the nest direction at all distances; n = 23 and 22 ants tested, respectively). However, the very same small deviations also occurred in a control experiment in which ants were again trained exclusively with the sun compass but then tested without the trolley, that is, with a full view of the sky [POL and sun compass (Fig. 2b); n = 26 (AM) and 25 (PM)]. The Mardia–Watson–Wheeler test indicates a good agreement between the mean vector directions in Fig. 2a, b (differences not significant: p = 0.143–0.26 for the AM tests at 2–4 m distance, and 0.098–0.90 for the PM tests).

Results of paradigm I: training with the sun as the only sky compass cue. a Critical test with POL only. b Control with sun and POL visible (without trolley), c control with only sun visible. 0° indicates nest direction; open and closed symbols correspond to AM and PM tests, respectively. Arrows indicate lengths of the mean vectors [r between 0.75 and 0.97 in a; 0.84–0.98 in b; 0.78–0.92 in c; all vectors indicate highly significant orientation, p << 0.0001 (Rayleigh test)]. Arcs below the diagram indicate the ranges of the sun’s azimuth during AM and PM tests. Samples of individual trajectories in the different test situations can be found in Online Resource 1 (Fig. S2)
Figure 2c shows the results of an additional control: as before the ants were trained with only the sun compass, but in this case the return path was also performed with only sun compass cues available (under the trolley with orange Perspex; see Table 1). In this control the ants showed substantial deviations from the expected nest direction, in particular during the AM tests. However, the homing paths were clearly non-random [Rayleigh test: all p << 0.001, n = 17 (AM) and 19 (PM)], and the ants seemed to head in the direction of the nest rather than towards the sun’s azimuth. The deviations from the nest direction in this control can be explained qualitatively by previous observations that the orange Perspex filter not only abolishes the POL cues but also colour vision since only the long wavelength (“green”) receptors are left functional. In this condition, the ants exhibited a strong phototactic response towards the brightest part of the sky even if the sun itself was occluded (Lanfranconi 1982; Wehner 1997). This positive phototaxis caused substantial deviations from the trained compass direction (up to 38°; pp. 165–169 in Wehner 1997). In the experiment of Fig. 2c the sun was mostly located at the northern side of the training channel during the morning (cf. Fig. 1); i.e. an ant returning to the nest experienced the brightest part of the sky on its right side. Correspondingly, during the test under the trolley the phototactic response would induce a deviation to the north, i.e. to the right side in Fig. 2c. During the afternoon tests the northern side of the channel was again brighter; hence a returning ant experienced a phototactic drive to its left side that competed with the compass response. This effect is also visible in Fig. 2c (filled symbols). In contrast, in the tests of Fig. 2a, b, the returning ants could rely on the celestial POL pattern and in this situation the phototactic response plays no role (Wehner 1997).
When interpreting the results of Fig. 2a, b a caveat has to be mentioned. An essential condition for this experiment was that the ants could see the sun while walking in the narrow training channel. Thus, the channel had to be aligned in a way that the ants walked towards a feeder which was located more or less in the sun’s direction; conversely the nest direction coincided roughly with the antisun meridian. For this reason, we cannot separate the nest direction from the antisun azimuth, and indeed if we normalize the data to the antisun azimuth this yields almost the same accuracy as normalization to the nest direction (lengths of mean vectors 0.771, 0.759, and 0.721 (antisun) versus 0.816, 0.789, and 0.754 (nest) in the morning session of Fig. 2a; and 0.789, 0.831, 0.864 versus 0.842, 0.875, 0.886 for Fig. 2b; for the afternoon the results of both normalizations yielded indistinguishable mean vector lengths). Nonetheless, based on evidence that ants exhibit strong phototaxis in some situations (Duelli and Wehner 1973; Wehner and Müller 1985; Wehner 1997), we consider it rather unlikely that in the training paradigm I the ants had acquired a vector directed 180° away from the sun. On the contrary, in view of the phototaxis effects described above, the opposite effect (i.e., a bias towards the sun’s azimuth) would be expected. Such a bias is clearly absent in Fig. 2a, b. In any case, the well-focused homing paths in Fig. 2a show that the information derived during the training from the sun’s azimuth was transferred and enabled a recall of the trained direction based on the POL compass. Thus, Fig. 2a provides the crucial proof for a transfer from sun compass to POL compass.
Recall with the sun compass after training with the POL compass?
The reciprocal experiment differs from the first in that the training with a single e-vector direction does not allow an ant to pinpoint the nest direction. Instead, the expected homing direction shifts during the day along the sun’s azimuth (Lebhardt et al. 2012). Hence, in these experiments we normalized the homing directions with respect to the sun’s azimuth (0° in Figs. 3, 4). If during training the ant experienced an orthogonal e-vector (Paradigm II), i.e. perpendicular to its walking direction, this corresponds to a direction in the solar or antisolar meridian (Fent 1986; Lebhardt et al. 2012). Figure 3a summarizes the results of the critical test. The ants were trained with the orthogonal POL filter without view of the sun (see Fig. 1b; Table 1). On the test field they performed their homebound walk under the trolley equipped with orange Perspex, thus preventing the use of the POL compass. Although the scatter is relatively large, the ants showed a significant orientation towards the sun’s azimuth (Fig. 3a, Rayleigh test: p = 0.004 to p < 0.0001; the 99 % confidence intervals include 0° for all AM data and for the 4 m PM data). Figure 3b shows the control experiment: the ants were trained in the same orthogonal e-vector paradigm, in which they had no sight of the sun but an almost full view of the celestial POL pattern during their homebound run under the trolley. Again the ants showed very precise orientation, with little scatter; [the mean vector directions are not significantly different between Fig. 3a, b, while the circular standard deviations are more than twice as large in a (mean difference 110 %, significant in four out of six cases, p = 0.009 to p < 0.001; Wilcoxon–Mann–Whitney test for dispersion)]. The data of Fig. 3a demonstrate that the ants can use directional information obtained with the POL compass in a test situation where only the sun compass is functional.

Results of paradigm II: training with an orthogonal e-vector. a Critical test with sun only, b control with celestial POL pattern visible and shielded sun, i.e. compass condition similar to training. 0° indicates the sun’s azimuth; open and closed symbols correspond to AM and PM tests, respectively. The arcs beside the diagrams indicate the nest positions relative to the sun’s azimuth; other conventions as in Fig. 2. Lengths of the mean vectors r = 0.457–0.68 (AM) and r = 0.905–0.928 (PM) in a; r = 0.83–0.98 in b all values highly significant (Rayleigh test: p = 0.004 to p < 0.0001 for the AM data and all p << 0.0001 for the PM data in a, all p << 0.0001 in b)

Results of paradigm III: training with a parallel e-vector. a Critical test with sun only, b control with celestial POL pattern visible and shielded sun. 0° indicates the sun azimuth; open and closed symbols correspond to AM and PM tests, respectively. The arcs beside the diagrams indicate the nest positions relative to the sun azimuth; other conventions as in Fig. 2. The lengths of the mean vectors were determined separately for the left and right halves of the diagrams except for the cases with less than five animals [AM 90° in a and b, PM 270° in b; in a r = 0.79–0.98, in b r = 0.96–0.99; all p < 0.007 (Rayleigh test)]
An analogous experiment was performed with a different orientation of the POL filter (Paradigm III). Now the e-vector direction in the training channel was parallel to the ant’s walking direction. In that situation the ants expect their homing direction to lie perpendicular to the sun-antisun meridian (Fent 1986; Lebhardt et al. 2012). Indeed, in the control experiment (Fig. 4b), the mean vector directions are close to the expected 90° and 270°, though with small but significant deviations towards the sun’s azimuth (0°; AM 270°: p < 0.01 for 2 and 3 m data; 4 m n.s. (n = 11); PM all p < 0.01 for 90° data (n = 17–15), n.s. for 270° data). In the critical test the homing directions deviate more towards the sun’s azimuth (Fig. 4a; the respective distributions are significantly different from the control; Mardia–Watson–Wheeler test: p < 0.001 for the PM and p = 0.055 to p < 0.03 for AM data). However, this result again is most likely due to the phototaxis effect mentioned above. In this paradigm, the ants should move at right angles to the sun meridian, and hence the phototaxis effect towards the brightest part of the sky would pull the ants towards the sun azimuth—which is what we see in Fig. 4a. The mean deviation from the control was 38° and 19° for the AM and PM data, respectively—which is in the same range as observed for the phototaxis effect by Wehner (Figs. 12, 13 in Wehner 1997). Note that in the paradigm with the orthogonal POL filter (Fig. 3) the phototaxis effect would work in the same direction as the compass direction and therefore, was not evident in Fig. 3a.
Discussion
In the experiments presented above the ants had to cope with different combinations of celestial cues they experienced during the respective training and test situations. The data in Figs. 2, 3, and 4 suggest that the ants can transfer directional information from the sun compass to the POL compass system, and vice versa.
Salience of different orientation cues
To substantiate the conclusion that directional information is transferable from one compass system to the other we must rule out other possible orientation cues. First, we consider the diverse cues known to be used by Cataglyphis to infer directional information (see “Introduction”). All terrestrial cues—landmarks, panorama and wind (e.g. Wolf and Wehner 2000; Collett et al. 2001; Müller and Wehner 2007; Bregy et al. 2008; Graham and Cheng 2009; Zeil 2012; Collett et al. 2013)—were excluded to a large degree by the experimental design. The landscape where the experiments were performed offered very few potential landmarks. In addition, during training the view was restricted by the channel system, while during the tests under the trolley, landmarks were obscured by a curtain. During training in the channel the ants experienced some head or tail wind but in the test situation under the trolley they were shielded from wind. Hence, only celestial cues were available to infer walking directions: POL pattern, sun position and spectral or intensity gradients.
During training in paradigms II and III the ants experienced only a modified POL pattern, i.e. an overhead stripe with a single e-vector orientation (either orthogonal or parallel to their walking direction). In a previous experiment, after the same type of training the ants performed well-oriented homing courses when tested with a free view of the sky (Lebhardt et al. 2012). This was confirmed here in the controls (Figs. 3b, 4b); the difference between the two experiments was that in the current design the trolley prevented the direct view of the sun. This type of experiment showed again that the POL (plus spectral) cues are sufficient to provide very good orientation.
The long-wavelength filter induces an additional phototaxis effect
In the sun-only training (paradigm I) and in the sun-only tests (see Table 1) the orange Perspex eliminated not only the POL cues, but also prevented the perception of spectral gradients, since only the long-wavelength receptor type was left functional (Lanfranconi 1982; Wehner and Müller 2006). In this condition, the ants exhibited a strong phototactic response towards the brightest part of the sky even if the sun itself was occluded (Lanfranconi 1982; Wehner 1997). This positive phototaxis caused substantial deviations from the trained compass direction (up to 38°; Figs. 12, 13 in Wehner 1997), and can explain the observed deviations from the expected homing directions in paradigm I (Fig. 2c) and paradigm III (Fig. 4a)—see “Results”. However, since the orange filter affected not only the UV-based POL compass, but also produced an unnatural percept of the sun, this could have influenced the salience of the sun compass. Nonetheless, we found clear evidence that the sun was used as a compass cue, even under the long-wavelength filter. Thus, even when there were restrictions on the available spectral information (or when the spectral information typically associated with the sun was distorted to a high degree), ants were able to orient with the sun compass. This conclusion corresponds with results of an earlier experiment reported by Wehner (Figs. 16, 17 in Wehner 1997), while it is in contrast to Wehner and Müller (2006) who concluded that the natural spectral radiance distribution is essential for the compass.
Although the orange Perspex prevented the use of the POL compass and of spectral gradients, an intensity gradient across the sky still remained as a potential compass cue. Hence, theoretically the ants could have relied on the intensity gradient both under POL filter conditions during training as well as under the orange Perspex during the test, without the necessity of a transfer of directional information between sun and POL compass. This assumption, however, seems highly unlikely for the following reasons. During the AM POL filter training the nest direction was at 45° (Fig. 1b) while the sun, representing the gravity centre of the intensity gradient, wandered from 90° to 130°. The walking direction experienced during training thus was at −45° to −85° relative to the intensity gradient and the sun azimuth (see arcs in Figs. 3a, 4a). Relying on the intensity gradient alone, the ants should have chosen the same homing direction, irrespective of the different POL patterns experienced during training II and III. This was clearly not the case (compare Figs. 3a, 4a). In particular after orthogonal POL filter training, the homing paths of ants should deviate about 45°–85° from the sun azimuth, which is contradicted by the data of Fig. 3a. Finally, navigation based on the intensity gradient should have led to a much larger scatter of homing directions (Wehner 1997) than that observed in our experiments (see Figs. 2a and 3a (PM data) and Fig. 4a).
The sky’s polarization pattern as well as the pattern provided by a POL filter are symmetric and therefore, yield a 180° ambiguity resulting in a bimodal distribution of heading directions. Interestingly, this bimodality could be observed after training with the parallel oriented POL filter, but was absent for the orthogonal POL filter training (only one ant headed towards the antisolar direction) or the sun-only training which was followed by the test with only the sky’s polarization pattern available. In earlier studies, this bimodality was also absent in the paradigm with an orthogonal POL filter (Fig. 2a, b in Lebhardt et al. 2012). Thus, it seems that the ants indeed can distinguish between the solar and the antisolar half of the sky (the latter is more polarized) and therefore, the ambiguity of the POL pattern is resolved (Fent 1986). This leads to an unimodal distribution in the sun-only training situation as well as in the orthogonal POL filter training. Only after training under a parallel e-vector orientation relative to the walking direction, the directional information is ambiguous and the ants were not able to distinguish between the two possible homing directions directed perpendicular to the solar meridian.
Dominance of the POL compass?
According to Wehner and Müller (2006) the polarization compass of Cataglyphis completely dominates over the sun compass (Duelli and Wehner 1973; see also Müller and Wehner 2007). However, in a recent experiment, which set the POL compass in conflict with the sun compass, the ants adhered to a compromise direction between sun and POL-based homing directions indicating a roughly equal contribution of the two compass systems (Lebhardt and Ronacher 2013). How can this result be reconciled with the postulated dominance of the POL compass system, and do the present results bear on this question?
The precision of homing directions in the present experiments may give additional hints to the salience of the two compass systems. First, a comparison of Fig. 2a, b shows the impact of eliminating the sun as a compass cue. The circular standard deviations in Fig. 2a were somewhat larger than in Fig. 2b, although the differences were significant in only one out of six comparisons (two-sided F test: p < 0.01). In addition, some of the increased variances may be attributed to the test situation under the trolley—in the control tests with both the sun and POL pattern visible (Fig. 2b) the ants moved freely after release on the test field. Thus, elimination of the sun as a compass cue only marginally deteriorated the precision of orientation via the POL compass, if at all. The standard deviations in Fig. 2c cannot be compared directly to Fig. 2a, because of the influence of phototaxis on the compass direction (cf. “Results”).
A direct comparison of the two compass systems can be made in paradigm II, in which the ants could rely either on the sun (Fig. 3a) or on the sky’s POL pattern (Fig. 3b) for their homing path. The circular standard deviations were on average more than twice as large if the homing was performed under the sun condition (see “Results”). Although the significant increase in the variances under the sun condition may in part be attributable to the switch from one compass system to the other, the strength of the effect indicates that the sun is an inferior cue compared to the celestial POL pattern. Furthermore, removing the spectral information by the orange cut-off filter may have caused a further imbalance against the sun compass and in favour of the POL pathway. The standard deviations for the transfer experiments from POL to sun compass (Fig. 3a) were on average 57 % larger than those for the transfer experiments from sun to POL compass (Fig. 2a; the differences were significant in three out of six cases, one sided F test: p < 0.01, < 0.005, < 0.001). Taken together, these comparisons suggest a stronger weight for the POL compass. This conclusion is further supported by the observation that the phototaxis effect appeared to have an effect only when the sun compass was used and not in combination with the POL compass (compare Fig. 2a, c; see also Wehner 1997).
Conclusion: a common final stage of compass direction processing
The results of Figs. 2, 3, and 4 show that directional information obtained from the sun compass can be used to retrace a home vector, if only the POL compass is available, and vice versa. Hence, we conclude that in the ant’s brain an anatomical structure exists, before the motor output, into which both types of information—arriving via anatomically distinct pathways—are fed and can substitute each other. It is, however, not possible solely based on behavioral experiments to determine at which stage exactly a transfer or substitution of directional information takes place. Conceivably, the directional information obtained by either the POL or the sun compass could be forwarded via two separate pathways until they reach a common ‘path integration module’ where they may substitute each other for the computation of the ‘home vector’ without requiring any prior transfer of information between the two pathways. Alternatively, the information of two different compass systems may already be consolidated at an earlier, more peripheral stage, and then be further processed as a combined message in downstream centers. Indeed in locusts and monarch butterflies several neurons have been recorded at different stages of the polarization vision pathway that could play such an integrative role (the respective ganglia range from the optic lobe over the anterior optic tubercle to the central complex; Pfeiffer and Homberg 2007; Heinze and Reppert 2011; el Jundi et al. 2011; reviewed in el Jundi et al. 2014). If both pathways feed into a common centre, an exchange between sun and POL compasses as well as the resulting compromise directions in conflict experiments (Lebhardt and Ronacher 2013) are easily explained. During training and test in the present experiments an ant had access to different reference signals, which are transmitted via two different anatomical substrates. The results of our experiments provides an evidence that the compass information obtained by one system can be successfully recalled by the other, and that at some stage a transfer of directional information occurs.
We hypothesize that under normal conditions the POL compass may indeed be dominant since a large number of DRA ommatidia are exclusively devoted to the processing of POL information. In addition, the ocelli contribute to this pathway (Fent and Wehner 1985; Schwarz et al. 2011). In contrast, the sun is perceived by only one or a few ommatidia. Conceivably, the determination of an intensity or spectral gradient across the sky may yield smaller excitation values in downstream neurons because the ommatidia outside the DRA are not specifically tailored to such a task. In the cue conflict experiments of Lebhardt and Ronacher (2013) only a single e-vector direction was visible during training. Hence, fewer DRA ommatidia were optimally excited than would have been if the complete celestial POL pattern was visible, which may have reduced the strength of the POL input and favoured homing in a compromise direction. In the experiments yielding a dominance of the POL compass the ants could see the celestial POL pattern in a 90° stripe and thus, experienced a broad range of e-vector directions (Wehner and Müller 2006). This may have increased the impact of the POL compass relative to the sun compass.
We propose an integration stage that combines the input from the two compass pathways in a weighted fashion, with the weights depending on how many ommatidia are involved, and how strongly these are stimulated. By partial covering of the DRA it should be possible to reduce the input strength of the POL compass. Furthermore, we predict a difference in homing precision between experiments performed under the orange filter and experiments in which only the DRA and ocelli are covered: in the latter, the sun should have a stronger influence compared to the former because the filter eliminates spectral cues. Such experiments are presently under way.
References
Baddeley B, Graham P, Husbands P, Philippides A (2012) A model of ant route navigation driven by scene familiarity. PLoS Comput Biol 8:e1002336. doi:10.1371/journal.pcbi.1002336
Batschelet E (1981) Circular Statistics in Biology. Academic Press, London
Bregy P, Sommer S, Wehner R (2008) Nest-mark orientation versus vector navigation in desert ants. J Exp Biol 211:1868–1873. doi:10.1242/jeb.018036
Collett TS, Collett M, Wehner R (2001) The guidance of desert ants by extended landmarks. J Exp Biol 204:1635–1639
Collett M, Chittka L, Collett TS (2013) Spatial memory in insect navigation. Curr Biol 23:789–800. doi:10.1016/j.cub.2013.07.020
Duelli P, Wehner R (1973) The spectral sensitivity of polarized light orientation in Cataglyphis bicolor (Formicidae, Hymenoptera). J Comp Physiol A 86:37–53. doi:10.1007/BF00694476
el Jundi B, Pfeiffer K, Homberg U (2011) A distinct layer of the medulla integrates sky compass signals in the brain of an insect. PLoS One 6:e27855. doi:10.1371/journal.pone.0027855
el Jundi B, Pfeiffer K, Heinze S, Homberg U (2014) Integration of polarization and chromatic cues in the insect sky compass. J Comp Physiol A 200:575–589. doi:10.1007/s00359-014-0890-6
Fent K (1986) Polarized skylight orientation in the desert ant Cataglyphis. J Comp Physiol A 158:145–150
Fent K, Wehner R (1985) Ocelli: a celestial compass in the desert ant Cataglyphis. Science 228:192–194. doi:10.1126/science.228.4696.192
Grah G, Wehner R, Ronacher B (2005) Path integration in a three-dimensional maze: ground distance estimation keeps desert ants Cataglyphis fortis on course. J Exp Biol 208:4005–4011. doi:10.1242/jeb.01873
Graham P, Cheng K (2009) Ants use the panoramic skyline as a visual cue during navigation. Curr Biol 19:935–937. doi:10.1016/j.cub.2009.08.015
Heinze S, Reppert SM (2011) Sun compass integration of skylight cues in migratory monarch butterflies. Neuron 69:345–358. doi:10.1016/j.neuron.2010.12.025
Heß D, Koch J, Ronacher B (2009) Desert ants do not rely on sky compass information for the perception of inclined path segments. J Exp Biol 212:1528–1534. doi:10.1242/jeb.027961
Labhart T (2000) Polarization-sensitive interneurons in the optic lobe of the desert ant Cataglyphis bicolor. Naturwissenschaften 87:133–136. doi:10.1007/s001140050691
Lanfranconi BC (1982) Kompassorientierung nach dem rotierenden Himmelsmuster bei der Wüstenameisen Cataglyphis bicolor. Dissertation, Univerität Zürich
Lebhardt F, Ronacher B (2013) Interactions of the polarization and the sun compass in path integration of desert ants. J Comp Physiol A. doi:10.1007/s00359-013-0871-1
Lebhardt F, Koch J, Ronacher B (2012) The polarization compass dominates over idiothetic cues in path integration of desert ants. J Exp Biol 215:526–535. doi:10.1242/jeb.060475
Müller M, Wehner R (1988) Path integration in desert ants, Cataglyphis fortis. Proc Natl Acad Sci USA 85:5287–5290
Müller M, Wehner R (2007) Wind and sky as compass cues in desert ant navigation. Naturwissenschaften 94:589–594. doi:10.1007/s00114-007-0232-4
Pfeiffer K, Homberg U (2007) Coding of azimuthal directions via time-compensated combination of celestial compass cues. Curr Biol 17:960–965. doi:10.1016/j.cub.2007.04.059
Ronacher B (2008) Path integration as the basic navigation mechanism of the desert ant Cataglyphis fortis (Forel, 1902) (Hymenoptera: Formicidae). Myrmecol News 11:53–62
Ronacher B, Westwig E, Wehner R (2006) Integrating two-dimensional paths: do desert ants process distance information in the absence of celestial compass cues? J Exp Biol 209:3301–3308. doi:10.1242/jeb.02370
Schwarz S, Albert L, Wystrach A, Cheng K (2011) Ocelli contribute to the encoding of celestial compass information in the Australian desert ant Melophorus bagoti. J Exp Biol 214:901–906. doi:10.1242/jeb.049262
Wehner R (1982) Himmelsnavigation bei Insekten. Neurophysiologie und Verhalten. Neujahrsbl Naturforsch Ges Zürich 184:1–132
Wehner R (1989) Neurobiology of polarization vision. Trends Neurosci 12:353–359. doi:10.1016/0166-2236(89)90043-X
Wehner R (1994) The polarization-vision project: championing organismic biology. In: Schildberger K, Elsner N (eds) Neural basis of behavioural adaptations. Fischer G, Stuttgart, pp 103–143
Wehner R (1997) The ant’s celestial compass system: spectral and polarization channels. In: Lehrer M (ed) Orientation and Communication in Arthropods. Birkhäuser, Basel, pp 145–185
Wehner R (2003) Desert ant navigation: how miniature brains solve complex tasks. J Comp Physiol A 189:579–588. doi:10.1007/s00359-003-0431-1
Wehner R, Duelli P (1971) Spatial orientation of desert ants, Cataglyphis bicolor, before sunrise and after sunset. Experientia 27:1364–1366. doi:10.1007/BF02136743
Wehner R, Labhart T (2006) Polarization vision. In: Warrant E, Nilsson DE (eds) Invertebrate Vision. Cambridge University Press, Cambridge, pp 291–347
Wehner R, Müller M (1985) Does interocular transfer occur in visual navigation by ants? Nature 315:228–229. doi:10.1038/315228a0
Wehner R, Müller M (2006) The significance of direct sunlight and polarized skylight in the ant’s celestial system of navigation. Proc Natl Acad Sci USA 103:12575–12579. doi:10.1073/pnas.0604430103
Wehner R, Rossel S (1985) The bee’s celestial compass: a case study in behavioral neurobiology. Fortschr Zool 31:11–53
Wehner R, Srinivasan MV (2003) Path integration in insects. In: Jeffery KJ (ed) The neurobiology of spatial behaviour. Oxford University Press, Oxford, pp 9–30
Wehner R, Gallizzi K, Frei C, Vesely M (2002) Calibration processes in desert ant navigation: vector courses and systematic search. J Comp Physiol A 188:683–693. doi:10.1007/s00359-002-0340-8
Wittlinger M, Wehner R, Wolf H (2006) The ant odometer: stepping on stilts and stumps. Science 312:1965–1967. doi:10.1126/science.1126912
Wolf H, Wehner R (2000) Pinpointing food sources: olfactory and anemotactic orientation in desert ants, Cataglyphis fortis. J Exp Biol 203:857–868
Wystrach A, Beugnon G, Cheng K (2011) Landmarks or panoramas: what do navigating ants attend to for guidance? Front Zool 8:21. doi:10.1186/1742-9994-8-21
Zar JH (1999) Biostatistical Analysis. Prentice Hall, New Jersey
Zeil J (2012) Visual homing: an insect perspective. Curr Opin Neurobiol 22:285–293. doi:10.1016/j.conb.2011.12.008
Acknowledgments
We thank Julia Stephan and Joshua Alcaniz for their help on the field and the anonymous referees for useful suggestions. Special thanks are due to Rüdiger Wehner for his help with the interpretation of the present data and many discussions over the last years. We also want to express our gratitude to the Tunisian government for kindly allowing us to carry out the investigations in this beautiful country. Financial support was provided by grants from the Deutsche Forschungsgemeinschaft DFG (Ro 547/10-1) and the Volkswagen Foundation (I/78 574) to B.R. The experiments comply with the “Principles of animal Care” and with the current German law.
Conflict of interest
The authors declare that they have no conflict of interests.
Author information
Authors and Affiliations
Corresponding author
Additional information
We dedicate this paper to Rüdiger Wehner who initiated research on desert ant navigation and helped it to blossom by infecting so many students and collaborators with his love for the amazing Cataglyphis ants. Rüdiger Wehner truly deserves the title Tabib-en-Neml—the ant doctor—a nickname given to Felix Santschi 100 years ago.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Lebhardt, F., Ronacher, B. Transfer of directional information between the polarization compass and the sun compass in desert ants. J Comp Physiol A 201, 599–608 (2015). https://doi.org/10.1007/s00359-014-0928-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-014-0928-9