
Abstract
The posterior parietal cortex (PPC) is strongly related to task performance by evaluating sensory cues and visually guided movements. Sensorimotor processing is improved by task repetition as indicated by reduced response time. We investigated practice-induced changes in PPC visuomotor processing during a Go/NoGo task in humans using 306-channel magnetoencephalography. Eleven healthy adult males were instructed to extend the right index finger when presented with the Go stimulus (a red circle), but not to react to the NoGo stimulus (a green circle or a red square). Magnetic fields over the visual, posterior parietal, and sensorimotor cortices were measured before and after 3 days of task practice. The first peak of the visual-evoked field (VEF) occurred at approximately 80 ms after presentation of either the Go or NoGo stimulus, while a PPC response, with latency to a peak of 175.8 ± 26.7 ms, occurred only after the Go stimulus. No significant change in the first peak of VEF was measured after 3 days of task practice, but there was a significant reduction in the latency to peak PPC activity (160.1 ± 27.6 ms) and in the time from peak PPC activity to electromyogram onset. In all participants, practice resulted in a significant reduction in reaction time. These results demonstrate that practicing a sensorimotor task induces neuroplastic changes in PPC that accelerate sensorimotor processing and reduce motor response times.
Similar content being viewed by others


Introduction
Motor reaction tasks that require articular movements in response to a sensory stimulus are used extensively to assess sensorimotor and cognitive functions. The Go/NoGo task adopted in this study can measure reaction time and requires a response to a specific “Go” stimulus, but no response to the other “NoGo” stimulus. Moreover, coupling these Go/NoGo tasks with modern neuroimaging or electrophysiological methods can reveal the aspects of cortical sensorimotor processing (Hoshiyama et al. 1996; Kida et al. 2006; Nakata et al. 2006; Stinear et al. 2009).
Reaction times are a reliable measure of improved sensorimotor task performance and can be divided into premotor time (PMT) from visual presentation to electromyogram (EMG) onset and the electromechanical time from EMG onset to movement onset. It is believed that PMT reflects the time required for processing of sensory stimuli, selection of the appropriate response, motor planning, and transmission of motor signals from the primary motor area (M1) to articular muscles via alpha motor neurons of the spinal cord. Visuomotor integration and control are a central function of the posterior parietal cortex (PPC) (Iacoboni 2006). Previous studies in primates measuring simple motor responses to sensory stimuli reported PPC and premotor cortex (PMC) activity during the time between the stimulus and motor execution (Kurata and Wise 1988; Burnod et al. 1992; Sakata et al. 1995). PPC projects strongly to PMC and is believed to process sensory information, select appropriate responses, and transmit this information to PMC (Mushiake et al. 1991; Kalaska and Crammond 1995; Sakata et al. 1995; Andersen et al. 1997; Snyder et al. 1997). Studies with human participants have also reported that PPC is critical for sensorimotor tasks (Kertzman et al. 1997; Toni et al. 1999; Beurze et al. 2010). PPC may function in “transforming” visual information into motor plans (Buneo and Andersen 2006; Stocco et al. 2012). Improved Go/NoGo task performance is associated with changes in cortical processing that are reflected by both reduced response times and spatiotemporal changes in cortical activity. Presumably, improvements in task performance due to practice would be reflected by changes in activity within PPC, but this has not been demonstrated.
Well-practiced movements are performed more accurately and quickly than novel movements, presumably because of reduced cortical processing times within perceptual and motor systems (Kida et al. 2005). Yotani et al. (2011) reported a significant reduction in the time between visual stimulus presentation and M1 activity following 8 weeks of task practice. However, many previous studies using reaction time as the main output measure have not clarified reductions due to cortical processing following practice.
The purpose of this study was to investigate the changes in cortical information processing during a visual reaction task in humans using high spatiotemporal resolution 306-channel magnetoencephalography (MEG) and to investigate the effects of practice on cortical information processing. We hypothesized that activity in the contralateral PPC during the time from visual stimulus presentation to movement onset would be altered by practice. Specifically, we speculated that the time to peak PPC activity after stimulus presentation and the time from peak PPC activity to the initiation of movement would be reduced by practice. In contrast, the early sensory processing events, as reflected by the first peak of the visual-evoked field (VEF) in the occipital cortex, and the peripheral motor response, reflected by the time between EMG onset and movement onset, should be unchanged.
Methods
Participants
The visuomotor Go/NoGo task was performed by 11 healthy male volunteers (mean ± SD, 23.0 ± 2.7 years of age; range, 20–27 years) who gave informed written consent. The study was approved by the ethics committee of Niigata University of Health and Welfare and conformed to The Code of Ethics of the World Medical Association (Declaration of Helsinki).
Go/NoGo task
Cortical visuomotor processing was assessed during a Go/NoGo task. Participants were required to extend the index finger of the right hand approximately 3 cm above the horizontal surface as quickly as possible in response to the Go stimulus (a red circle) and to remain motionless in response to the NoGo stimulus (a green circle or a red square). The duration of visual stimulus presentation was 100 ms, and the interval between successive visual stimuli was 4,000 ms. The width of the visual stimulus was approximately 10 cm. The order of Go and NoGo stimuli was randomized and included 45 “Go” trials and 90 “NoGo” trials (a green circle 45 stimuli and a red square 45 stimuli) per session. We confirmed that no EMG activity was observed in response to the NoGo stimulus.
The method for recording movement-related cerebral fields (MRCFs) has been described in detail elsewhere (Kato et al. 2006; Onishi et al. 2006, 2011). Each participant’s index finger was placed on a small plate with a light-emitting diode (LED) sensor. When the fingertip was detached from the plate by index finger extension, the LED signal was disrupted. In this study, visual stimulus onset and movement onset were used to trigger MRCF acquisition. Using a projector positioned outside the magnetically shielded room, visual stimuli were projected on to a screen placed in front of the participants at a distance of 1 m.
Median nerve stimulation
Sensory-evoked fields (SEFs) were measured to establish a standard cortical reference location. In brief, the participant’s right median nerve was electrically stimulated at the wrist at an intensity of 1.2 times that of the motor threshold using 0.2-ms monophasic square wave pulses at 1.5 Hz. The mean intensity for SEF was 5.9 mA (range, 4.5–9.6 mA). The equivalent current dipole (ECD) of the first peak response occurring approximately 20 ms after median nerve stimulation (N20m) was used to locate the median nerve field of the somatosensory cortex.
EMG
Electromyogram (EMG) from the right extensor indicis muscle was recorded using a surface electrode and the signal was filtered at 20-Hz high-pass. We calculated the times from visual stimulus onset to the rectified EMG onset and from the rectified EMG onset to movement onset. The point at which the rectified EMG exceeded two standard deviations above baseline was considered as EMG onset. Based on previous studies (Endo et al. 1999; Kida et al. 2005) and our own pilot experiment, trials with reaction times shorter than 100 ms and longer than 400 ms were excluded from the analysis.
Practice protocol
We first recorded MRCFs and reaction times at baseline, and participants then practiced the Go/NoGo task under the same experimental conditions. Practice consisted of three sessions per day for 3 days, with each session consisting of 45 Go trials randomly interspersed among “NoGo” trials. We then recorded MRCFs and reaction times again using the same methodology and stimulus conditions.
Data acquisition
Neuromagnetic signals were recorded using a 306-channel whole-head MEG system (Vectorview; Elekta, Helsinki, Finland). This 306-channel device contains 102 identical triple sensors, each housing two orthogonal planar gradiometers and one magnetometer. In this study, we analyzed MEG signals recorded from 204 planar-type gradiometers. This configuration of gradiometers specifically detects the signal just above the source current. Continuous MEG signals were sampled at 1,000 Hz using a band-pass filter ranging between 0.03 and 330 Hz. The participants were comfortably seated inside a magnetically shielded room (Tokin Ltd., Sendai, Japan). MEG recordings were acquired from 1,000 before to 1,500 ms after visual stimulus onset for analyzing visuomotor brain activity and from 1,500 before to 1,000 ms after movement onset for analyzing movement-evoked fields 1 (MEF1). The average of 45 Go trial records was obtained during each session. Before MEG measurements, three anatomical fiducial points (nasion and bilateral preauricular points) and four indicator coil locations on the scalp were digitized using a three-dimensional digitizer (Polhemus, Colchester, VT, USA). The fiducial points provided the spatial information necessary for the integration of magnetic resonance (MR) images and MEG data, while the indicator coils determined the position of the participant’s head in relation to the helmet. T1-weighted MR images were obtained using a 1.5-T system (Signa HD; GE Healthcare, Milwaukee, WI, USA).
Data analysis
For analysis of MEGs, the band-pass filter was set to 0.5 Hz and 60 Hz. The first 200 ms was used as the baseline, the epoch from −1,000 to −800 ms before visual stimulus onset for analyzing visuomotor brain activity, and the epoch from −1,500 to −1,300 ms before movement onset for analyzing MEF1. If visual stimulus onset was used as a trigger, MEF1 was not clearly recorded because the time from visual onset to movement onset varied widely. Therefore, movement onset was used as a trigger for analyzing MEF1. For SEF analysis, the band-pass filter was set to 0.5 and 100 Hz, and the 20-ms period preceding stimulation was used as the baseline.
The source components of interest for MEGs were estimated as ECDs using a least-squares search within a subset of 22–26 channels over the peak response area. We used source modeling software (Elekta) to estimate the sources. The ECD locations were calculated using the fiducial points (nasion and bilateral preauricular points). We calculated ECDs from 50 to 400 ms for each 1 ms and accepted ECDs corresponding with a peak amplitude from sensor levels and a goodness-of-fit (g) of >80% for analysis.
Data are expressed as mean ± standard deviation. Paired t tests were used to evaluate the statistical significance of pre-practice versus post-practice differences in EMG onset, movement onset, the latency to peak PPC activity, and the peak latencies and strengths of the source activity for VEF, PPC, and MEF1 during the Go task. Two-way repeated ANOVA was performed to compare differences in VEF between the task conditions (Go vs. NoGo) and practice conditions (pre-practice vs. post-practice). Differences between source localizations were assessed by the Friedman test with post hoc Wilcoxon rank tests. The significance level was set at 0.05 for all tests.
Results
Superimposed over the scalp at recording positions are MEG waveforms measured in response to the Go task (solid lines) and the NoGo task (dotted lines) prior to practice for 1 participant (Fig. 1). Prominent peaks at 95.0 ms in response to the Go stimulus and 96.8 ms in response to the NoGo stimulus were observed over the occipital cortex (Fig. 1a). A peak at 190.6 ms was measured over the vertex in response to the Go stimulus (Fig. 1b), but not the NoGo stimulus. The peak MEF1 was observed 345.4 ms after the Go stimulus at Fig. 1c. Table 1 shows the each time for all participants at pre- and post-practice in Go stimulus. We have used the largest amplitude on each waveform within the sensors of interest in VEF over the occipital area, PPC over the parietal area, and MEF1 over the central area with each subset of 12–14 channels. Source modeling used in this study takes into account peak measurements on waveforms for analysis.

The typical waveforms and whole-scalp magnetic response to Go and NoGo stimuli. Superimposed response from individual channels during the Go task (solid lines) and the NoGo task (dotted lines) prior to task practice (data from Participant 10). Typical waveforms starting from 1,000 ms before to 1,000 ms after visual stimulus presentation in response to the Go task (solid line) or NoGo task (dotted lines) are enlarged at (a), (b), and (c). Over the occipital cortex (a), prominent peaks were observed in both the Go and NoGo conditions. Over the vertex (b), a prominent peak was observed only after the Go stimulus (at 190.6 ms). At c, MEF1 was observed immediately after movement onset and at 345.4 ms after the Go stimulus
The first peak of VEF after visual stimulus onset did not differ significantly between pre- and post-practice conditions for the Go task (80.8 ± 17.2 ms vs. 80.5 ± 12.8 ms) or the NoGo task (78.5 ± 15.8 ms vs. 79.5 ± 12.5 ms). Two-way ANOVA revealed no significant main effect of the task (F = 0.462, p = 0.512) or practice condition (F = 0.017, p = 0.899) and no significant task × condition interaction (F = 0.029, p = 0.869). In contrast to the first peak of VEF, the latency to peak PPC activity after visual stimulus onset in the Go task condition was significantly shorter following practice (p < 0.05) (Table 1). The latency from movement onset to MEF1 was not significantly changed following practice (pre, 35.9 ± 3.7 ms; post, 34.5 ± 3.6 ms, p = 0.345).
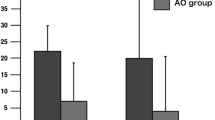
Equivalent current dipoles (ECDs) at peak VEF were located over the occipital cortex for both task conditions (Go and NoGo) and both practice conditions (pre-practice and post-practice). However, ECD estimated at peak PPC had a g value of >80 % only for the Go task. The mean ECD locations for all participants at peak PPC activity and at peak MEF1 for both pre- and post-practice in Go tasks were calculated relative to that of N20m (Fig. 2). The mean ECD locations at peak PPC activity during Go tasks were significantly medial (pre, 15.4 ± 7.8 mm; post, 13.2 ± 14.4 mm) and posterior (pre, −23.8 ± 7.9 mm; post, −21.2 ± 14.3 mm) to that of N20m, but not significantly superior to that of N20m. The mean ECD location of MEF1 was significantly medial (pre, 6.1 ± 3.8 mm; post, 7.2 ± 4.8 mm) to that of N20m, but not significantly superior to those of N20m at pre- and post-practice. The ECD locations at peak PPC and MEF1 for 1 participant (Participant 3) after practice are superimposed on that participant’s MR images (Fig. 3). All other participants exhibited ECD locations close to those of this participant, with g values of >80% for the Go task when visual stimulus onset was used to analyze PPC activity.

The mean ECD locations for PPC activity and MEF1 relative to that of N20m. Average ECD locations (for all 11 participants) at peak MEF1 pre-practice (diamond) and post-practice (square) and at peak PPC activity pre-practice (multiplication) and post-practice (circle) relative to that at N20m (triangle) on horizontal and frontal planes in Go task. The error bars indicate standard deviations. ECD at MEF1 was significantly medial to N20m for both pre- and post-practice conditions (pre, 6.1 ± 3.6 mm; post, 7.2 ± 4.8 mm). Both pre- and post-practice ECDs at peak PPC were significantly medial (pre, 15.4 ± 7.8 mm; post, 13.2 ± 14.4 mm) and significantly posterior (pre, −23.8 ± 7.9 mm; post, −21.2 ± 14.3 mm) to the ECD at N20m. ECDs at peak MEF1 and peak PPC activity were not significantly superior to those at N20m for either pre- or post-practice conditions

The ECD locations are superimposed on the subject’s MR images. Result of ECD analysis in Participant 3 following practice in Go task. The locations of ECDs are superimposed on the participant’s MR images. a Estimated ECD locations at peak PPC (circle), MEF1 (square), and N20m (triangle) in the horizontal plane. b Estimated ECD location at peak PPC activity in the frontal and sagittal planes. c Estimated ECD locations at MEF1 and at N20m in the frontal and sagittal planes
The peak latencies and strengths of the source activity for VEF were 81.0 ± 16 ms (15.3 ± 2.1 nAm) in pre-practice and 81.1 ± 14.7 ms (14.7 ± 2.2 nAm) in post-practice after the Go stimulus, and the peak latencies and strengths for VEF did not change significantly following practice. These for PPC were 175.8 ± 29.1 ms (23.3 ± 8.7 nAm) in pre-practice and 159.9 ± 28.2 ms (21.4 ± 8.9 nAm) in post-practice. The peak latency of the source activity for PPC after the Go stimulus changed significantly following practice, and the strength of the source activity was not significant. These for MEF1 were 34.6 ± 4.0 ms (29.6 ± 6.4 nAm) in pre-practice and 34.2 ± 4.0 ms (28.5 ± 7.2 nAm) in post-practice after movement onset, and the peak latencies and strengths for MEF1 did not change significantly following practice.
The times from visual stimulus onset to EMG onset and movement onset were all significantly shorter following practice (p < 0.05) (Table 2). Moreover, the times at sensor levels from the first peak of VEF to peak PPC activity and from peak PPC activity to EMG onset were significantly shorter following practice (p < 0.05), while no difference was observed in the time between EMG onset and movement onset (p = 0.478).
Discussion
Posterior parietal cortex (PPC) is critical for visuomotor task performance. In the present study, we observed PPC activity only during the Go condition of the Go/NoGo task, implicating PPC in response selection. Moreover, the total response time, the latency to peak PPC activity, and the time from peak PPC to EMG onset were significantly reduced following task practice, suggesting that practice induces neuroplastic changes in PPC that accelerate sensory evaluation, response selection, and speed of transmission from PPC to M1.
Neural activity was measured from the visual cortex in response to both Go and NoGo visual stimuli. In previous studies, the time from visual stimulus presentation until peak visual cortex activity ranged from 37 to 120 ms (Endo et al. 1999; Moradi et al. 2003; Inui et al. 2006). In the present study, the first peak of VEF was observed approximately 80 ms after presentation under both Go and NoGo task conditions and was not altered by practice. Thus, the early stages of visual processing from the retina to the visual cortex are likely not influenced by the behavioral salience of the stimulus and practice.
The posterior portion of PPC is adjacent to the occipital visual area, and these two regions are strongly connected (Petrides and Pandya 1984; Felleman and Van Essen 1991; Cavada 2001). Furthermore, PPC is connected to PMC. Based on these anatomical connections, it is believed that PPC may function in “transforming” visual information into motor plans (Buneo and Andersen 2006; Stocco et al. 2012). Indeed, PPC activation is observed during the interval between visual stimulus presentation and motor response (Deiber et al. 1996; Andersen et al. 1997; Desmurget et al. 1999; Creem-Regehr 2009) and during visually guided movements (Sasaki and Gemba 1991; Amino et al. 2001). The previous study reported that PPC showed strong activity when participants responded with a finger key press to the visual stimulus and that PPC played a key role in visuomotor transfer (Iacoboni and Zaidel 2004). Shibata and Ioannides (2001) measured cortical activity during a Go/NoGo task and reported prominent activity in the contralateral PPC only after the Go stimulus, in accordance with our results and supporting a role for PPC in selecting appropriate motor responses to visual cues.
After 3 days of practice, there was a significant reduction in the time from visual stimulus presentation to EMG onset for all participants. This time reflects transmission of neural signals from the retina to the primary visual cortex via the optic nerve and lateral geniculate nucleus, from visual cortices to PPC (Kalaska and Crammond 1995; Iacoboni 2006), and from PPC to PMC and M1 (Nishitani et al. 1999), and from M1 to the muscle via alpha motor neurons of the spinal cord. In the present study, the time from stimulus onset to peak visual field activity was unchanged by practice, while there was an approximately 15 ms reduction in the latency to peak PPC activity following 3 days of task repetition, strongly suggesting that pathways from the visual cortex to PPC are potentiated by practice. Kida et al. (2005) compared the Go/NoGo reaction times of baseball players to sedentary adult males and found that reaction times were significantly shorter in the athletes. They attributed this acceleration to cortical processes, such as movement selection based on sensory discrimination. Hihara et al. (2006) investigated PPC activity in monkeys trained for 3 weeks to use tools and reported post-training changes in the characteristic PPC responses to visual information (Hihara et al. 2006). Thus, both human and animal studies suggest that practice leads to neuroplastic changes in PPC. In this study, these changes are manifested by a reduced latency from visual stimulus onset to peak PPC activity.
We also observed a reduction in the latency from peak PPC activity to EMG onset following practice, suggesting that the contralateral M1 was facilitated and changes occur in intracortical networks (Schlaug et al. 1994; Karni et al. 1995; Pascual-Leone et al. 1995; Hayashi et al. 2002). It has been reported that there are no changes in the latency between M1 activity and initiation of muscle activity following practice (Yotani et al. 2011). Hence, these post-practice reductions in the latency suggest that processing within PPC and transmission from PPC to M1 are accelerated by task practice.
The Go/NoGo task used in the present study involved a simple index finger extension movement. Nevertheless, even such simple tasks require PPC and are improved by practice, likely by increasing processing speed within PPC and through the PPC–PMC–M1 pathway. However, we could not determine the presence of another pathway (not through the PPC) that contributed during the Go/No task in this study. We cannot rule out that the other pathway may be related to the visuomotor task. Further investigations are required to gain more insights into other pathways that are related to the visuomotor task. It is likely that complex tasks are relatively more dependent on PPC processing. Additional studies using more complex tasks are clearly warranted to investigate the contribution of PPC neuroplasticity and facilitation of PPC–PMC–M1 transmission to improvements in task performance following practice.
References
Amino Y, Kyuhou S, Matsuzaki R, Gemba H (2001) Cerebello-thalamo-cortical projections to the posterior parietal cortex in the macaque monkey. Neurosci Lett 309:29–32
Andersen RA, Snyder LH, Bradley DC, Xing J (1997) Multimodal representation of space in the posterior parietal cortex and its use in planning movements. Annu Rev Neurosci 20:303–330. doi:10.1146/annurev.neuro.20.1.303
Beurze SM, Toni I, Pisella L, Medendorp WP (2010) Reference frames for reach planning in human parietofrontal cortex. J Neurophysiol 104:1736–1745. doi:10.1152/jn.01044.2009
Buneo CA, Andersen RA (2006) The posterior parietal cortex: sensorimotor interface for the planning and online control of visually guided movements. Neuropsychologia 44:2594–2606. doi:10.1016/j.neuropsychologia.2005.10.011
Burnod Y, Grandguillaume P, Otto I, Ferraina S, Johnson PB, Caminiti R (1992) Visuomotor transformations underlying arm movements toward visual targets: a neural network model of cerebral cortical operations. J Neurosci 12:1435–1453
Cavada C (2001) The visual parietal areas in the macaque monkey: current structural knowledge and ignorance. Neuroimage 14:S21–S26. doi:10.1006/nimg.2001.0818
Creem-Regehr SH (2009) Sensory-motor and cognitive functions of the human posterior parietal cortex involved in manual actions. Neurobiol Learn Mem 91:166–171. doi:10.1016/j.nlm.2008.10.004
Deiber MP, Ibanez V, Sadato N, Hallett M (1996) Cerebral structures participating in motor preparation in humans: a positron emission tomography study. J Neurophysiol 75:233–247
Desmurget M, Epstein CM, Turner RS, Prablanc C, Alexander GE, Grafton ST (1999) Role of the posterior parietal cortex in updating reaching movements to a visual target. Nat Neurosci 2:563–567. doi:10.1038/9219
Endo H, Kizuka T, Masuda T, Takeda T (1999) Automatic activation in the human primary motor cortex synchronized with movement preparation. Brain Res Cogn Brain Res 8:229–239
Felleman DJ, Van Essen DC (1991) Distributed hierarchical processing in the primate cerebral cortex. Cereb Cortex 1:1–47
Hayashi S, Hasegawa Y, Kasai T (2002) Transcranial magnetic stimulation study of plastic changes of human motor cortex after repetitive simple muscle contractions. Percept Mot Skills 95:699–705
Hihara S, Notoya T, Tanaka M et al (2006) Extension of corticocortical afferents into the anterior bank of the intraparietal sulcus by tool-use training in adult monkeys. Neuropsychologia 44:2636–2646. doi:10.1016/j.neuropsychologia.2005.11.020
Hoshiyama M, Koyama S, Kitamura Y, Shimojo M, Watanabe S, Kakigi R (1996) Effects of judgement process on motor evoked potentials in Go/No-go hand movement task. Neurosci Res 24:427–430
Iacoboni M (2006) Visuo-motor integration and control in the human posterior parietal cortex: evidence from TMS and fMRI. Neuropsychologia 44:2691–2699. doi:10.1016/j.neuropsychologia.2006.04.029
Iacoboni M, Zaidel E (2004) Interhemispheric visuo-motor integration in humans: the role of the superior parietal cortex. Neuropsychologia 42:419–425
Inui K, Sannan H, Miki K, Kaneoke Y, Kakigi R (2006) Timing of early activity in the visual cortex as revealed by simultaneous MEG and ERG recordings. Neuroimage 30:239–244. doi:10.1016/j.neuroimage.2005.09.003
Kalaska JF, Crammond DJ (1995) Deciding not to GO: neuronal correlates of response selection in a GO/NOGO task in primate premotor and parietal cortex. Cereb Cortex 5:410–428
Karni A, Meyer G, Jezzard P, Adams MM, Turner R, Ungerleider LG (1995) Functional MRI evidence for adult motor cortex plasticity during motor skill learning. Nature 377:155–158. doi:10.1038/377155a0
Kato Y, Endo H, Kizuka T, Asami T (2006) Automatic and imperative motor activations in stimulus-response compatibility: magnetoencephalographic analysis of upper and lower limbs. Exp Brain Res 168:51–61. doi:10.1007/s00221-005-0090-2
Kertzman C, Schwarz U, Zeffiro TA, Hallett M (1997) The role of posterior parietal cortex in visually guided reaching movements in humans. Exp Brain Res 114:170–183
Kida N, Oda S, Matsumura M (2005) Intensive baseball practice improves the Go/Nogo reaction time, but not the simple reaction time. Brain Res Cogn Brain Res 22:257–264. doi:10.1016/j.cogbrainres.2004.09.003
Kida T, Wasaka T, Nakata H, Kakigi R (2006) Centrifugal regulation of task-relevant somatosensory signals to trigger a voluntary movement. Exp Brain Res 169:289–301. doi:10.1007/s00221-005-0141-8
Kurata K, Wise SP (1988) Premotor cortex of rhesus monkeys: set-related activity during two conditional motor tasks. Exp Brain Res 69:327–343
Moradi F, Liu LC, Cheng K, Waggoner RA, Tanaka K, Ioannides AA (2003) Consistent and precise localization of brain activity in human primary visual cortex by MEG and fMRI. Neuroimage 18:595–609
Mushiake H, Inase M, Tanji J (1991) Neuronal activity in the primate premotor, supplementary, and precentral motor cortex during visually guided and internally determined sequential movements. J Neurophysiol 66:705–718
Nakata H, Inui K, Wasaka T, Tamura Y, Akatsuka K, Kida T, Kakigi R (2006) Higher anticipated force required a stronger inhibitory process in go/nogo tasks. Clin Neurophysiol 117:1669–1676. doi:10.1016/j.clinph.2006.03.032
Nishitani N, Uutela K, Shibasaki H, Hari R (1999) Cortical visuomotor integration during eye pursuit and eye-finger pursuit. J Neurosci 19:2647–2657
Onishi H, Soma T, Kameyama S et al (2006) Cortical neuromagnetic activation accompanying two types of voluntary finger extension. Brain Res 1123:112–118. doi:10.1016/j.brainres.2006.09.033
Onishi H, Oyama M, Soma T, Kirimoto H, Sugawara K, Murakami H, Kameyama S (2011) Muscle-afferent projection to the sensorimotor cortex after voluntary movement and motor-point stimulation: an MEG study. Clin Neurophysiol 122:605–610. doi:10.1016/j.clinph.2010.07.027
Pascual-Leone A, Nguyet D, Cohen LG, Brasil-Neto JP, Cammarota A, Hallett M (1995) Modulation of muscle responses evoked by transcranial magnetic stimulation during the acquisition of new fine motor skills. J Neurophysiol 74:1037–1045
Petrides M, Pandya DN (1984) Projections to the frontal cortex from the posterior parietal region in the rhesus monkey. J Comp Neurol 228:105–116. doi:10.1002/cne.902280110
Sakata H, Taira M, Murata A, Mine S (1995) Neural mechanisms of visual guidance of hand action in the parietal cortex of the monkey. Cereb Cortex 5:429–438
Sasaki K, Gemba H (1991) Cortical potentials associated with voluntary movements in monkeys. Electroencephalogr Clin Neurophysiol Suppl 42:80–96
Schlaug G, Knorr U, Seitz R (1994) Inter-subject variability of cerebral activations in acquiring a motor skill: a study with positron emission tomography. Exp Brain Res 98:523–534
Shibata T, Ioannides AA (2001) Contribution of the human superior parietal lobule to spatial selection process: an MEG study. Brain Res 897:164–168
Snyder LH, Batista AP, Andersen RA (1997) Coding of intention in the posterior parietal cortex. Nature 386:167–170. doi:10.1038/386167a0
Stinear CM, Coxon JP, Byblow WD (2009) Primary motor cortex and movement prevention: where Stop meets Go. Neurosci Biobehav Rev 33:662–673. doi:10.1016/j.neubiorev.2008.08.013
Stocco A, Lebiere C, O’Reilly RC, Anderson JR (2012) Distinct contributions of the caudate nucleus, rostral prefrontal cortex, and parietal cortex to the execution of instructed tasks. Cogn Affect Behav Neurosci 12(4):611–628
Toni I, Schluter ND, Josephs O, Friston K, Passingham RE (1999) Signal-, set- and movement-related activity in the human brain: an event-related fMRI study. Cereb Cortex 9:35–49
Yotani K, Tamaki H, Yuki A, Kirimoto H, Kitada K, Ogita F, Takekura H (2011) Response training shortens visuo-motor related time in athletes. Int J Sports Med 32:586–590. doi:10.1055/s-0031-1275299
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sugawara, K., Onishi, H., Yamashiro, K. et al. Repeated practice of a Go/NoGo visuomotor task induces neuroplastic change in the human posterior parietal cortex: an MEG study. Exp Brain Res 226, 495–502 (2013). https://doi.org/10.1007/s00221-013-3461-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-013-3461-0