
Abstract
Rationale
The endocannabinoid and endovanniloid anandamide (AEA) exerts biphasic effects when injected into the dorsolateral periaqueductal grey (dlPAG) in rats submitted to threatening situations. Whereas lower doses of AEA induce anxiolytic-like effects by activating cannabinoid CB1 receptors, no effects are observed with higher doses, possibly due to the simultaneous activation of transient receptor potential vanilloid type 1 (TRPV1) receptors. This activation would facilitate glutamatergic neurotransmission.
Objective
Considering that the blockade of TRPV1 or NMDA receptors in the dlPAG induces anxiolytic-like effects, we tested the hypothesis that facilitation of glutamate transmission through TRPV1 is responsible for the lack of anxiolytic-like effect observed with high AEA doses.
Methods
Male Wistar rats with a unilateral cannula aimed at the dlPAG received injections of an ineffective dose of AP7 (an NMDA antagonist, 1 nmol) or capsazepine (CPZ, a TRPV1 antagonist, 10 nmol), followed by a high dose of AEA (50 and 200 pmol) and were exposed to the elevated plus maze (EPM) or the Vogel conflict test (VCT).
Results
AP7, CPZ, or AEA did not induce any significant effects when administered alone. However, AP7 or CPZ prior to AEA significantly increased the percentage of entries and time spent in the open arms of EPM and the number of punished licks in the VCT suggesting an anxiolytic-like effect.
Conclusions
These results suggest that the lack of anxiolytic-like effect of higher AEA doses is due to facilitation of glutamate release in the dlPAG, probably via activation of TRPV1 receptors in this structure.
Similar content being viewed by others
Introduction
The dorsolateral periaqueductal grey (dlPAG) is responsible for elaborating active defensive responses displayed by animals in dangerous situations (Bandler et al. 2000; Carrive 1993). Under natural conditions, these responses are probably mediated by local glutamate release (Guimaraes et al. 2005). Accordingly, intra-dlPAG administration of glutamate ionotropic receptor (NMDA or non-NMDA) agonists induces escape reactions (Aguiar et al. 2006; Bittencourt et al. 2004), whereas the blockade of these receptors causes anxiolytic-like and anti-aversive effects in rodents (Aguiar and Guimaraes 2009; Guimaraes et al. 1991; Matheus and Guimaraes 1997; Molchanov and Guimaraes 2002).
In addition to glutamate, other neurotransmitters have also been shown to modulate anxiety-like behaviors in the dlPAG, including GABA, nitric oxide (NO), endocannabinoids and, more recently, endovanilloids (Casarotto et al. 2012; Finn et al. 2003; McGregor et al. 2004; Moreira et al. 2007; Terzian et al. 2009). Endovanilloids are putative endogenous agonists of the transient receptor potential vanilloid type 1 (TRPV1), a nonselective cation channel (Caterina et al. 1997; Clapham et al. 2005). The first endovanilloid described was anandamide (N-arachidonoylethanolamine, AEA), an arachidonic acid-derived compound, which is also an endocannabinoid and an agonist of cannabinoid receptor type 1 (CB1) (Di Marzo et al. 2001; Smart and Jerman 2000; Zygmunt et al. 2000). Although AEA is the endogenous ligand of both cannabinoid and vanilloid system, its affinity for TRPV1 is lower than that needed to activate CB1 receptors (Ross 2003).
Several results have suggested that endocannabinoids and endovanilloids modulate emotional states in opposite ways (for review see Moreira et al. 2012; Viveros et al. 2007). In line with these pieces of evidence, previous studies from our group showed that lower doses of AEA injected into the dlPAG caused anxiolytic-like effects in distinct animal models of anxiety. These effects were blocked by previous treatment with AM251, a CB1 receptor antagonist (Moreira et al. 2007). AEA, however, induced a bell-shaped dose–response curve, with higher doses being ineffective (Moreira et al. 2007).
Studies employing PAG slices have shown that activation of CB1 receptors inhibits GABAergic and glutamatergic synaptic transmission (Vaughan et al. 2000), which may be related to the biphasic effects on anxiety-like responses observed with CB1 receptor agonists in the PAG (Moreira et al. 2007). However, PAG neurons also express TRPV1 receptors (Cavanaugh et al. 2011; Cristino et al. 2006; Maione et al. 2006; Toth et al. 2005) and, similar to glutamate, TRPV1 activation has been suggested to facilitate anxiety-like responses (Marsch et al. 2007; Rubino et al. 2008; Terzian et al. 2009; Toth et al. 2005). In the central nervous system (CNS), TRPV1 receptors are proposed to modulate neuronal activity mainly through potentiation of glutamatergic signaling (Kawahara et al. 2011; McGaraughty et al. 2003; Starowicz et al. 2007; Xing and Li 2007), an effect that has also been shown in PAG (Kawahara et al. 2011; Palazzo et al. 2002; Starowicz et al. 2007).
Since high doses of cannabinoids such as AEA, ACEA, and WIN 55,212-2 can also activate TRPV1 receptors (Casarotto et al. 2012; Di Marzo et al. 2001; Fogaca et al. 2012; Pertwee 2006), we hypothesized that the bell-shaped dose–response curves observed in anxiety models after administration of AEA in the dlPAG, with higher doses being ineffective, depend on increased glutamate release via activation of TRPV1 receptors.
Materials and methods
Subjects
Male Wistar rats were obtained from the colony of rats maintained by the Campus of the University of São Paulo-Ribeirão Preto. The rats weighing 220–240 g at the beginning of each experiment were housed in groups of four in a temperature-controlled room (24 ± 1 °C) under standard laboratory conditions with free access to food and water and a 12-h light/12-h dark cycle (lights on at 6:30 a.m.). All the experiments were conducted between 8 a.m. and 12 a.m. To avoid interference from different batches of animals, each experiment always included control and treated rats. Procedures were conformed within the Brazilian Society of Neuroscience and Behavior guidelines for the care and use of laboratory animals. The experimental protocols were approved by the Brazilian College of Animal Experimentation and by the School of Medicine of Ribeirao Preto of the University of Sao Paulo Ethical Commission of Ethics in Animal Research (CETEA; no. 083/2008).
Drugs
The CB1/TRPV1 agonist anandamide (AEA) (50 and 200 pmol/0.2 μL; Tocris, USA) was dissolved in TocrisolveTM® 100 (a solvent that contains a 1:4 ratio of soya oil/water, emulsified with the block co-polymer Pluronic F68). The TRPV1 antagonist capsazepine (CPZ; 10 nmol/0.2 μL; Tocris, USA) was dissolved in dimethyl sulfoxide (DMSO) 100 %. The glutamate NMDA receptor antagonist, 2-amino-7-phosphonoheptanoic acid (AP7; 1 nmol/0.2 μL; Tocris, USA) was dissolved in sterile saline. The solutions were prepared immediately before use. The chosen doses of AEA were those that did not induce anxiolytic-like effects in a previous work from our group (Moreira et al. 2007). The doses of the antagonists used in this study were chosen based on the dose–response curves from previous studies. Since the blockade of TRPV1 and NMDA receptors in the dlPAG induces anxiolytic-like effects per se, doses that have been proven ineffective in these studies were chosen (Guimaraes et al. 1991; Molchanov and Guimaraes 2002; Terzian et al. 2009).
Apparatus
Elevated plus maze
The elevated plus maze (EPM) consisted of two opposite wooden open arms (50 × 10 cm) crossed at a right angle by two arms of the same dimensions enclosed by 40-cm-high walls (Pellow and File 1986). The maze was located 50 cm above the floor and the ANY-maze Software (version 4.5, Stoelting, USA) was employed for behavioral analysis in the apparatus. The EPM was located in a sound attenuated, temperature controlled (23 °C) room, and the environment was illuminated by a 40-W fluorescent light placed 1.3 m away from the maze. The sessions lasted for 5 min, and after each trial, the maze was cleaned with an alcohol solution (Aguiar et al. 2009).
Vogel conflict test
The Vogel conflict test (VCT) was performed in a Plexiglas box (42 × 25 × 20 cm) with a stainless grid floor. The metallic spout of a drinking bottle containing water projected into the box. The contact of the animal with the spout and the grid floor closed an electrical circuit controlled by a sensor (Anxiometer model 102, Columbus, USA), which recorded the total number of licks and shocks delivered during the test period (3 min). The animals received a 0.5-mA shock for 2 s, after 20 licks (Geller and Seifter 1960; Moreira et al. 2006).
Water consummatory evaluation
To assess whether the treatments could have affected the VCT by altering water consumption, independent groups of animals were exposed to the same experimental conditions (3 min each/section) described above except that the electric shock delivering system was rendered inoperative.
Tail flick test
To verify if the drugs interfered with nociceptive threshold, rats were submitted to the tail flick test. The apparatus consisted of an acrylic platform with a nichrome wire coil maintained at room temperature (24–26 °C). The rats were gently handled, and their tails were laid across the coil where the temperature was raised at 9 °C/s by the passage of electric current. The system had a cutoff time of 6 s to prevent tissue damage when the coil temperature approached 80 °C (Moreira et al. 2006).
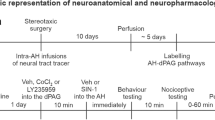
Surgery
The rats were anesthetized with tribromoethanol (250 mg/kg i.p., Sigma-Aldrich, USA) and were fixed in a stereotaxic frame. A stainless steel guide cannula (11 mm; 0.6 mm, outer diameter (OD)) was implanted unilaterally on the right side aimed at the dlPAG (coordinates: AP = 0 from lambda, L = −1.9 mm at an angle of 16°, D = −4.0 mm) according to the atlas published by Paxinos and Watson (Paxinos and Watson 1997). The cannula was attached to the bones with stainless steel screws and acrylic cement. An obturator inside the guide cannulae prevented obstruction. Immediately after the surgery, the animals received benzylpenicillin (0.27 g/kg i.m.; Pentabiotico®, Fort Dodge, Brazil) to prevent infection and a nonsteroidal anti-inflammatory drug, flunixin meglumine (0.025 g/kg s.c.; Banamine®, Schering Plough, Brazil) for postoperative analgesia.
Procedure
Intra-dlPAG injection
Seven days after surgery, the animals were randomly assigned to one of the treatment groups. Intracerebral injections were performed with a thin dental needle (0.3 mm, OD) introduced through the guide cannula until its tip was 1.0 mm below the cannula end. A volume of 0.2 μL was injected in 30 s using a microsyringe (Hamilton, USA) connected to an infusion pump (KD Scientific, USA). In order to prevent reflux, the guide cannula was left in place for 30 s after the end of each injection. A polyethylene catheter (PE 10) was interposed between the upper end of the dental needle and the syringe.
Experiment 1
The rats received intra-dlPAG injections of vehicle (saline) or AP7 (1 nmol) followed, 5 min later, by vehicle (Tocrisolve® in saline) or AEA (50 or 200 pmol). Ten minutes later, they were placed in the center of the EPM facing an enclosed arm. The percentage of entries and time spent in the open arms and the number of entries into the enclosed arms of the maze was recorded for 5 min. The experimental groups were vehicle (veh) + veh, veh + AEA 50, veh + AEA 200, AP7 + veh, AP7 + AEA 50, AP7 + AEA 200.
Experiment 2
The procedure was similar to experiment 1 except that the animals received vehicle (DMSO 100 %) or capsazepine (CPZ, 10 nmol) before vehicle (Tocrisolve® in saline) or AEA (50 or 200 pmol) injection. No additional tissue damage was observed in DMSO-treated animals compared to the other vehicles. As previously described, 10 min after the second injection the animals were submitted to the EPM test for 5 min. The experimental groups were veh + veh, veh + AEA50, veh + AEA 200, CPZ + veh, CPZ + AEA 50, CPZ + AEA 200.
Experiment 3
The animals were water-deprived for 48 h before the test. After the first 24 h of deprivation, they were allowed to drink freely for 3 min in the test cage in order to find the drinking bottle spout. Twenty-four hours later, they received intra-dlPAG injections of vehicle, capsazepine (10 nmol) or AP7 (1 nmol) followed, 5 min later, by vehicle or AEA injection (200 pmol). Ten minutes after the second injection, they were placed in the apparatus. The test period lasted for 3 min, and the animals received a 0.5-mA shock through the bottle spout every 20 licks. The number of punished licks was registered. The procedure was similar to the one already used and validated in the laboratory (Aguiar et al. 2009; Lisboa et al. 2008). The experimental groups were veh + veh, CPZ + veh, AP7 + veh, veh + AEA, CPZ + AEA, AP7 + AEA. Results from the vehicles used for CPZ and AP7 were not statistically different. Therefore, they were grouped together in the vehicle + vehicle group.
Experiment 4
The tail flick test was conducted in independent groups of animals receiving vehicle + vehicle, capsazepine + AEA, or AP7 + AEA into the dlPAG. Morphine was administered systemically (5 mg/kg i.p.) as a positive control. The animals were gently handled, and their tails were laid across the coil. The heating was applied to a portion of the ventral surface of the tail between 4 and 6 cm from its end. The time to withdraw the tail was recorded as tail flick latency. The electric current was calibrated to provoke this reflex within 2.5–3.5 s in nontreated animals. The tail flick latency was measured at 5-min intervals until a stable baseline was obtained over three consecutive trials. After drug administration, the tail withdrawal latencies were measured again at 10-min intervals for up to 30 min.
Experiment 5
The procedure was the same used in the punished licking test; however, the electric shock delivering system was render inoperative. Ten minutes after vehicle + vehicle, CPZ + AEA, or AP7 + AEA injections, rats were allowed to drink freely for 3 min. The number of licks during this period was registered.
Histology
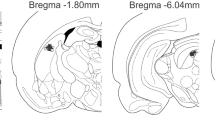
After the behavioral tests, the rats were sacrificed under deep chloral hydrate 5 % (Sigma-Aldrich, USA) anesthesia and perfused through the left ventricle of the heart with isotonic saline followed by 10 % formalin solution. After that, the brains were removed and, after a minimum period of 5 days, immersed in a 10 % formalin solution, 50 μm sections were obtained in a Cryostat (Cryocut 1800, Leica, Germany). The injection sites were identified in diagrams from the Paxinos and Watson atlas (Paxinos and Watson 2007) and are illustrated in Fig. 1. Rats receiving drug injections outside the aimed area were included in an additional (OUT) group.

a Photomicrography of a representative site injection in the dlPAG. b Histological localization of injection sites in diagrams based on the atlas of Paxinos and Watson (2007). The black and the grey circles represent the injection sites inside and outside of the dlPAG, respectively
Statistical analysis
The percentages of entries and time spent in the open arms [(open/open + enclosed) × 100] during the 5-min sessions in the EPM were calculated for each animal. Since the data from animals submitted to experiment 1 did not follow a normal distribution, the EPM data from experiment 1 and 2, percentage of entries and time spent in the open arms and number of enclosed arms, were analyzed by Kruskal–Wallis followed by Mann–Whitney test. The number of punished licks (experiment 3) was analyzed by two-way ANOVA using the first and the second injections as main factors. Data from the tail flick (experiment 4) were analyzed by repeated measures ANOVA with treatment as the independent factor and time as the repeated measure. In the case of significant interactions, a one-way ANOVA was performed followed by the Duncan test for multiple comparisons. Results from the water consumption test (experiment 5) were analyzed by one-way ANOVA followed by Duncan test. Differences were considered significant at p < 0.05 level.
Results
Experiment 1
There were significant effects of treatment on the percentage of entries (X 2 (5) = 16.96, p < 0.05) and time spent in the open arms of EPM (X 2 (5) = 12.33, p < 0.05). AP7 (1 nmol) or AEA (50 and 200 pmol) did not induce any significant effects when administered alone. However, AP7 prior to AEA significantly increased the percentage of entries (50 and 200 nmol) and time spent (only the 200 nmol dose) in the open arms of the EPM in comparison to control group (Mann–Whitney test, p < 0.05; Fig. 2), suggesting an anxiolytic-like effect. No effect was found in the number of enclosed arms entries (X 2 (5) = 8.615, p = 0.125), indicating that this effect is not affecting basal motor activity. Additionally, animals that received AP7 + AEA (200 pmol) outside the dlPAG (percentage of open arm entries = 26.2 ± 1.5, percentage of time spent in open arms = 8.8 ± 2.2, n = 6) were not different from the vehicle-treated group.

Effects of vehicle or AP7 (1 nmol) followed by vehicle or anandamide (AEA; 50 or 200 pmol) administered into the dlPAG of rats tested in the EPM. The bars represent the mean ± S.E.M. of the number of entries into the enclosed arms (a), the percentage of entries in the open arms (b), and percentage of time in the open arms (c). The data were analyzed by Kruskal–Wallis followed by Mann–Whitney test (asterisks indicate the significant difference from vehicle–vehicle group, p < 0.05, n = 5–7)
Experiment 2
There were significant effects of the percentage of time (X 2 (5) = 11.74, p = 0.038) and in the percentage of entries (X 2 (5) = 16.26, p = 0.006) in the open arms of EPM. As previously observed, AEA (50 and 200 pmol) did not induce any behavioral change (Mann–Whitney test, p > 0.05). Moreover, no effect was detected with capsazepine (10 nmol) alone (Mann–Whitney test, p > 0.05). However, capsazepine prior to AEA (50 and 200 pmol) significantly increased the percentage of entries in the open arms of EPM compared to the control group (Mann–Whitney test, p < 0.05; Fig. 3). No effect was observed in the number of enclosed arms entries (X 2 (5) = 6.09, p = 0.296). Animals that received CPZ + AEA (200 pmol) outside the dlPAG ((percentage of open arm entries = 22.3 ± 1.5, percentage of time spent in open arms = 6.3 ± 1.8, n = 6) were not different from vehicle-treated rats.

Effects of vehicle or capsazepine (CPZ, 10 nmol) followed by vehicle or AEA (50 or 200 pmol) administered into the dlPAG of rats tested in the EPM. Bars represent the mean ± S.E.M. of the number of entries into the enclosed arms (a), the percentage of entries in the open arms (b), and percentage of time in the open arms (c). The data were analyzed by Kruskal–Wallis followed by Mann–Whitney test (asterisks indicate the significant difference from vehicle–vehicle group, p < 0.05, n = 7–9)
Experiment 3
Since no difference was found between animals that received saline (n = 4) or DMSO (n = 3) followed by Tocrisolve as vehicles, they were joined together in a control group. Two-way ANOVA showed significant effects of the first (F 2,38 = 4,36; p = 0.019) and second (F 1,38 = 12.38; p = 0.001) microinjections and a significant interaction (F 2,38 = 3.28; p = 0.048). Post hoc analysis indicated that AP7 and capsazepine prior to AEA injection increased the number of punished licks in the VCT (Duncan test, p < 0.05; Fig. 4). These drugs did not induce any effect when administered alone. Moreover, animals that received AP7 + AEA or CPZ + AEA (punished licks = 92 ± 10.6, n = 8) outside the dlPAG were not different from controls.

Effects of CPZ (10 nmol) and AP7 (1 nmol) followed by AEA (200 pmol) administered into the dlPAG of rats submitted to the Vogel conflict test. Bars represent the mean ± S.E.M. total number of punished licks in the 3-min session. Asterisks indicate the significant difference from vehicle–vehicle group. The data were analyzed by two-way ANOVA followed by the Duncan test (*p < 0.05 from vehicle–vehicle group; n = 7π11)
Experiment 4
A repeated measure ANOVA showed significant effects of time (F 5,84 = 32.0, p < 0.05), treatment (F 3,84 = 74.8, p < 0.05) and interaction (F 15,84 = 15.1, p < 0.05) on the withdrawal latency in the tail flick test. Morphine, the positive control, increased the latency of tail withdrawal 10, 20, and 30 min after drug administration (Duncan test, p < 0.05; Fig. 5). Moreover, there was an increase in the latency of tail withdrawal 20 min after drug injection in animals that received CPZ or AP7 prior to AEA (Duncan test, p < 0.05; Fig. 5). This latency, however, was not different from baseline values (p > 0.05).

Time course of the effects of intra-dlPAG administration of vehicle + vehicle (Veh + Veh), capsazepine (CPZ, 10 nmol) + AEA 200 pmol, AP7 (1 nmol) + AEA (200 pmol), or systemically injected morphine (5 mg/kg, i.p.) in the tail flick test. Each point represents the mean ± S.E.M. for the latency of tail withdrawal. The data were analyzed by two-way ANOVA followed by the Duncan test (asterisks indicate the significant difference p < 0.05, from vehicle group, n = 4–5)
Experiment 5
The effects in VCT were not due an increase of water consumption since there was no difference in the number of unpunished licks among the groups (veh + veh n = 4; mean number of licks, 964.3 ± 53.1; CPZ + AEA n = 5; mean number of licks, 991.6 ± 52.4; AP7 + AEA n = 5; mean number of licks, 876 ± 29.1; F 2,11 = 1.9, Duncan test, p = 0.19).
Discussion
The present results showed that high, ineffective doses of AEA in the dlPAG can become anxiolytic when animals were pretreated with TRPV1 or NMDA antagonists. This anxiolytic-like effect was observed in two distinct animal models of anxiety, the EPM and VCT, and depended on drug effects in the dlPAG, since no difference was observed when the injections were performed near, but not into, this region. The drug-induced decrease in anxiety was not accompanied by changes in the number of enclosed arms entries in the EPM, discarding motor effect as a possible confounding factor (File 1992). Moreover, control experiments for the VCT showed that neither AEA plus AP7 nor AEA plus CPZ altered water consumption. Both treatments, however, did increase the latency of tail withdrawal 20 min after injection, suggesting an antinociceptive effect at this time point. Nevertheless, in the present study the VCT was performed 10 min after injections, a time point where no differences were found in the tail flick test. In addition, the antinociceptive effect was small and did not differ from the baseline measures. Therefore, it seems unlikely that the anticonflict results obtained in the VCT were due to antinociceptive effects. In previous studies of our group, similar results (anxiolytic with a small antinociceptive effect) were obtained after intra-dlPAG injection of the inhibitor of anandamide uptake AM404 or the phytocannabinoid cannabidiol (Campos and Guimaraes 2008; Lisboa et al. 2008). This is hardly surprising, since cannabinoid-, vanilloid-, and glutamatergic-mediated neurotransmissions are involved in nociceptive control in this brain area (Finn et al. 2003; Maione et al. 2006, 2009; Palazzo et al. 2001, 2002; Walker et al. 1999).
Together, the present results indicate that at high doses, such as those used in the present study (Moreira et al. 2007), the anxiolytic-like effects of AEA in the dlPAG disappear due to its ability to activate TRPV1 receptors. In agreement with this possibility, the affinity of AEA at TRPV1 is relatively low compared to CB1 receptors (Ross 2003).
Different from CB1 receptors, TRPV1 seems to mediate pro-aversive effects. TRPV1 knockout mice showed reduced anxiety-like behavior in the EPM and light–dark box, as well as reduced fear conditioning (Marsch et al. 2007). Moreover, either systemic or local injection of TRPV1 antagonists into specific brain structures, such as the dlPAG (Terzian et al. 2009), prefrontal cortex (Aguiar et al. 2009; Fogaca et al. 2012; Rubino et al. 2008), and ventral hippocampus (Santos et al. 2008), induces anxiolytic-like effects in rats.
In the CNS, the main effect resulting from activation of TRPV1 receptors is the facilitation of glutamatergic neurotransmission (Starowicz et al. 2007; Urban and Dray 1992; Urban et al. 1985; Xing and Li 2007). Neurochemical and electrophysiological data showed that activation of TRPV1 induced glutamate release through a mechanism dependent on Ca+2, which was inhibited by TRPV1 antagonism (Palazzo et al. 2002).
Increased glutamate release in the dlPAG induced by activation of TRPV1 receptors after high doses of some cannabinoids such as AEA (Palazzos et al. 2006) could facilitate defensive responses (Guimaraes et al. 2005). Accordingly, in the presence of an NMDA antagonist, ineffective high doses of AEA became anxiolytic in animals exposed to both EPM and VCT. Even if we have not measured local glutamate release, several studies corroborate this possibility. For example, injection of capsaicin into the dlPAG induced nociception and local release of glutamate, an effect reversed by pretreatment with the TRPV1 antagonist capsazepine (Palazzo et al. 2002). Similar effects were described in vitro for other regions such as the hypothalamus, substantia nigra, and locus coeruleus (Marinelli et al. 2002, 2003; Palazzo et al. 2002). Furthermore, supporting the hypothesis of interaction between glutamatergic and vanilloid systems, Starowicz et al. (2007) showed that neurons in the ventrolateral portion of the PAG, which also express TRPV1 receptors, are surrounded by glutamatergic neurons. Additionally, these glutamatergic neurons respond to TRPV1 stimulation by releasing glutamate, an effect blocked by pretreatment with a vanilloid antagonist (Starowicz et al. 2007). The expression of TRPV1 in glutamatergic neurons was also detected in the dorsal horn of the spinal cord (Zhou et al. 2009).
In order to verify which glutamate receptors are related to the nociceptive responses induced by capsaicin, Jin et al. (2012) observed that capsaicin-evoked c-Fos expression in the spinal cord dorsal horn was prevented by NMDA receptor antagonists, suggesting that activation of ionotropic glutamate receptors are involved in nociceptive responses induced by capsaicin (Jin et al. 2012). Moreover, the antidepressant and anxiolytic-like behavioral profiles observed in TRPV1 knockout mice were associated with changes in the expression of NMDA receptors in the striatum and CA1 region of the hippocampus (You et al. 2012). Altogether, these results suggest that NMDA receptors are involved in the effects mediated by TRPV1 activation.
The atypical neurotransmitters nitric oxide has also been suggested to facilitate defensive responses in the dlPAG by facilitating glutamate release (Guimaraes et al. 2005). Further supporting an interaction between NMDA and TRPV1 receptors, AEA increased the pro-aversive effects of nitric oxide in the dlPAG through activation of latter receptors (Lisboa and Guimaraes 2012).
In conclusion, the present study showed that local dlPAG pretreatment with TRPV1 or NMDA receptor antagonists unveiled the anxiolytic-like effect of high, ineffective doses of AEA, suggesting that at higher doses, facilitation of glutamate release in this region by TRPV1 activation prevents the anxiolytic-like effects of this compound.
References
Aguiar DC, Guimaraes FS (2009) Blockade of NMDA receptors and nitric oxide synthesis in the dorsolateral periaqueductal gray attenuates behavioral and cellular responses of rats exposed to a live predator. J Neurosci Res 87:2418–2429
Aguiar DC, Moreira FA, Guimaraes FS (2006) Flight reactions induced by injection of glutamate N-methyl-d-aspartate receptor agonist into the rat dorsolateral periaqueductal gray are not dependent on endogenous nitric oxide. Pharmacol Biochem Behav 83:296–301
Aguiar DC, Terzian AL, Guimaraes FS, Moreira FA (2009) Anxiolytic-like effects induced by blockade of transient receptor potential vanilloid type 1 (TRPV1) channels in the medial prefrontal cortex of rats. Psychopharmacol (Berl) 205:217–225
Bandler R, Keay KA, Floyd N, Price J (2000) Central circuits mediating patterned autonomic activity during active vs. passive emotional coping. Brain Res Bull 53:95–104
Bittencourt AS, Carobrez AP, Zamprogno LP, Tufik S, Schenberg LC (2004) Organization of single components of defensive behaviors within distinct columns of periaqueductal gray matter of the rat: role of N-methyl-d-aspartic acid glutamate receptors. Neuroscience 125:71–89
Campos AC, Guimaraes FS (2008) Involvement of 5HT1A receptors in the anxiolytic-like effects of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacol (Berl) 199:223–230
Carrive P (1993) The periaqueductal gray and defensive behavior: functional representation and neuronal organization. Behav Brain Res 58:27–47
Casarotto PC, Terzian AL, Aguiar DC, Zangrossi H, Guimaraes FS, Wotjak CT, Moreira FA (2012) Opposing roles for cannabinoid receptor type-1 (CB(1)) and transient receptor potential vanilloid type-1 channel (TRPV1) on the modulation of panic-like responses in rats. Neuropsychopharmacology 37:478–486
Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D (1997) The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389:816–824
Cavanaugh DJ, Chesler AT, Jackson AC, Sigal YM, Yamanaka H, Grant R, O'Donnell D, Nicoll RA, Shah NM, Julius D, Basbaum AI (2011) Trpv1 reporter mice reveal highly restricted brain distribution and functional expression in arteriolar smooth muscle cells. J Neurosci 31:5067–5077
Clapham DE, Julius D, Montell C, Schultz G (2005) International Union of Pharmacology. XLIX. Nomenclature and structure-function relationships of transient receptor potential channels. Pharmacol Rev 57:427–450
Cristino L, de Petrocellis L, Pryce G, Baker D, Guglielmotti V, Di Marzo V (2006) Immunohistochemical localization of cannabinoid type 1 and vanilloid transient receptor potential vanilloid type 1 receptors in the mouse brain. Neuroscience 139:1405–1415
Di Marzo V, Bisogno T, De Petrocellis L (2001) Anandamide: some like it hot. Trends Pharmacol Sci 22:346–349
File SE (1992) Behavioural detection of anxiolytic action. In: Elliot JM, Heal DJ, Marsden CA (eds) Experimental approaches to anxiety and depression. Wiley, New York, pp 25–44
Finn DP, Jhaveri MD, Beckett SR, Roe CH, Kendall DA, Marsden CA, Chapman V (2003) Effects of direct periaqueductal grey administration of a cannabinoid receptor agonist on nociceptive and aversive responses in rats. Neuropharmacology 45:594–604
Fogaca MV, Aguiar DC, Moreira FA, Guimaraes FS (2012) The endocannabinoid and endovanilloid systems interact in the rat prelimbic medial prefrontal cortex to control anxiety-like behavior. Neuropharmacology 63:202–210
Geller I, Seifter J (1960) The effects of meprobamate, barbiturate, damphetamine and promazine on experimentally induced conflict in the rat. Psychopharmacologia 1:482–492
Guimaraes FS, Carobrez AP, De Aguiar JC, Graeff FG (1991) Anxiolytic effect in the elevated plus-maze of the NMDA receptor antagonist AP7 microinjected into the dorsal periaqueductal grey. Psychopharmacol (Berl) 103:91–94
Guimaraes FS, Beijamini V, Moreira FA, Aguiar DC, de Lucca AC (2005) Role of nitric oxide in brain regions related to defensive reactions. Neurosci Biobehav Rev 29:1313–1322
Jin YH, Takemura M, Furuyama A, Yonehara N (2012) Peripheral glutamate receptors are required for hyperalgesia induced by capsaicin. Pain Res Treat 2012:915706
Kawahara H, Drew G, Christie M, Vaughan C (2011) Inhibition of fatty acid amide hydrolase unmasks CB(1) receptor and TRPV1 channel-mediated modulation of glutamatergic synaptic transmission in midbrain periaqueductal grey. Br J Pharmacol 163:1214–1222
Lisboa SF, Guimaraes FS (2012) Differential role of CB1 and TRPV1 receptors on anandamide modulation of defensive responses induced by nitric oxide in the dorsolateral periaqueductal gray. Neuropharmacology 62:2455–2462
Lisboa SF, Resstel LB, Aguiar DC, Guimaraes FS (2008) Activation of cannabinoid CB1 receptors in the dorsolateral periaqueductal gray induces anxiolytic effects in rats submitted to the Vogel conflict test. Eur J Pharmacol 593:73–78
Maione S, Bisogno T, de Novellis V, Palazzo E, Cristino L, Valenti M, Petrosino S, Guglielmotti V, Rossi F, Di Marzo V (2006) Elevation of endocannabinoid levels in the ventrolateral periaqueductal grey through inhibition of fatty acid amide hydrolase affects descending nociceptive pathways via both cannabinoid receptor type 1 and transient receptor potential vanilloid type-1 receptors. J Pharmacol Exp Ther 316:969–982
Maione S, Cristino L, Migliozzi AL, Georgiou AL, Starowicz K, Salt TE, Di Marzo V (2009) TRPV1 channels control synaptic plasticity in the developing superior colliculus. J Physiol 587:2521–2535
Marinelli S, Vaughan CW, Christie MJ, Connor M (2002) Capsaicin activation of glutamatergic synaptic transmission in the rat locus coeruleus in vitro. J Physiol 543:531–540
Marinelli S, Di Marzo V, Berretta N, Matias I, Maccarrone M, Bernardi G, Mercuri NB (2003) Presynaptic facilitation of glutamatergic synapses to dopaminergic neurons of the rat substantia nigra by endogenous stimulation of vanilloid receptors. J Neurosci 23:3136–3144
Marsch R, Foeller E, Rammes G, Bunck M, Kossl M, Holsboer F, Zieglgansberger W, Landgraf R, Lutz B, Wotjak CT (2007) Reduced anxiety, conditioned fear, and hippocampal long-term potentiation in transient receptor potential vanilloid type 1 receptor-deficient mice. J Neurosci 27:832–839
Matheus MG, Guimaraes FS (1997) Antagonism of non-NMDA receptors in the dorsal periaqueductal grey induces anxiolytic effect in the elevated plus maze. Psychopharmacol (Berl) 132:14–18
McGaraughty S, Chu KL, Bitner RS, Martino B, El Kouhen R, Han P, Nikkel AL, Burgard EC, Faltynek CR, Jarvis MF (2003) Capsaicin infused into the PAG affects rat tail flick responses to noxious heat and alters neuronal firing in the RVM. J Neurophysiol 90:2702–2710
McGregor IS, Hargreaves GA, Apfelbach R, Hunt GE (2004) Neural correlates of cat odor-induced anxiety in rats: region-specific effects of the benzodiazepine midazolam. J Neurosci 24:4134–4144
Molchanov ML, Guimaraes FS (2002) Anxiolytic-like effects of AP7 injected into the dorsolateral or ventrolateral columns of the periaqueductal gray of rats. Psychopharmacol (Berl) 160:30–38
Moreira FA, Aguiar DC, Guimaraes FS (2006) Anxiolytic-like effect of cannabidiol in the rat Vogel conflict test. Prog Neuropsychopharmacol Biol Psychiatry 30:1466–1471
Moreira FA, Aguiar DC, Guimaraes FS (2007) Anxiolytic-like effect of cannabinoids injected into the rat dorsolateral periaqueductal gray. Neuropharmacology 52:958–965
Moreira FA, Aguiar DC, Terzian AL, Guimaraes FS, Wotjak CT (2012) Cannabinoid type 1 receptors and transient receptor potential vanilloid type 1 channels in fear and anxiety-two sides of one coin? Neuroscience 204:186–192
Palazzo E, Marabese I, de Novellis V, Oliva P, Rossi F, Berrino L, Maione S (2001) Metabotropic and NMDA glutamate receptors participate in the cannabinoid-induced antinociception. Neuropharmacology 40:319–326
Palazzo E, de Novellis V, Marabese I, Cuomo D, Rossi F, Berrino L, Rossi F, Maione S (2002) Interaction between vanilloid and glutamate receptors in the central modulation of nociception. Eur J Pharmacol 439:69–75
Palazzos E, de Novellis V, Marabese I, Rossi F, Maione S (2006) Metabotropic glutamate and cannabinoid receptor crosstalk in periaqueductal grey pain processing. Curr Neuropharmacol 4:225–231
Paxinos G, Watson C (1997) The rat brain in stereotaxic coordinates, 3rd edn. Academic Press, New York
Paxinos G, Watson C (2007) The rat brain in stereotaxic coordinates, 6th edn. Academic Press/Elsevier, London
Pellow S, File SE (1986) Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: a novel test of anxiety in the rat. Pharmacol Biochem Behav 24:525–529
Pertwee RG (2006) The pharmacology of cannabinoid receptors and their ligands: an overview. Int J Obes (Lond) 30(Suppl 1):S13–S18
Ross RA (2003) Anandamide and vanilloid TRPV1 receptors. Br J Pharmacol 140:790–801
Rubino T, Realini N, Castiglioni C, Guidali C, Vigano D, Marras E, Petrosino S, Perletti G, Maccarrone M, Di Marzo V, Parolaro D (2008) Role in anxiety behavior of the endocannabinoid system in the prefrontal cortex. Cereb Cortex 18:1292–1301
Santos CJ, Stern CA, Bertoglio LJ (2008) Attenuation of anxiety-related behaviour after the antagonism of transient receptor potential vanilloid type 1 channels in the rat ventral hippocampus. Behav Pharmacol 19:357–360
Smart D, Jerman JC (2000) Anandamide: an endogenous activator of the vanilloid receptor. Trends Pharmacol Sci 21:134
Starowicz K, Maione S, Cristino L, Palazzo E, Marabese I, Rossi F, de Novellis V, Di Marzo V (2007) Tonic endovanilloid facilitation of glutamate release in brainstem descending antinociceptive pathways. J Neurosci 27:13739–13749
Terzian AL, Aguiar DC, Guimaraes FS, Moreira FA (2009) Modulation of anxiety-like behaviour by Transient Receptor Potential Vanilloid Type 1 (TRPV1) channels located in the dorsolateral periaqueductal gray. Eur Neuropsychopharmacol 19:188–195
Toth A, Boczan J, Kedei N, Lizanecz E, Bagi Z, Papp Z, Edes I, Csiba L, Blumberg PM (2005) Expression and distribution of vanilloid receptor 1 (TRPV1) in the adult rat brain. Brain Res Mol Brain Res 135:162–168
Urban L, Dray A (1992) Synaptic activation of dorsal horn neurons by selective C-fibre excitation with capsaicin in the mouse spinal cord in vitro. Neuroscience 47:693–702
Urban L, Willetts J, Randic M, Papka RE (1985) The acute and chronic effects of capsaicin on slow excitatory transmission in rat dorsal horn. Brain Res 330:390–396
Vaughan CW, Connor M, Bagley EE, Christie MJ (2000) Actions of cannabinoids on membrane properties and synaptic transmission in rat periaqueductal gray neurons in vitro. Mol Pharmacol 57:288–295
Viveros MP, Marco EM, Llorente R, Lopez-Gallardo M (2007) Endocannabinoid system and synaptic plasticity: implications for emotional responses. Neural Plast 2007:52908
Walker JM, Huang SM, Strangman NM, Tsou K, Sanudo-Pena MC (1999) Pain modulation by release of the endogenous cannabinoid anandamide. Proc Natl Acad Sci USA 96:12198–12203
Xing J, Li J (2007) TRPV1 receptor mediates glutamatergic synaptic input to dorsolateral periaqueductal gray (dl-PAG) neurons. J Neurophysiol 97:503–511
You IJ, Jung YH, Kim MJ, Kwon SH, Hong SI, Lee SY, Jang CG (2012) Alterations in the emotional and memory behavioral phenotypes of transient receptor potential vanilloid type 1-deficient mice are mediated by changes in expression of 5-HT(1A), GABA(A), and NMDA receptors. Neuropharmacology 62:1034–1043
Zhou HY, Chen SR, Chen H, Pan HL (2009) The glutamatergic nature of TRPV1-expressing neurons in the spinal dorsal horn. J Neurochem 108:305–318
Zygmunt PM, Julius I, Di Marzo I, Hogestatt ED (2000) Anandamide—the other side of the coin. Trends Pharmacol Sci 21:43–44
Acknowledgments
This research was supported by grants from CAPES, CNPq, FAPESP, and FAPEMIG (APQ-01883-10). We thank J.C. de Aguiar and E.T. Gomes for the excellent technical support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Fogaça M.V. and Gomes F.V. contributed equally to this work.
Rights and permissions
About this article
Cite this article
Fogaça, M.V., Gomes, F.V., Moreira, F.A. et al. Effects of glutamate NMDA and TRPV1 receptor antagonists on the biphasic responses to anandamide injected into the dorsolateral periaqueductal grey of Wistar rats. Psychopharmacology 226, 579–587 (2013). https://doi.org/10.1007/s00213-012-2927-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-012-2927-2