
Abstract
Nests of birds often possess a diverse invertebrate fauna, but almost no descriptions of the invertebrate fauna of bird nests from the High Arctic exist in spite of numerous studies at lower latitudes. Seventy-seven nests belonging to common eider, barnacle goose, glaucous gull, black-legged kittiwake and snow bunting were examined for the invertebrate fauna from Kongsfjorden and Longyearbyen, Svalbard. Twenty-seven nest-living species were found and thirty-one species in soil under common eider nests. The diversity in most of the nests was poor. True nidiculous species were absent, and ectoparasites dominated in nests of all bird species; the flea; Ceratophyllus vagabundus vagabundus in the nests of common eider, barnacle goose and glaucous gull, Mioctenopsylla arctica arctica in the nests of black-legged kittiwake; and the parasitic mesostigmatid mite Dermanyssus hirundinis in the nests of snow bunting. The most diverse group in nests was opportunistic soil-living oribatid mites. If soil samples taken beneath common eider nests are included, five species or genera of invertebrates not previously recorded from Svalbard were found: Cyrtozetes sp., Liochthonius simplex (Acari: Oribatida), Protaphorura subuliginata (Collembola: Onychiuridae) and larvae of Parochlus kiefferi and Paralimnophyes sp. (Diptera: Chironomidae). Of these, Cyrtozetes sp. is probably an undescribed species. The chironomid larvae from black-legged kittiwake nests have probably been transported together with nest material. With the exception of the ectoparasites, little indicates that invertebrates preferentially exploit nests in Svalbard, as most species were free living and are normally common in soils.
Similar content being viewed by others

Introduction
Bird nests can harbour rich invertebrate faunas (e.g. Hicks 1959, 1962, 1971; Ambros et al. 1992; Tryjanowski et al. 2001; Bajerlein et al. 2006; Sinclair and Chown 2006: Turienzo and Di Iorio 2008). Several factors combine to cause nests to be particularly attractive for invertebrates; left over food scraps brought to the chicks, favourable thermal conditions due to the incubating birds (Sinclair and Chown 2006) and, for the parasites, the birds themselves. While some of the invertebrates are attracted to the nest, others are introduced accidentally with nest material. Certain nest-living species may be beneficial, such as scavenging beetles removing food scraps and decaying organic material from the nest which may reduce disease amongst chicks (Neubig and Smallwood 1999; Majka et al. 2006).
Phoresy can in its simplest form be defined as one animal transported by another (Perotti and Braig 2009), and such fortuitous phoresy with birds has been proposed to be an important mode of dispersal for soil invertebrates to remote regions such as the islands of the Arctic (Lebedeva and Krivolutsky 2003; Lebedeva and Lebedev 2008). It has even been suggested that oribatid mites can complete their life cycle in the plumage of birds (Krivolutsky and Lebedeva 2004a; Shakhab 2006). Bird nests could provide favourable points of establishment in an otherwise often harsh environment for invertebrates dispersing to the High Arctic (Coulson et al. 2009). If this is the case, new or rare invertebrates for Svalbard might be observed within, or close to, nests of migratory birds.
In contrast to nests at lower latitudes, little is known about the communities of invertebrates present in High Arctic bird nests (Lebedeva and Lebedev 2008; Coulson et al. 2009). Only five studies with limited sampling and taxonomic extent are reported from Svalbard (Elton 1925; Cyprich and Krumpal 1991; Mehl 1992; Lebedeva et al. 2006; Coulson et al. 2009). Of these, only two studies describe invertebrates other than fleas (Lebedeva et al. 2006; Coulson et al. 2009). Coulson et al. (2009) found the invertebrate fauna of the nests of glaucous gull (Larus hyperboreus), black-legged kittiwake (Rissa tridactyla) and common eider (Somateria mollissima) to be species poor, mainly consisting of the flea Mioctenopsylla arctica arctica and six species of oribatid mites. However, this study had a restricted sample size. Lebedeva et al. (2006) reported five species of oribatid mite from nests of snow buntings (Plectrophenax nivalis) and arctic tern (Sterna paradisaea). Beyond these two studies, little is known of the nidiculous invertebrate fauna of High Arctic birds in Svalbard.
Several bird species in Svalbard show high nest fidelity or nest in colonies, for example the black-legged kittiwake. For these species, nests may grow in size each year, creating a distinctive habitat for invertebrates. With only low vegetation in High Arctic environments such as Svalbard, birds nest on the open tundra, on cliffs or exploit crevices under rocks or in scree slopes. Due to climate change, the Arctic is warming and the summer period prolonged (AMAP 2011). Such increase in temperature and potential extension of the summer period may enable new species of invertebrate to colonise Svalbard. However, this is dependent on the animals being able to disperse to the remote archipelago. Phoresy with birds may be one possible route permitting rapid colonisation of High Arctic latitudes. Five species of bird were selected with nests representative of each nest type and nesting location: (a) on open tundra: common eider and barnacle goose (Branta leucopsis), (b) on cliff tops or large ledges: glaucous gull, (c) nests constructed on cliff ledges: black-legged kittiwake and (d) in rocky crevices such as in talus slopes: snow bunting. In addition, these birds include both marine (common eider, black-legged kittiwake and glaucous gull) and terrestrial species (barnacle goose and snow bunting).
We aim here to (a) provide a detailed description of the fauna of nests from diverse migratory bird species with widespread breeding colonies in Svalbard, (b) identify differences in the invertebrate fauna of these nests and point to potential causes for faunal differences, (c) compare the nest fauna with the known soil and ground fauna of Svalbard and determine whether there are invertebrate species inhabiting these specialised microhabitats not known from other localities in Svalbard and (d) discuss whether invertebrates may disperse to Svalbard via phoresy with birds and if nests may facilitate colonisation.
Materials and methods
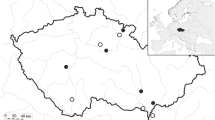
Svalbard is situated in the north-western region of the Barents Sea between 74 and 81°N and from 10 to 35°E, approximately 700 km north of the nearest mainland, Norway (Fig. 1). The land area covers approximately 63,000 km2, and about 60 % is permanently covered by ice and snow (Hjelle 1993). The summer is short, and only 4 months, June–September, have mean air temperatures above 0 °C (Norwegian Meteorological Institute 2013). Precipitation is generally low, annually between 190 mm (Svalbard airport) and 525 mm (Ny-Ålesund) (Norwegian Meteorological Institute 2013).

Location of Svalbard (box) and sampling sites at Longyearbyen and Ny-Ålesund
The five investigated bird species belong to three different families of birds: Anatidae, common eider and barnacle goose; Laridae, black-legged kittiwake and glaucous gull; and Emberizidae, snow bunting. These birds constructed dissimilar types of nests, and sampling was conducted in the summer 2010 (Table 1). All the investigated bird species are common or abundant breeders in Svalbard (Kovacs and Lydersen 2006). The snow bunting is the only passerine bird that breeds regularly in Svalbard migrating from wintering grounds believed to be on the Russian steppes north of the Caspian Sea and in Kazakhstan (Cramp and Perrins 1994; Bakken et al. 2006). In Svalbard, the population is estimated at 1,000–10,000 pairs. The common eider in Svalbard belongs to the sub-species S. mollissima borealis and spends the winter along the coast of northern Norway and Iceland (Bakken et al. 2003). In Svalbard, this species normally breeds on small islands to avoid predation of arctic foxes and can breed together with other birds such as barnacle goose or arctic terns (Ahlen and Andersson 1970). In 2010, more than 2,500 pairs of common eider were breeding in Kongsfjorden (Norwegian Polar Institute 2014), and the total breeding populations, including Bear Island, have been estimated to 13,500–27,500 pairs. The Svalbard barnacle goose population is geographically isolated from the populations breeding in Greenland, northwest Russia and the Baltic, and overwinters on the Solway Firth on the English–Scottish border (Owen and Black 2005) where the population consists of 27,000 individuals. The black-legged kittiwake has a circumpolar distribution and breeds on narrow ledges or outcrops of cliffs in the western Atlantic in Nova Scotia, Newfoundland and Greenland east to Iceland, the British Isles, France and Spain, and northeast to Norway and the Barents Sea. Outside the breeding season, they disperse over most of the North Atlantic (Kovacs and Lydersen 2006). The population for Svalbard is estimated to 180,000 birds. The glaucous gull is one of the largest gulls in Svalbard and an important avian predator with a population in Svalbard of between 4,000 and 10,000 pairs. This gull has a circumpolar breeding distribution. The Svalbard population spends the winter dispersed throughout the North Atlantic both along the coast and in open waters (Kovacs and Lydersen 2006) and belongs to the sub-species L. hyperboreus gunnerus. Glaucous gulls breed alone or in small colonies often on elevated points close to, or in, other seabird colonies.
Most snow bunting nests were collected in Adventdalen close to Longyearbyen, nests from other species were collected on islets in Kongsfjorden, close to Ny-Ålesund (Fig. 1). On the islet Storholmen where barnacle goose and common eider nests were investigated, 175 pairs of goose and around 900 common eiders were breeding.
To avoid disturbance of incubating birds, different methods were used to sample nest material (Table 1). Nests that were in use were vacuumed with a battery-powered pooter (Hausherr’s Machine Works, USA) while parts of abandoned nests were collected from barnacle goose and common eiders. Nest material (down) was taken from common eider and barnacle goose nests shortly after the birds had abandoned their nest. Approximately half a 20 × 30 cm plastic bag was taken with down from each of these nests. If the down had largely been blown from the nest scoops, down still remaining was taken. It is likely that the soil under the nests can be influenced by species occurring in the nests and vice versa; therefore, soil samples were taken under nests of common eider with a 35 mm corer to compare to the nest fauna. Approximately 20–100 cm3 soil was taken for each sample. This was only done for one bird species due to animal welfare considerations (most of the birds were incubating or they re-use old nests), logistic problems in collecting such samples from all birds and limitations in extracting capacity. Complete nests of snow buntings were collected (see Gwiazdowicz et al. 2012 for full details). Due to difficulties in accessing natural snow bunting nests, nests were collected from the nesting boxes established on the cableway trestles in Adventdalen close to Longyearbyen. In addition, nests were sampled from one nest box in Ny-Ålesund and one in the settlement of Longyearbyen (Fig. 1).
Samples were stored at outdoor temperatures or in a +5 °C room at the University Centre in Svalbard (UNIS). The soil and down samples and whole snow bunting nests were extracted into 96 % alcohol using Tullgren funnels (Burkard Scientific Ltd., Uxbridge, UK) at UNIS. Each sample was extracted until the material was completely dry and no more invertebrates were emerging. This method of extraction relies on the active movement of animals away from the source of light and heat, but is not optimal for some taxa, such as nematodes and annelids, which can be underrepresented in the extracted material. Pootered material was sorted under a stereomicroscope as this material was difficult to extract. It was hence difficult to discriminate between individuals being dead or alive at point of sampling. After sorting, all specimens were maintained in 96 % alcohol. Due to the large amount of mites in the snow bunting nest samples, a sub-sampling method (Southwood and Henderson 2000) was used to estimate mite density. The samples were scanned for rare species to avoid only sub-sampling the most common ones. Damaged or regurgitated insects and spiders that probably have been brought to the nest as food for the chicks were excluded. However, whole insects which probably were alive at point of sampling were included, and chironomid head capsules believed not to be food for chicks.
Mites that could not be identified under a stereomicroscope were mounted on slides in Hoyer’s medium or lactic acid. Oribatid nymphs were only identified to genus and were assumed to belong to the same species as the adults when only one species within the genus was present in the bird colony. When more than one species within the genus was present, juveniles were noted as unidentified representatives of the genus. Collembola were identified by A. Fjellberg (Arne Fjellberg Entomological Research).
Systematics for Astigmatina (synonym Acaridei, Acaridida, Astigmata) and Prostigmata (synonym Promatides) follows Lindquist et al. (2009). Nomenclature for Oribatida follows Behan-Pelletier (1985, 1997), Weigmann (2006) or Colloff (1993). Mesostigmatid mites follow Ávila-Jiménez et al. (2011) and Gwiazdowicz et al. (2011a, b). Collembola Fjellberg (1998, 2007) and Siphonaptera follow Brinck-Lindroth and Smit (2007).
All material is deposited in various reference collections at the Department of Biology, University of Bergen, Norway; Department of Arctic Biology, UNIS, Norway; Arne Fjellberg Entomological Research, Tjøme, Norway (Collembola); Poznan University of Life Sciences, Poznan, Poland (Mesostigmata).
Statistical analyses
Direct ordination analyses were used to evaluate which predictors best explain the variation in species composition of invertebrate nest fauna. Specimens not determined to species were excluded from analysis unless they represented genus or taxa different from determined species. Chironomid species were not included as these probably were not part of the nest fauna, but rather brought in with nest material and dead when sampled. Two samples from glaucous gull nests were empty and could not be included in the statistical analysis as the ordinations do not accept empty samples (Lepš and Šmilauer 2003). Due to different sampling methods, and large variability in the number of specimens in each sample, presence–absence data were used in the ordination analyses rather than the actual species counts.
A preliminary detrended correspondence analysis (DCA) was performed prior to all ordination analyses to decide whether a linear or unimodal model should be chosen. The default DCA settings were used in R (v. 2.10.1), except that rare species were down-weighted. For the full dataset, the DCA axis was long (gradient length of 1 axis = 3.768) and the unimodal models, constrained correspondence analysis (CCA), were chosen to perform the analyses (Lepš and Šmilauer 2003). Prior to direct ordination (CCA), indirect ordination correspondence analysis (CA) was used to identify potential outliers. One of the snow bunting samples was excluded from the dataset since outliers can highly influence the analysis and make the detections of gradients difficult (Palmer 2010). A sample was identified as an outlier when it occurred at a distance away from the other samples in the CA plot, which were shown as one tight cluster. The nests identified as outliers differed from other nests by the presence of only Dermanyssus hirundinis and unidentified mesostigmatid mites, which were not included in the analyses.
In order to evaluate which predictor variables best explain the species composition, permutation tests in CCA were used. The following predictors were evaluated: bird species, type of sample (soil, nest or pootered material), location, weather, breeding period of the bird (abandoned nests, egg or chicken period) and date of sampling. The R default number of 199 permutations was used to test all variables separately and to choose the optimal combination of explanatory variables to explain the variance in species composition. The latter was done by backward elimination using the step function in R where the default selection is performed using the Akaike information criterion (AIC) (Oksanen et al. 2010), favouring the model with the lowest possible AIC.
To be consistent, CCA was also used when analysing a subset of the data only including black-legged kittiwake samples (gradient length of DCA 1 axis = 2.354). On this subset, a permutation test of CCA was used to test for significance between the two colonies (Blomstrandhalvøya and Observasjonsholmen) using location as a predictor to test if the species composition was affected by the colonies being situated on two different islets. The vegan library (Oksanen et al. 2010) in R was used to run the different multivariate analysis (CCA, CA and DCA).
To investigate to what degree the nest and soil fauna was properly sampled for the different bird species, taxon accumulation curves were made using the function specaccum in the vegan library in R. Samples were added in random order with 100 permutations. Same dataset was used as for the multivariate analyses, but empty samples and the outlier were included.
Results
Species composition
In total, 45 species of invertebrates were recorded in the soil and nest samples. In addition, 10 unidentified taxa were found and seven chironomid species that we consider unlikely to be part of the nest fauna not included in the species counts. Twenty-seven species were recorded in nests, in addition to the unidentified taxa. The greatest diversity occurred in soil samples with 31 species (Table 3), where the Collembola were the most species-rich group (15 species). Soil samples were dominated by Collembola common for Svalbard, except for Protaphorura subuliginata which is recorded for the first time from the archipelago. The greatest diversity of mesostigmatid mites was also found in soil samples (Table 3).
Most nests were species poor, with fleas being the dominant species for all bird nests except the snow bunting (Table 2). The flea Ceratophyllus vagabundus vagabundus was found in nest samples from all bird species except black-legged kittiwakes where M. a. arctica occurred. No bird species had both species of fleas present. The maximum abundances of C. v. vagabundus were found in the nests of common eider and barnacle goose with up to 463 and 276 adults, respectively (Table 2). Most of these nests contained fleas, but with large variation in numbers. Adult fleas were most common, in contrast to the pootered samples from glaucous gull and black-legged kittiwake where larva dominated. The body condition of dead fleas suggests that they had died recently, probably during the pooting process or during sample storage prior to examination. A few C. v. vagabundus specimens were also present in snow bunting nests. This is the first record of C. v. vagabundus in relation to snow buntings in Svalbard. M. a. arctica was only found in nests of black-legged kittiwakes and is the most common species in the nests with flea larva being present in ≥90 % of the nests of both colonies.
Eight mesostigmatid mite species were recorded, but with the exception of D. hirundinis, this group was largely restricted to soil samples (Table 3). Two of the species are parasitic: D. hirundinis dominated with abundances exceeding 8,000 individuals per snow bunting nest, while a second parasitic species, Haemogamasus ambulans, was only found in two soil samples (Table 3). Oribatid mites were the most species-rich group in the nests, with 16 species, and the most common taxon after fleas. The black-legged kittiwake has the greatest diversity of oribatid mites of the bird species studied, with 15 species present, and three species were only found here (Dissorhina ornata, Platynothrus punctatus and Hermannia scabra) (Table 2). Species composition was slightly different between the two black-legged kittiwake colonies with 13 oribatid mite species being present in Observasjonsholmen and 10 in Blomstrandhalvøya. Seven of these occurred in only one of the two colonies (Table 2). A unidentified oribatid mite species Cyrtozetes sp. (Mycobatidae) was present in the Observasjonsholmen black-legged kittiwake colony and from the nest and soil sample of one common eider. This species is new to Svalbard and most likely an undescribed species (V. Behan-Pelletier pers. comm.). The oribatid mite species Liochthonius simplex found in a barnacle goose nest is also recorded for the first time in Svalbard.
In the Blomstrandhalvøya black-legged kittiwake colony, chironomid head capsules (Diptera: Chironomidae) from larva were common and belong to seven different species; two that have not been recorded in Svalbard before are Parochlus kiefferi and Paralimnophyes sp. (Table 2).
The CCA model which included both bird species and sample type gave the best model for explaining the variance in species composition for the whole dataset. The other predictors: location, weather, breeding period of the bird and date of sampling, were not significant when using backward elimination and hence not included in the model. The final model explained 27.8 % of the total variance in species composition (Table 4). Soil samples and samples from black-legged kittiwakes come out as most different from the others in the CCA plot (Fig. 2), dominated by unique species not occurring in the other samples (cluster A and B in Fig. 2a). In the CCA species plot (Fig. 2a), cluster A consists largely of Collembola species and other species unique for soil samples (i.e. Enchytraeidae worms and most of the mesostigmatid mites) while cluster B consists of species only found in black-legged kittiwake nests, including the flea M. a. arctica, a few oribatid mites and a Collembola. The species composition of the barnacle goose nests is intermediate between the soil sample and the nest samples from the common eiders (Fig. 2b), and in general, samples from the different bird species seem to group together in the CCA plot, indicating similar species composition. However, for black-legged kittiwakes, the two colonies were significantly different with regard to species composition (permutation test of CCA, p = 0.015) and location explained 8.5 % of the variance between them.

Constrained correspondence analysis (CCA) of chosen model with bird species and sample type as significant predictors (backward elimination: p < 0.05) using presence–absence of species. a Species distribution. Species abbreviations and species belonging to cluster A and B are shown in Tables 2, 3. b Sample distribution. Glaucous gull and black-legged kittiwake samples were taken with an electric pooter, nest material was taken from common eider, barnacle goose and snow bunting nests and extracted in Tullgren funnels, and common eider soil samples were taken under the common eider nests. The plots are slightly modified to make them readable
All taxon accumulation curves suggest that more taxa would be found with a higher sampling intensity (Fig. 3). This is especially the case for the snow bunting where there is no indication of an asymptote in the curve. This is also the bird with the fewest nests sampled. Common eider, barnacle goose and glaucous gull had quite similar number of species if similar numbers of samples are compared. For glaucous gull, the accumulation shows a tendency to flattening out and could indicate that most of the species potentially found in these nests have been recorded (Fig. 3). The highest number of species when standardising the sampling effort was in black-legged kittiwake nests, but soil samples had most identified species in total.

Taxon accumulation curves of number of invertebrate species in nests from different bird species and soil under common eider nests in Svalbard 2010. Samples were added in random order using 100 permutations, showing the mean species accumulation curves and their standard deviations. Chironomid species were not included, as they were assumed not to be part of the nest fauna (limnic larva), and specimens not determined to species were excluded unless they represented genus or taxa different from determined species
Discussion
Parasites
In nests of all bird species, ectoparasites dominated in terms of abundance and prevalence. Fleas were present in nests of all the bird species examined, and except for the snow bunting, they were present in most of the nests and often in high densities (≥100 fleas were found in single nests). While M. a. arctica only occurred in nests of black-legged kittiwakes, the flea C. v. vagabundus was present in the nests of the other studied species. C. v. vagabundus is a common ectoparasite of gulls, ducks and geese (Brinck-Lindroth and Smit 2007). In both common eider and barnacle goose nests, fleas were highly aggregated in some nests. C. v. vagabundus was for the first time recorded in nests of snow buntings in Svalbard, although in low numbers. Coulson et al. (2009) made a pioneer study of invertebrates in nests of common eider, black-legged kittiwakes and glaucous gull breeding in Kongsfjorden. They also concluded that the nest fauna was meagre, mainly consisting of the flea M. a. arctica and a few oribatid mites.
High density of fleas in nests of black-legged kittiwakes is common also in mainland, Norway (Mehl 1992). High flea infestations may reduce breeding success in some species of bird (Fitze et al. 2004), including geese breeding in the Arctic such as Ross’s goose (Chen rossii) and lesser snow goose (Chen caerulescens) (Harriman and Alisauskas 2010). The dominance of fleas in the nests is not unexpected since the two flea species in Svalbard are specialised nest dwellers that reproduce in, and naturally spend much of their life in the nests of their avian hosts (Brinck-Lindroth and Smit 2007). It is not known whether the fleas survive the winter in the nests in Svalbard or whether they have to be reintroduced each year by their returning hosts. In more temperate areas, it is common that after the birds have left the nest, they complete their larval development and remain quiescent in the cocoons until temperature or mechanical disturbance triggers the emergence of adults (Humphries 1968; Nielsen 1976).
In the nests of the snow bunting, the mesostigmatic mite D. hirundinis dominated and was present in high numbers in all nests examined. It is a nidicolous species that feeds on a range of different bird species. Its presence in snow bunting nests is discussed fully by Gwiazdowicz et al. (2012), and this mite may be the reason why the snow buntings in Svalbard construct a new nest for the second clutch. Little is known about the ecology of D. hirundinis (Roy and Chauve 2007), but the related species Dermanyssus gallinae is known to have significant detrimental effects on the host (Chauve 1998). Apart from D. hirundinis, mesostigmatid mites were largely lacking from nests, and this is in contrast to bird nests at lower latitudes in Europe (Ambros et al. 1992; Tryjanowski et al. 2001; Błoszyk et al. 2009; Krištofík et al. 2009). The second parasitic mesostigmatic mite species observed was H. ambulans often found on mammals and occasionally in the nests of birds (Bregetova 1956).
Oribatida
Seventeen species of oribatid mites were recorded, and all were present in nests except L. lapponicus, which was found only in soils. Oribatid mites are common in the upper layers of soil, and in moss and lichen. The species observed are generally widespread in Svalbard (Bayartogtokh et al. 2011; Coulson et al. 2014). However, L. simplex and Cyrtozetes sp. were both recorded in Svalbard for the first time. Oribatid species diversity in nests of barnacle goose, common eider and snow bunting was low but not unusual when compared to studies of soil fauna in Svalbard (Coulson and Refseth 2004 and references therein).
The low diversity of oribatid mites found by Coulson et al. (2009) in black-legged kittiwake colonies (only Ameronothrus lineatus and Diapterobates notatus) may be due to lower sample size, but maybe more to that Coulson et al. (2009) extracted the nest material in Tullgren funnels and hence only collected individuals that were alive during extraction. In the present study, all pootered material was hand-sorted from black-legged kittiwake and glaucous gull so that dead mites and exoskeletons were included. High occurrence of old dead mites might imply that species being transported to the nest along with nest-building material do not thrive in the nests and die shortly after arrival. Lebedeva and Lebedev (2008) suggested that nests probably are more favourable for desiccation resistant species of oribatid mites as nests can be very dry.
Collembola
Collembola were found in high densities and diversity in the soil samples but were absent from the majority of nests. Collembola have previously been recorded in nests (Elton 1925; Hicks 1959; Ohyama and Hiruta 1995, Krivolutsky and Lebedeva 2004a), but the species are often not mentioned. It appears that Collembola do not frequently exploit bird nests in Svalbard, probably due to xeric conditions. Collembola are in general considered to be less desiccation resistant than, for example, oribatid mites, as most have limited control of water loss (Harrisson et al. 1991; Hodkinson et al. 1994; Elnitsky et al. 2008). The collembolan P. subuliginata was present in a soil sample, and although this species previously has been found in the archipelago (A. Fjellberg, unpublished material), the identity of the specimens was uncertain. It has previously been collected in rich vegetation sites in Svalbard and is not restricted to bird colony sites. P. subuliginata is not a typical Arctic species, but is known from Arctic regions in Eastern Europe, northern Norway and Greenland (Babenko and Fjellberg 2006).
Other groups
The lack of beetles in the nests was surprising as Coleoptera are commonly found in bird nests in temperate regions (Hicks 1959; Hågvar 1975; Majka et al. 2006; Turienzo and Di Iorio 2008; Krištofík et al. 2009). Only one damaged staphylinid beetle was found in a glaucous gull nest. The lack of beetles may be partly due to the low diversity of beetles in Svalbard: only 19 species (Coulson et al. 2014). Although some species can be abundant under bird cliffs (Hågvar 1971), there is no indication that they are attracted to nidicolous microhabitats in Svalbard. In general, there seems to be a lack of predatory species in the studied nests. Moreover, chewing lice (Phthiraptera: Mallophaga) were largely absent although several species have been recorded from Svalbard (Mehl et al. 1982). It is likely that most lice remain on the birds, rather than in the nests (Bush et al. 2001), and hence do not occur in the samples collected in this study.
Some of the chironomid head capsules found in kittiwake nests belonged to two species not previously recorded in Svalbard: P. kiefferi and Paralimnophyes sp. (Coulson 2007). Almost all the head capsules of chironomids were old, and it is unlikely that adult chironomids with aquatic larvae select to lay eggs in dry nests. Hence, it is more likely that the larvae have been transported to the nest along with building material, as black-legged kittiwakes can collect dripping wet moss along the waterline of freshwater bodies to bring to their nests (pers. obs.). The chironomid larvae in Table 2 should therefore more properly be regarded as parts of the nest material.
Statistical analyses and general observations
Differences in invertebrate community compositions were found between nests of different bird species, but were also to some degree affected by sampling procedure. 27.8 % of the variation in species composition could be explained by the CCA model with sample type and bird species as significant predictors. Occurrence of ectoparasites can be explained by the presence of the host bird species (Bush et al. 2001), but it is likely that other environmental factors play important roles in determining the overall invertebrate fauna. Different birds varied in species composition, and the soil samples and black-legged kittiwake samples were clearly distinguishable in the CCA from the other bird species by having many unique species and high species diversity. True nidicolous species (Krištofík et al. 2009) were absent. The studied bird species had different types of nests built from differing materials (Table 1), and it is likely that several of the invertebrates can have been transported to the nests of glaucous gull, black-legged kittiwake and snow bunting with local plant material used for constructing the nests. Different types of tundra in the Kongsfjorden area can harbour between 42,000 and 518,000 microarthropod individuals per m2 (Bengtson et al. 1974), and Seniczak and Plichta (1978) found that abundance of oribatid mites in moss and lichen in Hornsund in Svalbard largely was dependent on the plant composition of the tundra. Coulson et al. (2003) found that the invertebrate fauna of the soil varied over short distances partly related to the higher plant cover. It is therefore likely that the plant material used to construct the nest is important for the invertebrate species composition. According to Elton (1925), black-legged kittiwakes on Bear Island have been observed to carry moss some kilometres to their nests, and hence, invertebrates transported with nest material may potentially travel substantial distances.
The low species number in nests of barnacle goose and common eider could be due to that down is an unnatural habitat for most oribatid mites and Collembola. The high diversity of Collembola and mesostigmatid mites found immediately beneath the nests of common eider seems to support this. It is also likely that invertebrates brought to the nest will seek out more favourable conditions if the nest material itself is sub-optimal. The taxon accumulation curves (Fig. 3) also indicate that more species would probably have been recorded if we had the possibility to sample more nests. Glaucous gulls and black-legged kittiwakes build perennial nests where plant material can accumulate over several years, creating a more stable environment than the contemporary down nests of common eider and goose. The down of the latter species is blown away shortly after the birds leave. Microclimate in the nest is influenced by the incubating birds and presence of chicks (Sinclair and Chown 2006; Błoszyk et al. 2009). Since black-legged kittiwake chicks remain in the nest 5–6 weeks after hatching, this can possibly have a positive influence on the invertebrate fauna, similarly to the temperature ameliorated nests of wandering albatross (Sinclair and Chown 2006) in the sub-Antarctic islands.
Great diversity of uropodine mites has been observed in the large perennial nests belonging to white stork (Błoszyk et al. 2005) and birds of prey (Gwiazdowicz et al. 2006). Glaucous gulls make large nests, and although black-legged kittiwake nests are relatively small, they can be constructed very close together and could possibly function as large nests for invertebrates. However, location of the colony can also have significant effect, as demonstrated for the two black-legged kittiwake colonies (Blomstrandhalvøya and Observasjonsholmen colony) that had significantly different species composition.
Unfortunately, it was not possible to sample the nest material from all bird species the same way in our study due to animal welfare. It was therefore difficult to test whether nests made out of various material, or were of different age (annual or perennial), contributed to the dissimilar species compositions. The perennial nests made out of plant material were all pootered, and nest material was extracted from common eider and goose, which make annual nests. Snow buntings make annual nests out of plant material, but samples were few and limited by the number of birds occupying the nest boxes. It is therefore likely that some of the variation explained by sampling method in the CCA is due to other factors related to the nest, such as pootered nest, were also perennial and made out of plant material.
Bird phoresy?
In our study, five invertebrate species not previously recorded in Svalbard were found. An intriguing question is whether these species could indicate bird phoresy as none of them are considered to be associated with bird nests. Two species were the oribatid mites L. simplex and Cyrtozetes sp. found in nests. In contrast to mesostigmatid, prostigmatid and Astigmatina mites, few species of oribatid mites seem to have morphological adaptations to attach to other animals (Norton 1980). Norton (1980) points out that passive dispersal by nest-building birds and rodents is probably a common phenomenon. More recent studies indicate that oribatid mites are also capable of bird phoresy and hence can be present in plumage of a range of different birds (Krivolutsky and Lebedeva 2004a, b). L. simplex (Brachychthoniidae) is a semi-cosmopolitan species, present in Palaearctic, Nearctic (Greenland), Oriental countries and Australia (Subías and Shtanchaeva 2011), and other Brachychthoniidae have been found in bird plumage (Krivolutsky and Lebedeva 2004a, b; Lebedeva et al. 2006). The genus Cyrtozetes includes four described species with distributions limited to Japan, northern Alaska, the Russian Far East and British Columbia (Behan-Pelletier and Eamer 2008). The individuals from our study are thus the first record of this genus in Europe and the first from bird nests (V. Behan-Pelletier, pers.comm.). As the individuals of Cyrtozetes collected from Svalbard do not conform to the morphology of the described species, it is probably an undescribed species (V. Behan-Pelletier pers.comm.). The finding of this species in nests of two different bird species, common eider and black-legged kittiwake, can indicate that the species is established in Svalbard. Further studies are required to resolve whether these species are mainly associated with bird colony sites and hence possibly can have arrived with the birds or whether it has only been overlooked in other studies in the Kongsfjorden area.
Tectocepheus velatus, which was recorded in black-legged kittiwake and snow bunting nests, is a cosmopolitan species that on several occasions has been recorded in the plumage and nests of birds (Tryjanowski et al. 2001; Lebedeva and Krivolutsky 2003; Krivolutsky and Lebedeva 2004a, b; Lebedeva et al. 2006; Shakhab 2006; Lebedeva and Lebedev 2008). In Svalbard, T. velatus has previously been found in the nest and plumage of arctic tern (Lebedeva et al. 2006). It has even been hypothesised that it is able to fulfil its life cycle in the plumage of birds (Krivolutsky and Lebedeva 2004a). Krivolutsky and Lebedeva (2004a, b) found T. velatus in the plumage of 32 species of passerine birds and 29 non-passerines. This high occurrence can possibly indicate that T. velatus regularly use birds as a dispersal vector, but could also reflect that it is a very common and numerous species.
The Astigmatina mites that were found in nests and soil samples under common eider and barnacle goose were not determined to species, but phoresy is well known in this taxon (O’Connor 2009; Perotti and Braig 2009). They are often phoretic on flies or birds in certain developmental stages, having morphological adaptations to attach to vectors. Although no Astigmatina mite species have been listed from Svalbard (Coulson and Refseth 2004; Coulson 2007), unidentified individuals were found by Lebedeva and Lebedev (2008) in plumage of birds from Spitsbergen. It is not unlikely that the Astigmatina mites can have been transported with the birds, but they can also have been overlooked in Svalbard, as many studies are restricted to certain species or taxa, or because few parts of Svalbard have been properly sampled (Coulson 2013; Coulson et al. 2014).
Conclusions
The ectoparasites that dominated in the nests varied between the different species of bird. A remaining question is whether the various nest parasites survive the Svalbard winter in perennial nests, whether birds bring parasites in their plumage each spring, or both. Apart from the parasites, the invertebrate fauna was characterised by free living invertebrates that do not have any strong relationship with birds or bird nests. Low diversity was found in the down nests of barnacle goose and common eider despite that these nests are placed on the ground in close proximity to soil invertebrates in the surrounding environment. The highest diversity was found in the cliff nesting black-legged kittiwake nests. It is likely that this difference is associated with the different types of nest material used and that perennial nests are more favourable for invertebrates. Collembola were largely lacking from nests. Furthermore here were few predatory species, such as predatory mesostigmatid mites or beetles. When soil samples beneath eider duck nests were included, five invertebrate species new to Svalbard were recorded. Two of these might have been transported with birds, the oribatid mites Cyrtozetes sp. and L. simplex. Recent studies indicate that bird phoresy can be more common than previously thought. However, the nest fauna was not greatly dissimilar to the community expected in the vicinity of the nests, and there is little direct evidence that nests are acting as points of establishment for invertebrate species imported with the migrating birds. Nonetheless, care must be taken in drawing conclusions due to the poor state of knowledge of the Svalbard invertebrate fauna and that these species may have previously been overlooked elsewhere.
References
Ahlen I, Andersson A (1970) Breeding ecology of an Eider population on Spitsbergen. Ornis Scand 1:83–106
AMAP (2011) Arctic Monitoring and Assessment Programme 2011. SWIPA 2011. Snow, water, ice and permafrost in the Arctic. Arctic Monitoring and Assessment Programme (AMAP), Oslo
Ambros M, Krištofík J, Šustek Z (1992) The mites (Acari, Mesostigmata) in the birds’ nests in Slovakia. Biológia (Bratislava) 47:369–381
Ávila-Jiménez ML, Gwiazdowicz DJ, Coulson SJ (2011) On the mesostigmatid (Acari: Parasitiformes) mite fauna of Svalbard: a revised checklist of a High Arctic archipelago. Zootaxa 3091:33–41
Babenko A, Fjellberg A (2006) Collembola Septentrionale. A catalogue of springtails of the Arctic regions. KMK Scientific Press, Moscow
Bajerlein D, Błoszyk J, Gwiazdowicz DJ, Ptaszyk J, Halliday B (2006) Community structure and dispersal of mites (Acari, Mesostigmata) in nests of the white stork (Ciconia ciconia). Biologia 61:525–530
Bakken V, Runde O, Tjørve E (2003) Norsk ringmerkingsatlas, vol 1. Stavanger Museum, Stavanger
Bakken V, Runde O, Tjørve E (2006) Norsk ringmerkingsatlas, vol 2. Stavanger Museum, Stavanger
Bayartogtokh B, Schatz H, Ekrem T (2011) Distribution and diversity of the soil mites of Svalbard, with redescriptions of three known species (Acari: Oribatida). Int J Acarol 37:467–484
Behan-Pelletier VM (1985) Ceratozetidae of the western north-American Arctic. Can Entomol 117:1287–1366
Behan-Pelletier VM (1997) Oribatid mites (Acari: Oribatida) of the Yukon. In: Danks HV, Downess JA (eds) Insects of the Yukon. Biological Survey of Canada (Terrestrial Arthropods), Ottawa, pp 115–149
Behan-Pelletier VM, Eamer B (2008) Mycobatidae (Acari: Oribatida) of North America. Can Entomol 140:73–110
Bengtson SA, Fjellberg A, Solhøy T (1974) Abundance of tundra arthropods in Spitsbergen, Norway. Entomol Scand 5:137–142
Błoszyk J, Gwiazdowicz DJ, Halliday B, Dolata PT, Goldyn B (2009) Nests of the black stork Ciconia nigra as a habitat for mesostigmatid mites (Acari: Mesostigmata). Biologia 64:962–968
Błoszyk J, Gwiazdowicz DJ, Bajerlein D, Halliday B (2005) Nests of the white stork Ciconia ciconia (L.) as a habitat for mesostigmatic mites (Acari, Mesostigmata). Acta Parasitol 50:171–175
Bregetova NG (1956) Gamasid mites (Gamasoidea). The short key. Izdatelstvo Akademii Nauk CCCP, Moskva (in Russian)
Brinck-Lindroth G, Smit FGAM (2007) The fleas (Siphonaptera) of Fennoscandia and Denmark. Brill Academic Publishers, Leiden-Boston
Bush AO, Fernández JC, Esch GW, Seed JR (2001) The Arthropoda. Parasitism: the diversity and ecology of animal parasites. Cambridge University Press, New York, pp 225–284
Chauve C (1998) The poultry red mite Dermanyssus gallinae (De Geer, 1778): current situation and future prospects for control. Vet Parasitol 79:239–245. doi:10.1016/S0304-4017(98)00167-8
Colloff MJ (1993) A taxonomic revision of the oribatid mite genus Camisia (Acari: Oribatida). J Nat Hist 27:1325–1408
Coulson SJ (2007) Terrestrial and freshwater invertebrate fauna of the High Arctic Archipelago of Svalbard. Zootaxa 1448:41–68
Coulson SJ (2013) The terrestrial invertebrate fauna of the Svalbard archipelago in a changing world; history of research and challenges. Can Entomol 145:131–146. doi:10.4039/tce.2012.110
Coulson SJ, Refseth D (2004) The terrestrial and freshwater invertebrate fauna of Svalbard (and Jan Mayen). In: Prestrud P, Strøm H, Goldman H (eds) A catalogue of the terrestrial and marine animals of Svalbard. Norwegian Polar Institute, Tromsø, pp 57–122
Coulson SJ, Hodkinson ID, Webb NR (2003) Microscale distribution patterns in High Arctic soil microarthropod communities: the influence of plant species within the vegetation mosaic. Ecography 26:801–809
Coulson SJ, Moe B, Monson F, Gabrielsen GW (2009) The invertebrate fauna of High Arctic seabird nests: the microarthropod community inhabiting nests on Spitsbergen, Svalbard. Polar Biol 32:1041–1046
Coulson SJ, Convey P, Aakra K, Aarvik L, Ávila-Jiménez ML, Babenko A, Biersma E, Boström S, Brittain J, Carlsson AM, Christoffersen KS, De Smet WH, Ekrem T, Fjellberg A, Füreder L, Gustafsson D, Gwiazdowicz DJ, Hansen LO, Holmstrup M, Kaczmarek L, Kolicka M, Kuklin V, Lakka HK, Lebedeva N, Makarova O, Maraldo K, Melekhina E, Ødegaard F, Pilskog HE, Simon JC, Sohlenius B, Solhøy T, Søli G, Stur E, Tanaevitch A, Taskaeva A, Velle G, Zawierucha K, Zmudczyńska-Skarbek K (2014) The terrestrial and freshwater invertebrate biodiversity of the archipelagoes of the Barents Sea; Svalbard, Franz Josef Land and Novaya Zemlya. Soil Biol Biochem 68:440–470
Cramp S, Perrins CM (1994) Buntings and new world warblers. In: Perrins CM (ed) Handbook of the birds of Europe, the Middle East and North Africa: the birds of the Western Palearctic, vol 9. Oxford University Press, Oxford, pp 118–138
Cyprich D, Krumpal M (1991) Incidence and taxonomy of Mioctenopsylla arctica arctica Rothschild 1922 (Siphonaptera) in the Spitsbergen. Biológia (Bratislava) 46:545–549
Elnitsky MA, Benoit JB, Denlinger DL, Lee RE (2008) Desiccation tolerance and drought acclimation in the Antarctic collembolan Cryptopygus antarcticus. J Insect Physiol 54:1432–1439. doi:10.1016/j.jinsphys.2008.08.004
Elton CS (1925) The dispersal of insects to spitsbergen. Trans Entomol Soc Lond 73:289–299
Fitze PS, Tschirren B, Richner H (2004) Life history and fitness consequences of ectoparasites. J Anim Ecol 73:216–226. doi:10.1111/j.0021-8790.2004.00799.x
Fjellberg A (1998) The Collembola of Fennoscandia and Denmark. Part I: Poduromorpha. Fauna Entomol Scand 35. Brill, Leiden
Fjellberg A. (2007) The Collembola of Fennoscandia and Denmark. Part II: Entomobryomorpha and Symphypleona. Fauna Entomol Scand 42. Brill, Leiden
Gwiazdowicz DJ, Błoszyk J, Bajerlein D, Halliday RB, Mizera T (2006) Mites (Acari: Mesostigmata) inhabiting nests of the white-tailed sea eagle Haliaeetus albicilla (L.) in Poland. Entomol Fenn 8:366–372
Gwiazdowicz DJ, Teodorowicz E, Coulson SJ (2011a) Redescription of Zercon solenites Haarløv, 1942 (Acari: Zerconidae) with a key to the Svalbard species of the genus Zercon. Int J Acarol 37:135–148
Gwiazdowicz DJ, Teodorowicz E, Coulson SJ (2011b) Redescription of Arctoseius haarlovi Lindquist, 1963 (Acari: Ascidae) from Spitsbergen. Entomol Fenn 22:140–148
Gwiazdowicz DJ, Coulson SJ, Grytnes JA, Pilskog HE (2012) The bird ectoparasite Dermanyssus hirundinis (Acari, Mesostigmata) in the High Arctic; a new parasitic mite to Spitsbergen, Svalbard. Acta Parasitol 57:378–384. doi:10.2478/s11686-012-0050-5
Hågvar S (1971) Some observations on Coleoptera, Hymenoptera and Siphonaptera in Svalbard 1968. Norsk Polarinstitutt Årbok 1969. Norwegian Polar Institute, Oslo, pp 101–106
Hågvar S (1975) Coleoptera in nests of birds of prey. Norw J Entomol 22:135–141
Harriman VB, Alisauskas RT (2010) Of fleas and geese: the impact of an increasing nest ectoparasite on reproductive success. J Avian Biol 41:573–579. doi:10.1111/j.1600-048X.2010.05013.x
Harrisson PM, Rothery P, Block W (1991) Drying processes in the Antarctic collembolan Cryptopygus antarcticus (Willem). J Insect Physiol 37:883–890
Hicks EA (1959) Check-list and bibliography on the occurrence of insects in birds’ nest. Iowa State College Press, Ames
Hicks EA (1962) Check-list and bibliography on the occurence of insects in birds’ nest: supplement 1. Iowa State Journal of Science, Ames
Hicks EA (1971) Check-list and bibliography on the occurence of insects in birds’ nest: supplement 2. Iowa State University Press, Ames
Hjelle A (1993) Geology of Svalbard. Norsk Polarinstitutt, Oslo
Hodkinson ID, Healey V, Coulson SJ (1994) Moisture relationships of the High Arctic collembolan Onychiurus arcticus. Physiol Entomol 19:109–114
Humphries DA (1968) The host-finding behaviour of the hen flea Ceratophyllus gallinae (Schrank) (Siphonaptera). Parasitology 58:403–414
Kovacs KM, Lydersen C (2006) Birds and mammals of Svalbard. Norwegian Polar Institute, Tromsø
Krištofík J, Mašán P, Šustek Z, Karaska D (2009) Arthropods in the nests of lesser spotted eagle (Aquila pomarina). Biologia 64:974–980
Krivolutsky AD, Lebedeva NV (2004a) Oribatid mites (Oribatei) in bird feathers: Passeriformes. Acta Zool Litu 14:19–38
Krivolutsky AD, Lebedeva NV (2004b) Oribatid mites (Oribatei, Acariformes) in bird feathers: non-passerines. Acta Zool Litu 14:26–47
Lebedeva NV, Krivolutsky DA (2003) Birds spread soil microarthropods to Arctic islands. Dokl Biol Sci 391:329–332
Lebedeva NV, Lebedev VD (2008) Transport of oribatid mites to the polar areas by birds. In: Bertrand M, Kreiter S, McCoy KD, Migeon A, Navajas M, Tixier M‐S, Vial L (eds). Integrative Acarology. Proceedings of the 6th European Congress. European Association of Acarologists, Vienna, pp 359–367
Lebedeva NV, Lebedev VD, Melekhina E (2006) New data on the oribatid mite (Oribatei) fauna of Svalbard. Dokl Biol Sci 407:182–186
Lepš J, Šmilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Lindquist EE, Krantz GW, Walter DE (2009) Classification. In: Krantz GW, Walter DE (eds) A manual of acarology. Tech University Press, Texas, pp 97–103
Majka CG, Klimaszewski J, Lauff RF (2006) New Coleoptera records from owl nests in Nova Scotia, Canada. Zootaxa 1194:33–47
Mehl R (1992) Fleas (Siphonaptera) from seabirds and their nests in mainland Norway and Spitzbergen. NIPH (National Institute of Public Health) Annals (Oslo) 15:3–15
Mehl R, Bang C, Kjos-Hanssen B, Lie H (1982) Mallophaga from Svalbard. Fauna Nor Ser B 29:19–23
Neubig JP, Smallwood JA (1999) The “significant others” of American kestrels: cohabitation with arthropods. Wilson Bull 111:269–271
Nielsen BO (1976) Sanitær entomologi. Zoologisk Laboratorium. Århus Universitet, Århus
Norton RA (1980) Observations on phoresy by oribatid mites (Acari: Oribatei). Int J Acarol 6:121–129
Norwegian Meteorological Institute (2013). Climate statistics for Svalbard. http://www.eklima.no. Accessed 1 Feb 2013
Norwegian Polar Institute (2014) Environmental Monitoring of Svalbard and Jan Mayen (MOSJ). http://mosj.npolar.no/en/index.html. Accessed 1 Feb 2014
O’Connor BM (2009) Cohort Astigmatina. In: Krantz GW, Walters DE (eds) A manual of acarology, 3rd edn. Texas Tech University Press, Texas, pp 565–657
Ohyama Y, Hiruta SI (1995) The terrestrial arthropods of Sør Rondane in eastern Dronning Maud Land, Antarctica, with biogeographical notes. Polar Biol 15:341–347
Oksanen J, Blanchet FG, Kindt R, Legendre P, O’hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2010) Vegan: community ecology package. R package version 1.17-4. http://CRAN.R-project.org/package=vegan. Accessed Feb 2011
Owen M, Black JM (2005) Barnacle goose Branta leucopsis. In: Kear J (ed) Ducks, geese and swans, vol I. Oxford University Press, Oxford, pp 329–334
Palmer M (2010) The ordination web page: ordination methods for ecologists. http://ordination.okstate.edu. Accessed 19 June 2011
Perotti AM, Braig HR (2009) Phoretic mites associated with animal and human decomposition. Exp Appl Acarol 49:85–124
Roy L, Chauve CM (2007) Historical review of the genus Dermanyssus Duges, 1834 (Acari: Mesostigmata: Dermanyssidae). Parasite 14:87–100
Seniczak S, Plichta W (1978) Structural dependence of moss-mites populations (Acari: Oribatei) on patchiness of vegetation in moss–lichen tundra at the north coast of Hornsund, West Spitsbergen. Pedobiologia 18:145–152
Shakhab S (2006) Oribatid mites (Oribatei, Acariformes) in nests of passerine birds. Entomol Rev 86:173–176
Sinclair BJ, Chown SL (2006) Caterpillars benefit from thermal ecosystem engineering by wandering albatrosses on sub-Antarctic Marion Island. Biol Lett 2:51–54
Southwood TRE, Henderson PA (2000) Ecological methods. Blackwell Science, Oxford
Subías LS, Shtanchaeva UY (2011) Listado sistemático de los ácaros oribátidos (Acari: Oribatida) iberocaucásicos. Rev Iber Aracnol 19:55–132
Tryjanowski P, Baraniak E, Bajaczyk R, Gwiazdowicz DJ, Konwerski S, Olszanowski Z, Szymkowiak P (2001) Arthropods in nests of the red-backed shrike (Lanius collurio) in Poland. Belg J Zool 131:69–74
Turienzo P, Di Iorio O (2008) Insects found in birds’ nests from Argentina: Anumbius annumbi (Vieillot, 1817) (Aves: Furnariidae). Zootaxa 1871:1–55
Weigmann G (2006) Hornmilben (Oribatida). Goecke and Evers, Keltern
Acknowledgments
This paper is dedicated to the memory of Torstein Solhøy who passed away during a late stage in its preparation. We thank the following who were instrumental in either enabling access to the nests or obtaining the samples, Børge Moe (Norwegian Institute for Nature Research), Maarten J. J. E. Loonen and Thomas Oudman (University of Groningen), Geir Wing Gabrielsen, Erlend Lorentzen and Kjetil Sagerup (Norwegian Polar Institute), Elin Noreen, Anja Johansen Haugerud and Siv Kristoffersen (Norwegian University of Science and Technology). Thanks to Arne Moksnes and Claus Bech (Norwegian University of Science and Technology, NTNU) for granting access to the snow bunting nesting boxes. Thanks also to the following taxonomists who assisted in identifications: Arne Fjellberg (Arne Fjellberg Entomological Research), Gaute Velle (University of Bergen), Robert Lewis (Iowa State University), Petter Jordan (University Museum of Bergen) and Valerie Behan-Pelletier (Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa). Prof. Anna Sjöblom for assistance drawing Fig. 1. We also thank Sigmund Hågvar, Anatoly B. Babenko and Melodie McGeoch for valuable comments on the manuscript. The project was funded by the University Centre in Svalbard, the Arctic Field Grant and the L. Meltzers Høyskolefond, University of Bergen.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pilskog, H.E., Solhøy, T., Gwiazdowicz, D.J. et al. Invertebrate communities inhabiting nests of migrating passerine, wild fowl and sea birds breeding in the High Arctic, Svalbard. Polar Biol 37, 981–998 (2014). https://doi.org/10.1007/s00300-014-1495-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-014-1495-9