
Abstract
An archaeological excavation in the Tungelroyse Beek valley revealed the remains of two red deer specimens (Cervus elaphus) of Early Mesolithic age that possibly were the victims of hunter-gatherers. The find of animal remains of this age is unique in the Netherlands. In this respect, a sediment core taken close to the remains was investigated, i.e. to reconstruct the vegetation and landscape development of the site and to find more evidence for human activity at this site during the Early Mesolithic. The sediment core shows a typical Early Holocene palynological sequence from the Younger Dryas into the Middle Atlantic, which is supported by AMS dating. The microscopic charcoal record shows peaks in fire activity during the Younger Dryas and Friesland phase, probably wildfire related. Records of spores of coprophilous fungi indicate that the Tungeroyse Beek valley was a favourable place for large herbivores (game) to visit during the investigated period. However, around the age of the oldest red deer remains, no significant peak in fire activity or spores of coprophilous fungi is visible in the investigated record. The pollen diagram does not show disturbed or open vegetation around this age either. This study therefore suggests the impact of Early Mesolithic people on their environment was very low.
Similar content being viewed by others

Introduction
The Mesolithic (Middle Stone Age) is a cultural period that started at the beginning of the Holocene (~10,700 cal bp or 9500 bc) and is characterized by hunter-gatherer, pre-farming societies (Price 1987). The invention or introduction of new technologies, agriculture and the domestication of animals marks the end of the Mesolithic and the beginning of the Neolithic. In Europe, it occurred roughly around 7000 bc (Whittle 1996). In the Netherlands, and in particular in the southern part of the Netherlands, agriculture and animal domestication started around 5300 bc (Louwe Kooijmans et al. 2005).
During the Mesolithic, settlements were generally larger, of longer duration, and more differentiated than in the preceding Palaeolithic (Price 1987). Still, the activities of Mesolithic people had a supposedly small-localized impact on the vegetation. Their impact is, therefore, often poorly reflected in the pollen deposition and easily overlooked in pollen diagrams (Bos and Janssen 1996). Organic remains like wood, bone and antlers, are often not preserved because of the generally dryer conditions during the Boreal. In addition, many sites reached the final stages of sediment accumulation and hydroseral succession during this period. Not surprisingly, little is known about the impact of Mesolithic people on their environment in the Netherlands. There are a few cases described in literature (Bos et al. 2005; de Moor et al. 2009; van der Linden 2011; Bos and Verbruggen 2012; Woldring et al. 2012; Hamburg et al. 2012; Moree and Sier 2014).
Some late Palaeolithic and Mesolithic sites in northwestern Europe were investigated specifically to find evidence for the impact of humans on their environment (e.g. Bos and Janssen 1996; Mellars and Dark 1998; Innes and Simmons 2000; Bos and Urz 2003; Bos et al. 2005). Perhaps the most convincing case for human disturbance in the Mesolithic is recorded in two pollen diagrams near a Mesolithic site in Cambridgeshire (Smith et al. 1989), showing distinct rises in the pollen values of mainly Poaceae, Asteraceae and Galium-type pollen associated with diminished values of arboreal pollen. At some Mesolithic hunting sites, charcoal evidence in addition to palynological evidence was found for human impact on forest vegetation (Mellars and Dark 1998; Bos and Urz 2003; Bos et al. 2005).
In addition, there are a number of palynological studies from the Mesolithic period in northwest and central Europe that show evidence of woodland disturbance without any archaeological evidence for Mesolithic settlements in the direct surroundings (within 1 km) of these sites (e.g. Smith et al. 1989; Simmons 1993). Direct evidence, such as Mesolithic settlement sites and human involvement in the cycle of disturbance and regeneration, is therefore often absent or not conclusive (sensu e.g. Edwards 1989).
In the Tungelroyse Beek valley near Mildert (province of Limburg, the Netherlands) the remains of several larger mammals including two red deer specimens (Cervus elaphus) were discovered in 2010 by RAAP Archeologisch Adviesbureau (Roymans and de Nutte 2012; Roymans 2013) during an excavation at this site. Subsequently the National Heritage Agency of the Netherlands investigated the profiles, made some additional corings and took samples (de Kort 2010). The red deer remains were dated to 6,570 ± 40 bp and 9,650 ± 50 bp (ca. 5500 bc and ca. 9200–8900 bc), respectively. Figure 1 shows the retrieved remains of the red deer dated to 9,650 ± 50 bp. Non-dated flint tools of supposed Mesolithic age were found at the site as well, including a flint stone blade only 6 m southeast of the site that may have been used for butchering (Roymans and de Nutte 2012; Roymans 2013). Although no cut marks were found on the dated red deer remains, cut marks were present on non-dated remains of other bones found at the site, indicating Early Mesolithic human activity (Roymans and de Nutte 2012; Roymans 2013). In addition, the red deer skeletons are far from complete, which also suggests hunting activity (Roymans and de Nutte 2012; Roymans 2013). Animal remains, amongst others red deer remains, with a comparable age have only been investigated by archaeologists in the Netherlands in the valley of the Ooijerhoekse Laak near Zutphen in Gelderland (Groenewoudt et al. 2001; Bos et al. 2005). This makes the find of the red deer remains in the Tungelroyse Beek valley quite unique. The Tungelroyse Beek site has been nominated for national protection (Smit et al. 2013).

In order to gather more information on the palaeo-environment of supposedly Early Mesolithic people in the Tungelroyse Beek area, a sediment core from this area was investigated. The core was collected by the National Heritage Agency of the Netherlands about 5 m from where the remains of the two red deer specimens were found. The research included determining magnetic susceptibility and loss on ignition as lithological proxies, combined with palynological investigation of the core. In addition, the possible impact of Early Mesolithic man on the environment of the site was investigated using microscopic charcoal concentrations in order to link vegetation changes in the vicinity of the investigated site to human impact.
Materials and methods
Geographic location
The investigated archaeological site is located near Mildert in the Dutch province of Limburg, in the municipality of Nederweert (Fig. 2). The site is located in the stream valley of the Tungelroyse Beek, a tributary of the River Meuse. The Tungelroyse Beek is a rain-fed and seepage-fed stream, and its source lies in Belgium. The catchment of the Tungelroyse Beek is bordered in the west by the Dutch–Belgian border, in the south by the Haelensebeek catchment and in the east by the river Meuse. The riverbed is bordered by terraces of Weichselian age of the River Meuse and River Roer, and terraces of the river Meuse of earlier Pleistocene age. These terraces are composed of horizontally layered fine to coarse grained sand beds. In the area, aeolian cover sands of Weichselian age are present as well. These sands locally developed to dunes, but can also form a thin layer, positioned on top of older river sands and younger Meuse terraces. In the area of terraces and aeolian deposits, many local small brook deposits are present comprising both fluvial and lacustrine deposits, which occur alongside the Tungelroyse Beek sediments (Rijks Geologische Dienst 1933).

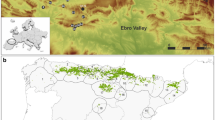
Location of the archaeological site in the Tungelroyse Beek valley near Mildert (the Netherlands). a Location the Netherlands. b Detailed location of the site. Coordinates in Netherlands coordinate system (or Rijksdriehoeksnet). Map provided by Actueel Hoogtebestand Nederland (AHN)
The darker colours showing elevation in Fig. 2b indicate the width of the Tungelroyse Beek valley. As can be derived from Fig. 2b, the investigated site is situated at a location where the valley is relatively narrow. This suggests that in the past, the Tungelroyse Beek may have been easiest to cross at this location for both people and animals, thereby functioning as a corridor (Roymans and de Nutte 2012; Roymans 2013). Today, the Tungelroyse Beek is channelled, which prevents river water reaching the investigated site. The subsurface of the site is classified by the Rijks Geologische Dienst (1933) as sandy peaty soil, and the soil at the archaeological site is relatively wet and organic. The site is comparable to a site near Haelen, investigated by Bos et al. (2007), which is located in the vicinity of the Tungelroyse Beek site.
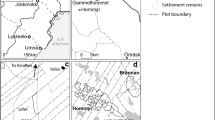
Two pits were made at the archaeological site in the Tungelroyse Beek valley (Roymans 2013). Pit 1 is located where the flint remains of Mesolithic age were found. Pit 2 is located where the red deer bones were found. The two pits are ~150 m apart (Fig. 3). The profiles of Pit 1 and Pit 2 are shown in Figs. 4 and 5, respectively. In addition in Fig. 5 the location of the investigated core is indicated.

Location of excavation pits at the Tungelroyse Beek site. Modified after de Kort (2010). Coordinates in Netherlands coordinate system (or Rijksdriehoeksnet)

Profile of excavation Pit 1, plane 102. Modified after de Kort (2010)

Profile of excavation Pit 2, plane 104 including location of investigated core. Modified after de Kort (2010)
Lithology
The investigated sediment core was taken close to Pit 2 (Figs. 3, 5). The core was collected with an open 6 cm diameter gouge in two segments obtained from a single borehole, resulting in two subsequent cores. The top of the first core is located at 26.05 m +Amsterdam Ordnance Datum (AOD), the top of the second core is located at 25.05 m +AOD. In between the cores, there is a gap of about 5 cm. The lithology of the composite core is presented in Table 1.
Radiocarbon dating
Based on lithology, four samples were taken from the core for AMS radiocarbon (14C) analysis to provide a chronostratigraphical framework for the data (Table 2). Organic material considered to reflect atmospheric 14C conditions, such as terrestrial plants, was preferred, in order to reduce the likelihood of contamination by older or younger carbon (Törnqvist et al. 1992). AMS dating was carried out at the Centrum voor Isotopen Onderzoek (CIO) in Groningen, the Netherlands. Radiocarbon dates are expressed in bp (uncalibrated years before present). Radiocarbon dates were converted to calendar years using the CALIB 7.1 computer program and the INTCAL13 calibration curve (Reimer et al. 2013) (Table 2).
Pollen analysis and microscopic charcoal
Pollen samples were extracted from the sediment core using a small sampler of defined volume (~0.3 cm3) about every 5 cm of the core. Samples were prepared for pollen analysis following Fægri and Iversen (1989) with the use of sodium polytungstate heavy liquid separation to remove clastic material. A known quantity of exotic spores (Lycopodium) was added to each sample (~2,136 spores per sample) using a suspension based on Lycopodium tablets (~10,679 spores per tablet). The pollen residues were mounted in glycerine. A light microscope with a magnification of 400× was used for routine pollen analysis. In the top and bottom samples of the core, some corroded pollen was observed although in general the preservation of pollen and spores in the profile was excellent. Pollen types were identified using the identification keys of Moore et al. (1991) and Beug (2004). The basic pollen sum includes all regional terrestrial pollen taxa typical for the Early Holocene, excluding Alnus (alder) pollen and Dryopteris-type spores but including Poaceae (grasses). Pollen and spores of the local aquatic or mire vegetation [including Cyperaceae (sedges)] were excluded from the pollen sum. The average number of counted pollen grains within the pollen sum is 200.
Furthermore, other microfossils or so-called non-pollen palynomorphs (such as algae and spores of coprophilous fungi) were identified using Pals et al. (1980), van Geel (1978, 2001), van Geel et al. (1981, 1989, 2003) and van Geel and Aptroot (2006). Non-pollen palynomorphs can give additional information about the local vegetation, fire activity and presence of dung of large herbivores.
Microscopic charcoal (pieces larger than 27 µm, but smaller than 250 µm because that was the sieve mesh size for preparation of the palynological samples) was counted relative to Lycopodium spores (number of charcoal pieces per 100 Lycopodium spores) to quantify fire activity in the Tungelroyse Beek area. In addition, Pediastrum colonies were counted with respect to Lycopodium spores.
Zonation of the regional pollen diagrams was mainly based on changes in the arboreal pollen/non-arboreal pollen (AP/NAP) ratio and trends in the percentages of Betula, Pinus and Corylus, all taxa that characteristically dominate the Late Glacial and Early Holocene pollen records in the Netherlands (Hoek 1997).
Organic content
LOI data provide information on the nature of the lithology of the Tungelroyse Beek. The organic content (in weight percentage) of the samples taken from the core from the Tungelroyse Beek was determined by loss on ignition (LOI). In this respect, samples were taken from the core every 2 cm in the lowest 20 cm of the core and every 1 cm in the remaining part of the core. The samples were dried (at 105 °C overnight to remove water from the sample) and ashed (4 h at 550 °C) to burn the organic matter from the sample. The weight difference between the weight of the ashed sample and the weight of the dried sample is the LOI.
Magnetic susceptibility
Magnetic susceptibility (MS) data provide information on the nature of the lithology of the Tungelroyse Beek. MS was measured every 1 cm of the core. It is a measure of the amount of magnetic material present, which is detected when subjected to a magnetic field. The magnetization is ultimately related to the concentration and composition (size, shape and mineralogy) of magnetisable material in the sample. Samples that are rich, per unit volume, in magnetisable substances will yield high readings. Samples that are poor in magnetisable material will yield lower or even negative values. Usually, clays yield higher values than sands. However, it is also possible that nutrient-rich seepage water causes metal ions, particularly iron and manganese, to precipitate as e.g. iron carbonate, which may yield higher values than expected.
Carbonate content
In the field it was noted that siderite (iron carbonate) was present in the peat layer of Pit 1, probably as a result of seepage. The presence of siderite could possibly interfere with the magnetic susceptibility signal. Although siderite was not observed in the field at Pit 2 where the core was taken, it could be present there as well. It was therefore decided to investigate the presence of carbonates in the core between 25.03 m +AOD and 24.53 m +AOD, taking samples every 2 cm. Carbonate content was determined by using the Scheibler method. By treating samples with HCl, carbonates present in the samples reacted with HCl to form Ca2+, Mg2+ and Fe2+ and CO2 gas, respectively. The escaping CO2 gas that was formed by adding HCl to the sample was measured volumetrically and results in a measure of the amount of carbonates present in the sample.
Results
The palynological results together with magnetic susceptibility, LOI and microscopic charcoal record of the Tungelroyse Beek core are shown in Figs. 6 and 7, which display the regional and local pollen records, respectively. Radiocarbon dates are displayed as well.

Pollen diagram for regional taxa, radiocarbon dates, magnetic susceptibility, loss on ignition, microscopic charcoal and carbonate content of the Tungelroyse Beek core. Alnus and indeterminate pollen are excluded from the pollen sum. The thick dashed line indicates the tentative position of red deer remains based on 14C dating

Pollen diagram for local taxa, radiocarbon dates, magnetic susceptibility, loss on ignition, microscopic charcoal and carbonate content of the Tungelroyse Beek core. The thick dashed line indicates the tentative position of red deer remains based on 14C dating
The carbonate content in the analysed samples appeared to be very low. Of 26 carbonate content samples, 21 samples had a carbonate content below 1 %, four samples had a carbonate content below 2 % and only one sample has a carbonate content of 2.5 % (Fig. 6). In addition, no trend in carbonate content can be observed. Because of this, it is assumed that magnetic susceptibility values are not significantly influenced by iron carbonates possibly present in the sediments.
The Tungelroyse Beek regional PAZ (pollen assemblage zones) can be linked with the Late Glacial and Early Holocene regional pollen zonation scheme of Hoek (1997) and those of the palaeoecological studies ‘De Borchert’ (van Geel et al. 1981) and nearby Haelen (Bos et al. 2007). The latter is located close to the Tungelroyse Beek. The Tungelroyse Beek regional PAZ and magnetic susceptibility, LOI and microscopic charcoal record are therefore described according to the pollen zonation schemes of the previously mentioned studies.
Zone Tung 1: Late Younger Dryas, below 24.43 m +AOD
The zone is characterized by relatively high NAP values, predominantly Poaceae. Upland herbaceous pollen values, especially of Artemisia (mugwort) and Apiaceae, (parsley family) are relatively high as well. These taxa indicate an open vegetation type. Pinus (pine) and Betula (birch) are the dominant arboreal taxa, although not abundant. Pediastrum was observed in this zone indicating nutrient-rich water conditions at the site. Myriophyllum verticillatum (whorled water milfoil) indicates seepage and aquatic conditions at the site. The occurrence of this species is often linked to the presence of siderite. However, the results obtained with the Scheibler method do not suggest significant amounts of siderite at the sampling site. The presence of hyphopodia of Gaeumannomyces (HdV-126), which has sedge (Carex) as a host plant, together with the high values of sedge (Cyperaceae) pollen suggest that sedges were part of the local wetland vegetation surrounding the open water. The various ascospores of coprophilous fungi recorded in this zone (i.e. Sordaria-type, Cercophora-type and Arnium-type; Table 3) suggest that dung of large herbivores (game) was dropped in the wetlands in the vicinity of the coring location.
The low arboreal pollen values in this zone and the following increase in zone Tung 2 point to the Younger Dryas period (PAZ-3b of Hoek 1997). The absence of Populus (poplar) pollen, relatively low birch values, high NAP (Artemisia and Poaceae) values and the presence of a Betula nana (dwarf birch) fruit in the 14C sample suggest a Younger Dryas age as well (cf. Bos et al. 2005). The values of Empetrum nigrum-type pollen (black crowberry) and Juniperus (juniper) are relatively low, probably caused by the moist and humic conditions in the vicinity of the site during the Early Holocene which, on the other hand, caused the good preservation conditions.
The LOI values and magnetic susceptibility values in zone Tung 1 are low. This indicates that the sediments of the Tungelroyse Beek stream are mainly sandy with presumably large quartz and low clay content. High Lycopodium spore values indicate a low pollen concentration, probably caused by a high accumulation rate (i.e. sand) during zone Tung 1. A low LOI could indicate a relatively high stream velocity in the Tungelroyse Beek. However, a low LOI together with the fact that the vegetation was relatively open could also suggest that windy conditions caused relatively high percentages of clastic coversand, in which the entire Tungelroyse Beek valley is situated, to have been deposited in the Tungelroyse Beek during Zone Tung 1. This suggests that the sediment that accumulated during zone Tung 1 dates in the later, dryer part of the Younger Dryas. Especially during the later part of the Younger Dryas, aeolian activity became more prominent because of an increased aridity (Kasse 1999). In the Netherlands, aeolian deposition during this period occurred especially in topographically higher places within the coversand area and along river valleys, forming river dunes of up to 10 m high (Kasse 2002).
Microscopic charcoal concentrations are relatively high in zone Tung 1. Moreover, charred epidermis fragments of grasses and sedges and the presence of Gelasinospora spores suggest that fires occurred in the area. Fire activity may have been relatively high during this phase. However, deposition of wind-blown microscopic charcoal particles may have been enhanced during this dryer period. Peaks in arboreal pollen, Poaceae and dry herbs curves do not coincide clearly with peaks in charcoal concentration.
Zone Tung 2a: Friesland Phase of the Preboreal (24.43–24.68 m +AOD)
The boundary between zone Tung 1 and zone Tung 2 is characterized by a decrease in the Poaceae and other NAP pollen percentages, and an associated rise in Betula values. Zone Tung 2a is dominated by Betula. This indicates the development from open vegetation, dominated by grasses and dry herbs, towards Betula woodland. The Pinus curve shows persistent low values during this phase. Towards the end of zone Tung 2a, the values of Betula decline and NAP (especially Poaceae) values increase. Sedges remained part of the local wetland vegetation as is suggested by the high values of Cyperaceae pollen and presence of hyphopodia of Gaeumannomyces (HdV-126). Zone Tung 2a is comparable with the Friesland phase (Behre 1966; van Geel et al. 1981) of the Preboreal (PAZ 4a, Hoek 1997), which starts at ca. 10,150 bp (Hoek 1997). In addition, the boundary between Tung 1 and Tung 2 is comparable with the boundary between PAZ 3 and PAZ 4a as described by Hoek (1997).
In zone Tung 2a, vegetation started to develop towards a forest and organic production increased as a consequence of higher temperatures. As a result, soils stabilized and the organic content of the sediments started to rise gradually as is visible in the LOI curve. Microscopic charcoal concentrations are highest in zone Tung 2a. Fire activity is therefore considered to have been relatively high. Although the Poaceae curve shows some peaks during the same phase, and a small peak in Apiaceae is observed in this phase as well, these do not coincide clearly with peaks in charcoal concentration. The arboreal pollen curves do not show an irregular pattern coinciding with peaks in the charcoal curve either. The observed fire activity therefore cannot directly be linked to the observed changes in the vegetation pattern in this phase. The pollen data and microscopic charcoal concentrations probably reflect regional fires and/or regional changes in the vegetation cover. Finds of charred epidermis fragments of grasses and sedges indicate that during this period, (natural or man-induced) fires in the reed swamp vegetation in the area occurred. In addition, large herbivores were present in the valley during this zone, as is suggested by finds of ascospores of coprophilous fungi such as Sordaria-type (HdV-55A) and Cercophora-type (HdV-112).
Zone Tung 2b: Rammelbeek Phase of the Preboreal (24.68–24.94 m +AOD)
Poaceae values increase and high NAP percentages are recorded, while the Betula values decrease. This suggests a development towards more open vegetation and regional expansion of grasses, while forest development was interrupted. The slight increase in typical light-demanding taxa such as Artemisia, Rumex acetosa/acetosella (sorrel) and Galium-type and the re-occurrence of Helianthemum (rock-rose) indicates a development towards a more open vegetation type as well. Local vegetation is dominated by aquatics with a change from Myriophyllum verticillatum to Nymphaea (white water lily) indicating a shift to stagnant water and possibly decreased seepage as a result of a drier climate.
The regional expansion of grasses suggests that this subzone represents the Rammelbeek phase of the Preboreal (Behre 1966). A similar development can be seen in the record of the Borchert during the Rammelbeek phase (van Geel et al. 1981; Bos et al. 2007).
The presence of high Cyperaceae values and hyphopodia of Gaeumannomyces (HdV-126) indicates that during this zone sedges still remained part of the local vegetation around the sample site, although their importance in the wetland vegetation was decreasing. Various ascospores of coprophilous fungi were recorded, which may suggest that the wetlands were used as foraging and drinking places by large herbivores. The ascomycete fungus Bombardioidea-type, of which ascospores were recorded in this zone, is dung-inhabiting, and it may be associated with the presence of European elk (Alces alces) (HdV-575, Bos et al. 2005). The records of charred epidermis fragments of grasses and sedges suggest that during the Rammelbeek phase—and also during the previous Younger Dryas and Friesland phase—fires (natural or man-induced) occurred in the reed swamps of the area. During these periods active burning of reed swamps by Late Palaeolithic or Early Mesolithic people (sensu Starr Carr in Mellars and Dark 1998) may have taken place. There is however, no archaeological evidence to support this. At the transition from subzone 2a to 2b, the deposits change from humic sand to sandy peat, and LOI values increase slightly during this subzone, indicating further soil development. Microscopic charcoal concentrations are lower and less irregular than during the Friesland phase and decreasing, but still relatively high.
Zone Tung 3: Late Preboreal (24.94–25.48 m +AOD)
This zone is dominated by Pinus. In addition, an increase in Salix (willow) pollen and Dryopteris-type spores can be observed, suggesting that these taxa became an important component in the local vegetation, when a fern-rich willow carr started to develop. The local diagram shows a decrease in the values of Nymphaea and Myriophyllum verticillatum. This indicates that the hydroseral succession process advanced further in the residual channel of the Tungelroyse Beek, although Salix pollen and Dryopteris-type spores still indicate rather moist conditions. A similar signal was observed by Bos et al. (2007) at other Dutch locations.
Increased arboreal pollen indicates that vegetation further developed into a pine-birch forest. At the boundary between Tung 3 and Tung 4, a slight increase in non-arboreal pollen can be observed. Tung 3 is comparable with the Late Preboreal PAZ 5 (Hoek 1997), starting at the 14C plateau at ca. 9,600–9,500 bp (van Geel et al. 1981; Hoek 1997; Bos et al. 2007).
During Tung 3, accumulation of organic matter in the sediments occurs faster compared to zones Tung 1 and 2. Furthermore, at the start of Tung 3 there is a transition from sandy peat to peat, as can also be recognized by increasing LOI values. This lithological change at the start of the Late Preboreal can be observed as well in other Dutch Preboreal records (Bos et al. 2007). At many locations, Sphagnum peat growth started or higher lake water tables are recorded (van Geel et al. 1981; Hoek and Bohncke 2002), which are probably indications for a sudden change to a more humid climate (van der Plicht et al. 2004; Bos et al. 2007).
Microscopic charcoal concentrations are low compared to Tung 1 and Tung 2, although a minor peak can be observed on the boundary between zone Tung 3 and zone Tung 4. This peak coincides with the slight increase in non-arboreal pollen. Single finds of charred epidermis fragments of grasses and sedges indicate that during this period fires in the reed swamp vegetation occurred only occasionally. This is probably related to a change in the wetland vegetation from reed swamp to a fern-rich willow carr. Large herbivores, as reflected by the spores of coprophilous fungi, remained present in the Tungelroyse Beek valley during this zone.
Zone Tung 4: Boreal (25.48–25.69 m +AOD)
From 25.48 m +AOD upwards, the pollen diagram shows increasing Corylus (hazel) values, indicating the start of the Boreal (Hoek 1997). This event is dated at ca. 9,100 bp (van Geel et al. 1981). Quercus (oak) and Ulmus (elm) are still absent but appear shortly after the appearance of Corylus, at a depth of 25.60–25.66 m +AOD which is dated at ca. 9,000 bp (van Geel et al. 1981). The Tilia (lime) curve starts at 25.66–25.70 m +AOD, dated at ca. 8,300 bp (van Geel et al. 1981).
LOI values remain high during zone Tung 4, although between 25.45 and 25.70 m +AOD, a sudden decrease in LOI values can be observed which could indicate a dry phase. Lycopodium spore values indicate a low accumulation rate during zone Tung 4. The local pollen diagram shows a low amount of aquatics (e.g. Nymphaea) and a decrease in riparian vegetation (e.g. Filipendula, Rumex aquaticus and Sparganium (bur-reed)). Dryopteris-type spores and Salix pollen decrease as well. These observations suggest further infilling of the Tungelroyse Beek residual channel and possibly a transition towards a drier climate. Microscopic charcoal concentrations are relatively low after they peaked at the boundary between zone Tung 3 and zone Tung 4. Large herbivores remained present in the area during this zone.
Zone Tung 5: Atlantic (25.68–26.05 m +AOD)
Alnus appears at 25.66-25.70 m +AOD, marking the beginning of the Atlantic. van Geel et al. (1981) dated the immigration of Alnus at ca. 7,900 bp. Alnus could settle in moist places, especially in stream valleys, at the expense of Betula and Salix (van Geel et al. 1981). The lithology shows the presence of many macroremains from this depth onwards, which are probably roots of Alnus. In addition, an increase of Quercus and Ulmus can be observed during this phase. Finds of fungal spores of Kretzschmaria deusta (HdV-44; van Geel 1978) may be related to the appearance of Alnus as well, as the species is a host to this fungus. However, K. deusta may also have grown on Salix, Ulmus and Tilia. Apiaceae increase as well, but this could also be a local signal from the riparian vegetation. Furthermore, the increase of Tilia coincides with a slight decrease of the Corylus percentages, which was also observed by van Geel et al. (1981).
LOI values remain high in zone Tung 5, whereas microscopic charcoal concentrations are relatively low throughout the whole zone. In the uppermost part of zone Tung 5 LOI values are lower. This coincides with a significant quantity of corroded pollen in the upper 10–20 cm, which indicates oxidation of this part of the core. In addition, up to ~25.50 m +AOD, some corroded pollen can be observed. Low values of Lycopodium spore sums could indicate a low accumulation rate in zone Tung 5, although perhaps slightly higher than in zone Tung 4. It should be noted that an overall increase of tree pollen also results in low Lycopodium records. Aquatics such as Nymphaea and riparian herbs such as Filipendula reach very low values or are even absent in zone Tung 5 and suggest further infilling of the site. Dryopteris-type spores and Salix pollen values are low as well. These taxa indicate an advanced state of the hydroseral succession process during this zone. Large herbivores remained present in the area.
Discussion
Microscopic charcoal particle size
In this study, microscopic charcoal counts of particles of 27–250 µm are used as a proxy for fire activity in the vicinity of the investigated site. According to several studies, small microscopic charcoal fragments, (<100, <125, <200 µm) originate from more distant rather than local fires (e.g. Whitlock and Larsen 2001). This would suggest that such small charcoal fragments are inappropriate for reconstructing local fire activity. However, Innes and Simmons (2000) state that charcoal data almost certainly comprise a composite assemblage with a complex taphonomy. Micro-charcoal frequencies are a combination of local and regional signals (Innes and Simmons 2000). Small charcoal abundances may therefore be better regarded as a general, relative estimate of overall fire activity in the region, thus including local fire activity. The microscopic charcoal particle sizes used in this study as a proxy for local man-made fire activity therefore seem valid, although a possible more regional origin of microscopic charcoal particles should be taken into consideration.
Palynological and radiocarbon age models
The characteristics of the pollen diagram of the Tungelroyse Beek valley generally correspond with other pollen diagrams of the Netherlands covering a similar period (e.g. Hoek 1997; van Geel et al. 1981; Bos et al. 2007). Based on the similarity of the pollen diagram of Tungelroyse Beek and the other diagrams, an age model based on palynological and lithological evidence was tentatively made in the Results section (Figs. 6, 7).
The AMS date from 24.36–24.38 m +AOD depth generally fits the palynological results. The pollen assemblage suggests a Younger Dryas phase age for this sample, which would indicate an age between 10,950 and 10,150 bp for this sample. Indeed, the AMS age of ~10,540 bp thus fits with the Younger Dryas age.
The AMS date of the sample from 24.84–24.85 m +AOD depth is ~10,140 bp. This age dates the sample at the start of the Holocene. Palynology however, suggests a Rammelbeek phase age for this sample, so an age between ~9,950 and ~9,750 bp (Hoek 1997) which is at least ~200 year younger than the AMS date. Lithology does not support the AMS date either. If the sample indeed originated from the beginning of the Holocene, lithology would most likely have shown a clear transition to more organic-rich deposits at this depth, which is not the case. Remarkably however, it can be seen in the literature (van der Plicht et al. 2004; Bos et al. 2007) that similar radiocarbon age outliers during the Rammelbeek phase can also be found at other sites.
The AMS sample at 25.23–25.24 m +AOD depth appears to be situated in the middle of the late Preboreal part of the pollen record (Fig. 6). According to van Geel et al. (1981) and Hoek (1997) the Late Preboreal starts at 9,500 bp. The AMS date of ~9,640 bp thus seems to be at least 150 yr too old. However, the zone transition between the Rammelbeek and Late Preboreal phase lies within a 14C plateau (9,600–9,500 bp). Samples from different ages can therefore provide similar 14C dates during this interval. This is also the case at many other Dutch locations (Bos et al. 2007).
Compared to the other diagrams, the Boreal (zone Tung 4) is rather compact. The curves of Tilia and Alnus start at the same depth although they should be ~400 years apart, and the Alnus curve shows a sharp entrance into the pollen diagram. In addition, the AMS date just before the assumed beginning of the Atlantic (25.63–25.64 m +AOD) provides an age of ~9,070 bp, which is ~1000 years older than the 7,900 bp suggested by van Geel et al. (1981) for the rise of Alnus at the beginning of the Atlantic. These facts combined indicate the presence of a hiatus in the Late Boreal part of the diagram. Hiatuses occur more often in the Boreal period, which was supposedly a dry period (Hoek and Bohncke 2002; Bos et al. 2007).
It should be noted that three out of four AMS dates do not correspond completely with the expected age derived from biostratigraphy. However, as was mentioned before, similar age offsets can be found in other Early Holocene sites as well. As can be derived from the Haelen site (Bos et al. 2007) for example, even AMS dates of an undisturbed profile can be quite erratic. An alternative explanation might be linked to old carbon uptake by Carex and other riparian plants, in calcareous environments. Turney et al. (2000) report comparable age offsets of several hundreds of years between AMS dates from Salix and Carex from the same stratigraphic levels in Lateglacial calcareous deposits.
Human activity at the time the red deer died
As the youngest red deer remains, dated around 6,570 bp, are younger than the material investigated in the core, they are not further discussed. The oldest red deer remains, dated around 9,650 bp, however, do fit in the investigated period (Fig. 7). The trends in the pollen diagram (zone Tung 3) show that the vegetation did not change significantly around this period and microscopic charcoal concentrations are low. The pollen diagram, the concentrations spores of coprophilous fungi and the microscopic charcoal record thus do not suggest increased human activity around this period.
This does not necessarily mean that no human activity occurred in the area at this time. First of all, it should be noted that Pit 2, where the analysed core was taken, is located southwest of Pit 1, against the prevailing wind direction. Pollen and charcoal from the surroundings of the possible human settlement at Pit 1, where Early Mesolithic artefacts were discovered, may therefore not have reached the location of Pit 2. Moreover, Pit 2 is located 150 m away from Pit 1. This distance may be too far and therefore the palynological evidence from Pit 2 does not necessarily reflect human activity around Pit 1. Hicks (1993) for instance showed that localized impact on the vegetation of northernmost Finland by hunter-gatherers is discernible by pollen analysis only within a distance of some 50 m (Hicks 1993). Also Bos and Janssen (1996) obtained their pollen evidence over a distance of less than 150 m. They concluded that the detection of human induced small local vegetation changes is thus strongly dependent on the distance between the area of activity and the area from which the samples were derived. Sampling should therefore take place within the immediate vicinity of the settlement. Defining the exact location of the supposed Early Mesolithic settlement is however not that straightforward. Some finds of supposed Early Mesolithic age were discovered in Pit 1, but no evidence for a settlement was found there.
No settlement may have been present in the vicinity of both Pit 1 and Pit 2, or a settlement may have been located a few hundred meters away from Pit 2. The area could also have been a hunting area rather than an area used for a settlement. Bos and Janssen (1996) concluded that sampling preferably should take place from several cores at various distances from a settlement, thereby allowing a useful comparison between local and regional pollen deposition and thus revealing small-scaled vegetation patterns (Bos and Janssen 1996). Sampling of multiple cores was however not done in the present study.
Human induced fires or natural fires?
Using the increased microscopic charcoal concentrations as an indicator for both local and regional fires, the records of coprophilous fungi as a sign for the presence of game, and corresponding vegetation changes as an indicator for human activity, seems valid. Fire ecology (the burning of vegetation in order to create open patches in the vegetation for non-agricultural reasons (Out 2009)) is a method of vegetation management often suggested to be practised, especially during the Mesolithic. For example, for the Starr Carr site in England, Mellars and Dark (1998) suggested that active burning of reed swamp was used to keep the occupation area largely free of reeds and other vegetation, and thus to maintain visibility and easy access to a lake. In addition, the purpose of deliberate burning of reed swamp vegetation may have been to create openings in the vegetation that induced the growth of herbs, forbs and shrubs, which are attractive to herbivores. Clearance may have facilitated hunting, and enabled people to move around easier (Bos and Urz 2003). In this way fire may have been used as a direct hunting technique. Similar evidence for burning of reed swamps was also found at Zuthphen, in the Netherlands (Bos et al. 2005).
During the Mesolithic, reed vegetation was commonly present around Dutch wetland sites. Burning of eutrophic wetland vegetation may have been related to management of natural vegetation (sensu Starr Carr in Mellars and Dark (1998); see also Bos et al. 2007; Out 2009). Therefore, charred epidermis fragments of Poaceae and Cyperaceae were also recorded in the pollen slides (besides charcoal fragments originating from wood). Poaceae and Cyperaceae may have been part of the former reed swamp vegetation. Charred epidermis fragments of sedges and grasses are mainly concentrated in the lower part of the core, corresponding with the period from the Younger Dryas to the older part of the Late Preboreal. Epidermis fragments were absent from the layers were the bones were found.
The records of charred epidermis fragments of grasses and sedges however, may suggest that during the Younger Dryas, the Friesland and the Rammelbeek phases, natural or man-induced fires occurred in the reed swamps of the area. It is thus possible that active burning of reed swamps took place by Late Palaeolithic or Early Mesolithic people (sensu Starr Carr in Mellars and Dark 1998). There is however no archaeological evidence, as Late Palaeolithic and Mesolithic sites were not recovered in the Tungelroyse Beek valley. This suggests that the fires probably had natural causes. Furthermore, it should be noted that an increase in fire activity does not necessarily indicate an increase in human activity. In a study of modern day fire frequencies in Boreal forests, Senici et al. (2010) not only found a correlation between wildfire occurrence and human activity, but also between wildfire occurrence and vegetation type, drainage patterns and changing climate. During the Early Holocene, natural fires that started in the woods by lightning possibly occurred more frequently, because climate may have been more continental (Huntley 1993). This may have been the case especially during zone Tung 1 and Tung 3 (Younger Dryas and Rammelbeek phase) as during these periods, vegetation was relatively open and climate was windy and dry. As can be seen in Figs. 6 and 7, during these periods the microscopic charcoal record shows relatively high values.
Conclusions
The palynological record of the Tungelroyse Beek is generally comparable to Early Holocene records of other sites in northwestern Europe. It encompasses the Younger Dryas to Atlantic period. A hiatus is present in the Late Boreal. AMS dates generally correspond with the age model based on biostratigraphical evidence, although AMS ages and palynology derived ages are not completely similar. Similar offsets between the two age models are however also found at other comparable Dutch sites.
Vegetation developed from dry open grasslands in the Younger Dryas towards a less dry birch forest during the Friesland phase. During the Late Preboreal, vegetation developed towards a Pinus dominated forest, while local taxa indicate a hydroseral succession of the Tungelroyse Beek residual channel. In the Boreal, a mixed temperate forest developed, hosting species like Corylus, Quercus, Ulmus and Tilia. This vegetation development continued in the Atlantic period, when deciduous forest with shade tolerant trees developed while Alnus appeared at moist locations.
The palynological data combined with microscopic charcoal concentrations (including the charred epidermis fragments of grasses and sedges), and records of spores of coprophilous fungi and LOI values do not point to increased Early Mesolithic human activity in the vicinity of the investigated site during the time that the red deer possibly was killed. The investigated records do show that during the Younger Dryas, Friesland and Rammelbeek phase fire activity was higher in comparison to the later periods. However, it is questionable if fire activity was caused by human activity. The dry and windy climate is more likely to have caused fires to spread. In addition, it is questionable if this fire activity occurred close to the investigated site during this time. The investigated microscopic charcoal record most likely includes both local and regional fires.
Finds of bones of several larger mammals, including red deer at two dated levels, and the presence of spores of coprophilous fungi present throughout the investigated core suggest that the Tungelroyse Beek valley was a favourable place for large herbivores (game) to visit over a long period (at least ca. 4000 years). As game was certainly present in the valley during the Early Holocene, it certainly would have attracted Mesolithic man. The presence of undated bones with cut-marks thus confirms the presence of Early Mesolithic people in this area. The fact that their impact on the vegetation is not visible in the pollen data of the present study suggests that their impact on the vegetation was very low. Moreover, the distance between where the core was collected and where the activity took place was possibly too large and against the prevailing wind direction to allow it to be recorded in the sediments.
References
Behre K-E (1966) Untersuchungen zur spätglazialen und frühpostglazialen Vegetationsgeschichte Ostfrieslands. Eiszeitalt Ggw 17:69–84
Beug H-J (2004) Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete. Pfeil, München
Bos JAA, Janssen CR (1996) Local impact of Palaeolithic Man on the environment during the end of the Last Glacial in The Netherlands. J Archaeol Sci 23:731–739
Bos JAA, Urz R (2003) Late Glacial and early Holocene environment in the middle Lahn river valley (Hessen, central-west Germany) and the local impact of early Mesolithic people—pollen and macrofossil evidence. Veget Hist Archaeobot 12:19–36
Bos JAA, Verbruggen F (2012) Pollen onderzoek. In: Opbroek M, Lohof E (eds) Tijd in centimeters. Een kijkje in het landschap van een dekzandrug te Almere; een inventariserend veldonderzoek in de vorm van proefsleuven en een hoogwaardig booronderzoek. ADC Rapport 2662. ADC, Amersfoort
Bos JAA, van Geel B, Groenewoudt BJ, Lauwerier RCGM (2005) Early Holocene environmental change, the presence and disappearance of early Mesolithic habitation near Zutphen (The Netherlands). Veget Hist Archaeobot 15:27–43
Bos JAA, van Geel B, van der Plicht J, Bohncke SJP (2007) Preboreal climate oscillations in Europe: wiggle-match dating and synthesis of Dutch high-resolution multi-proxy records. Quat Sci Rev 26:1,927–1,950
de Kort JW (2010) Mildert, dal van de Tungelroysche Beek (TUBE-11). Veldwerkverslag profielopname en monstername. Intern verslag Rijksdienst voor het Cultureel Erfgoed. Rijksdienst voor het Cultureel Erfgoed, Amersfoort
de Moor JJW, Bos JAA, Bouman MTIJ, Moolhuizen C, Exaltus R, Maartense FPA (2009) Definitief Archeologisch Onderzoek in het tracé van de Hanzelijn in het Nieuwe Land: een interdisciplinaire geo-archeologische waardering van het begraven landschap van Oostelijk Flevoland. Deltares, Delft
Edwards KJ (1989) Meso-Neolithic vegetational impacts in Scotland and beyond: Palynological considerations. In: Bonsall C (ed) The mesolithic in Europe. Donald, Edinburgh, pp 143–155
Fægri K, Iversen J (1989) In: Fægri K. Kaland PE, Krzywinski K (eds) Textbook of pollen analysis. 4th edn. Wiley, Chichester
Groenewoudt BJ, Deeben J, van Geel B, Lauwerier RCGM (2001) An early Mesolithic assemblage with faunal remains in a stream valley near Zutphen, the Netherlands. Arch Korrespondenzbl 31:329–348
Hamburg T, Müller A, Quadflieg B (eds) (2012) Mesolithisch Swifterbant. Mesolithisch gebruik van een duin ten zuiden van Swifterbant (8300-5000 v. Chr.). Een archeologische opgraving in het tracé van de N23/N307, provincie Flevoland. Archol Rapport 174, ADC Rapport 3250:157–241
Hicks S (1993) Pollen evidence of localized impact on the vegetation of northernmost Finland by hunter-gatherers. Veget Hist Archaeobot 2:137–144
Hoek WZ (1997) Late-glacial and early Holocene climatic events and chronology of vegetation development in the Netherlands. Veget Hist Archaeobot 6:197–213
Hoek WZ, Bohncke SJP (2002) Climatic and environmental events over the Last Termination, as recorded in The Netherlands: a review. Neth J Geosci 81:123–137
Huntley B (1993) Rapid early-Holocene migration and high abundance of hazel (Corylus avellana L.): alternative hypotheses. In: Chambers FM (ed) Climate change and human impact on the landscape. Chapman & Hall, London, pp 205–215
Innes JB, Simmons IG (2000) Mid-Holocene charcoal stratigraphy, fire history and palaeoecology at North Gill, North York Moors, UK. Palaeogeogr Palaeoclimatol Palaeoecol 164:151–165
Kasse C (1999) Late Pleniglacial and Lateglacial aeolian phases in The Netherlands. In: Schirmer W (ed) Dunes and fossil soils. GeoArchaeoRhein 3:61–82
Kasse C (2002) Sandy aeolian deposits and environments and their relation to climate during the Last Glacial Maximum and Lateglacial in northwest and central Europe. Prog Phys Geogr 26:507–532
Louwe Kooijmans LP, van den Broeke PW, Fokkens H, van Gijn AL (2005) The prehistory of the Netherlands. Amsterdam University Press, Amsterdam
Mellars P, Dark P (1998) Star Carr in context: new archaeological and palaeoecological investigations at the Early Mesolithic site of Star Carr, North Yorkshire. McDonald Institute Monographs, McDonald Institute for Archaeological Research, Cambridge
Moore PD, Webb JA, Collinson ME (1991) Pollen analysis, 2nd edn. Blackwell, Oxford
Moree JM, Sier MM (eds) (2014) Twintig meter diep! Mesolithicum in de Yangtzehaven-Maasvlakte te Rotterdam. Landschapsontwikkeling en bewoning in het Vroeg Holoceen. BOOR Rapport 523. BOOR, Rotterdam
Out W (2009) Sowing the Seed? Human Impact and Plant Subsistence in Dutch Wetlands during the Late Mesolithic and Early and Middle Neolithic (5500–3400 Cal BC). University Press, Leiden
Pals JP, van Geel B, Delfos A (1980) Paleoecological studies in the Klokkeweel bog near Hoogkarspel (prov. of Noord Holland). Rev Palaeobot Palynol 30:371–418
Price TD (1987) The Mesolithic of Western Europe. J World Prehist 1:225–335
Reimer PJ, Bard E, Bayliss E et al (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55:1,869–1,887
Rijks Geologische Dienst (1933) Geologische kaart van Nederland: 58 Roermond, kwartblad III. Topografische Dienst, ‘s Gravenhage
Roymans JAM (2013) Hert in de pot, herinrichting Tungelroyse Beek deeltrajecten C, D, en E, gemeenten Nederweert en Leudal. Archeologisch onderzoek: een archeologische begeleiding van de graafwerkzaamheden. RAAP Rapport 2680. RAAP, Weesp
Roymans J, de Nutte G (2012) Het edelhert komt naar je toe: Mesolithische sporen van jacht in het beekdal van de Tungelroyse beek. Westerheem 61:82–89
Senici D, Chen HYH, Bergeron Y, Cyr D (2010) Spatiotemporal Variations of Fire Frequency in Central Boreal Forest. Ecosystems 13:1,227–1,238
Simmons IG (1993) Vegetation change during the Mesolithic in the British Isles: some amplifications. In: Chambers FM (ed) Climate change and human impact on the landscape. Chapman & Hall, London, pp 109–118
Smit BI, Deeben J, van Doesburg J, Rensink E, ter Schegget M, Theunissen EM (eds) (2013) Beschermingsprogramma archeologie 2013. Selectievoorstel voor 28 nieuwe archeologische rijksmonumenten. Rapportage Archeologische Monumentenzorg 213. Rijksdienst voor het Cultureel Erfgoed, Amersfoort
Smith AG, Whittle A, Cloutman EW, Morgan LA (1989) Mesolithic and Neolithic activity and environmental impact on the south-east fen-edge in Cambridgeshire. Proc Prehist Soc 55:207–249
Törnqvist TE, de Jong AFM, Oosterbaan WA, van der Borg K (1992) Accurate dating of organic deposits by AMS 14C measurement of macrofossils. Radiocarbon 34:566–577
Turney CSM, Coope R, Harkness DD, Lowe JJ, Walker MJC (2000) Implications for the dating of Wisconsinan (Weichselian) late-glacial events of systematic radiocarbon age differences between terrestrial plant macrofossils from a site in SW Ireland. Quat Res 53:114–121
van der Linden M (2011) Palynologisch onderzoek aan een veen- en kleipakket uit het Laat-Mesolithicum bij Almere-De Vaart. BIAXiaal 501. BIAX, Zaandam
van der Plicht J, van Geel B, Bohncke SJP, Bos JAA, Blaauw M, Speranza AOM, Muscheler R, Björck S (2004) The Preboreal climate reversal and a subsequent solar forced climate shift. J Quat Sci 19:263–269
van Geel B (1978) A palaeoecological study of Holocene peat bog sections in Germany and The Netherlands. Rev Palaeobot Palynol 25:1–120
van Geel B (2001) Non-Pollen palynomorphs. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments, vol 3., Terrestrial, algal, and siliceous indicatorsKluwer Academic Publishers, Dordrecht, pp 99–119
van Geel B, Aptroot A (2006) Fossil ascomycetes in Quaternary deposits. Nova Hedwigia 82:313–329
van Geel B, Bohncke SJP, Dee H (1981) A palaeoecological study of an upper Late Glacial and Holocene sequence from “De Borchert”, The Netherlands. Rev Palaeobot Palynol 31:367–448
van Geel B, Coope GR, van der Hammen T (1989) Palaeoecology and stratigraphy of the Lateglacial type section at Usselo (The Netherlands). Rev Palaeobot Palynol 60:25–129
van Geel B, Buurman J, Brinkkemper O, Schelvis J, Aptroot A, van Reenen GBA, Hakbijl T (2003) Environmental reconstruction of a Roman Period settlement site in Uitgeest (The Netherlands), with special reference to coprophilous fungi. J Archaeol Sci 30:873–883
Whitlock C, Larsen C (2001) Charcoal as a fire proxy. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments, vol 3., Terrestrial, algal, and siliceous indicatorsKluwer, Dordrecht, pp 75–97
Whittle AWR (1996) Europe in the Neolithic: the creation of new worlds. Cambridge University Press, Cambridge
Woldring H, Schepers M, Mendelts J, Fens R (2012) Camping and foraging in Boreal hazel woodland—the environmental impact of Mesolithic hunter-gatherers near Groningen, the Netherlands. In: Niekus MJLT, Barton RNE, Street M, Terberger T (eds) A mind set on flint. Archaeological Studies 16. Barkhuis, Eelde, pp 381–392
Acknowledgments
We would like to thank the two anonymous reviewers and the editor for their thorough reviews which have significantly improved the quality of the manuscript. In addition, we would like to thank Jan Lenaerts (Utrecht University) for his technical assistance in producing Fig. 2.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by K.-E. Behre.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Woelders, L., Bos, J.A.A., de Kort, JW. et al. Early Holocene environmental change and the presence of Mesolithic people in the Tungelroyse Beek valley near Mildert, the Netherlands. Veget Hist Archaeobot 25, 177–189 (2016). https://doi.org/10.1007/s00334-015-0543-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-015-0543-x