
Abstract
For the first time in the Western Alps, radiolarite pebbles collected from Tertiary foreland molasse conglomerates are treated for microfossil extraction and dated. Among forty pebbles collected in the field, seven of them released diagnostic radiolarian assemblages ranging in age from Late Bajocian–Early Callovian to Middle Oxfordian–Early Tithonian. These ages overlap previous biochronological data obtained from in situ localities of the Schistes Lustrés Piemont zone of the French-Italian Alps and triple the number of diagnostic radiolarite samples known so far in this segment of the chain. The diagnostic pebbles are characterized by low grade metamorphism, showing that some eroded thrust-sheets from the oldest parts of the ocean escaped any tectonic burial during the Alpine convergence. Mixing of low and high-grade radiolarites, mafics and ultramafics pebbles implies that various ocean-derived units were exposed. This tectonic scenario involves tight refolding and severe uplift of the Eocene subduction wedge in the Early Oligocene.
Similar content being viewed by others

1 Introduction
The European Alps correspond to the plate boundary between Europe and Africa where divergence dominated during most of Jurassic–Cretaceous times followed by convergence from the Late Cretaceous onwards (e.g. Tricart 1984). As a major witness to plate divergence, the Alpine ophiolites originated from a small Tethyan Ocean basin, the Alpine Tethys also named Liguria, Liguria–Piemont or Piemont–Liguria (review in Bernoulli and Jenkyns 2009; Lagabrielle 2009; Manatschal and Müntener 2009; de Graciansky et al. 2010). In the Western Alps, dismembered ophiolites and associated deep-sea sediments compose most of the Piemont zone (review in Schmid et al. 2004). They display a subduction-related HP-LT metamorphic imprint evolving from blueschist facies in the outer Piemont units to eclogite facies in the inner ones (review in Bousquet et al. 2008). As an exception, the Chenaillet ophiolite (Fig. 1) escaped this metamorphism, suggesting that it derived from an obducted slab of oceanic lithosphere (Mevel et al. 1978; Barféty et al. 1995 with references therein).

Location map of southern French-Italian Alps (modified from Schmid et al. 2004). Location of sampled molasse conglomerates: M Montmaur, B Barrême. Known sites with in situ bedrock fossiliferous radiolarites: P Rocher de la Perdrix (Chenaillet Massif), C Cascavelier Peak (southern Queyras), T Traversiera Massif. Red Major fault zones
Reconstructing the paleogeography of this vanished ocean at successive steps from opening to closure remains a major challenge among Alpine geologists. Two difficulties remained for a long time: the rarity of radiometric ages from ophiolites and the missing fossil record due to metamorphism and/or erosion of sedimentary covers. Fortunately, some of the Western Alps units closely resemble the unmetamorphic ophiolite thrust sheets of the Northern Apennines, which suggests that Tethyan Ocean spreading occurred during the Middle–Late Jurassic and possibly the Early Cretaceous (e.g. Lemoine et al. 1970, de Graciansky et al. 2010). Most modern reconstructions for the Western Alpine Ocean derive from Marcel Lemoine’s sketches of the early 1980s. They show the Alpine Tethys and the Central Atlantic as twin oceans for their initial opening and spreading (e.g. Lemoine 1983). On the Tethyan European paleomargin, the transgressive units and their fossil record exposed in the Briançonnais domain document the breakup unconformity (Lemoine et al. 1986) and provide a Middle Jurassic age (Late Bathonian) for the timing of initial opening of the nearby ocean (Lemoine et al. 2000; Bill et al. 2001).
A significant improvement came with the successive dating of three radiolarite supraophiolitic localities: (1) the Cascavelier Peak near Saint-Véran in Southern Queyras (De Wever and Caby 1981; De Wever and Baumgartner 1995), (2) the Traversiera massif near the French–Italian border in the upper Ubaye valley (Schaaf et al. 1985; De Wever et al. 1987; De Wever and Baumgartner 1995), and (3) more recently the Rocher de la Perdrix in the Montgenèvre massif, located structurally below the Chenaillet ophiolite s.s. (Cordey and Bailly 2007). In these three sites (Fig. 1), radiolarian faunas are preserved in low-temperature blueschist-bearing metacherts at the front of the Piemont “Schistes lustrés” complex (Gidon et al. 1994; Tricart et al. 2003). These three fossil localities directly argue for an ocean opening during Middle and Late Jurassic times (Cordey and Bailly 2007).
In order to improve our knowledge of the Alpine ocean history, we explored the potential of synorogenic conglomerates deriving from the erosion of oceanic thrust sheets building the Alpine wedge. Rüst (1885) was the first to describe Jurassic radiolarians from chert pebbles of western Switzerland molasse, but microfossil chemical extractions were not yet applied at the time. A more recent interest for syn- or postorogenic clastics was developed in the North American Cordillera on the well-known “chert pebble conglomerates” associated with Mesozoic ocean sutures and accreted terranes (Seiders and Blome 1984; Cordey 1992a, b, 1998). In this Alpine study, our objectives were not only to find new dates related to the Jurassic opening of Tethys but also to document ocean closure and subsequent mountain building during Early Oligocene times.
2 The Oligocene polygenic conglomerates in the foreland of the southern Western Alps
Within the Paleogene molasse of the Western Alps external zone, our investigation focused on the French Subalpine chains located to the south of the Pelvoux massif (Ford et al. 1999; Ford and Lickorish 2004). In this region, Early Oligocene intra-orogenic basins (Fig. 2) contain conglomerates known for their ocean-derived pebbles (Termier 1895 in Gubler 1958; Boussac 1912) such as fragments of mafics and ultramafics rocks as well as radiolarian chert (Ferry et al. 2005). On the outcrop, these radiolarite pebbles are easily recognizable by their dark red colour (Fig. 3). They represent the “exotic” pebbles par excellence, occupying a privileged position in the Alpine literature as symbols of a “Penninic” signature, the Swiss term to describe the internal Alpine zones. The Early Oligocene molasse units are preserved within small basins in the foreland of the Paleogene Alpine accretionary wedge (e.g. Sissingh 2001) and were fed by clastic sediments derived from the erosion of uplifting Briançonnais and Piemont zones (Morag et al. 2008; Bernet and Tricart 2011). Initially, our main target was the Barrême basin, a small and well-studied valley located to the east of Digne thrust (de Graciansky 1972; Artoni and Meckel 1998; Evans and Elliott 1999). A second site was found farther north in the “Petit Buëch” valley near Montmaur (Fig. 1) where tectonically pinched molasses named “Nagelfluh” correspond to the southward extension of the Devoluy basin (Gidon 1971; Meckel et al. 1996).

Schematic stratigraphy of the Tertiary foreland series exposed in the French Subalpine zones, more specifically the Barrême basin. Barton. Bartonian, Priabon. Priabonian, Aquitan. Aquitanian, conglo. conglomerate, biostr. biostrome. The origin of Barrême basin pebbles are the Clumanc conglomerates

Outcrop of “Nagelfluh” molasse conglomerate near the village of Montmaur. Circles Pebbles of red radiolarites
3 Biostratigraphic results: faunal assemblages and ages
Seven diagnostic chert pebbles were found out of 40 pebbles collected in the field. Each pebble was processed individually using standard chert-etching techniques with hydrofluoric acid. Pebble 7064-E1 was collected at “La Poste de Clumanc” conglomerates (Fig. 2) near the northern end of the Barrême basin (e.g. Chauveau and Lemoine 1961). Six pebbles numbered 7057-3 to 7057-22 were sampled in the “Petit Buëch” valley (Gidon 1971).
The faunal contents and ages of the studied pebbles are presented in Table 1. Key radiolarian taxa are illustrated in Fig. 4. We use the standard Tethyan radiolarian zonation of Baumgartner et al. (1995) based on Unitary Associations (UAZ) as well as complementary data from O’Dogherty et al. (2006). The “age of a pebble” refers here to the age of the source radiolarite bed, not the age of formation or deposition of the pebble.

Middle and Late Jurassic radiolarians from Oligocene molasse chert pebbles (scanning electron microscope). For each microfossil: taxon, sample number, FC database picture number, length of scale bar. 1 Pseudoeucyrtis sp., 7057-14, no. 10, 260 μm; 2 Archeospongoprunum elegans Wu, 7057-22, no. 13, 150 μm; 3 Archeospongoprunum sp., 7057-E1, no. 2, 250 μm; 4 Cinguloturris carpatica Dumitrica, 7057-14, no. 11, 200 μm; 5 Cinguloturris sp., 7057-22, no. 6, 130 μm; 6 Archeodictyomitra apiarum (Rüst), 7057-14, no. 26, 200 μm; 7 Archeodictyomitra sp., 7057-14, no. 28, 150 μm; 8 Archeodictyomitra sp., 7057-22, no. 20, 125 μm; 9 Archeodictyomitra sp., 7057-14, no. 14, 150 μm; 10 ?Parvicingula mashitaensis Mizutani, 7057-14, no. 4, 230 μm; 11 Parahsuum carpathicum Widz and De Wever, 7057-14, no. 15, 150 μm; 12 Yaocapsa sp., 7057-15, no. 4, 160 μm; 13 Protunuma sp., 7057-14, no. 22, 140 μm; 14 ?Stichomitra sp., 7057-08, no. 14, 150 μm; 15 Pseudodictyomitrella cf. tuscanica Chiari, Cortese and Marcucci, 7057-22, no. 12; 100 μm; 16 Zhamoidellum cf. argandi O’Dogherty, Gorican and Dumitrica, 7057-15, no. 16, 130 μm; 17 Zhamoidellum ovum Dumitrica, 7057-14, no. 19, 160 μm; 18 Stichocapsa aff. robusta, 7057-08, no. 6, 150 μm; 19 Zhamoidellum sp., 7057-14, no. 27, 185 μm; 20 Pseudoristola tsunoensis (Aita), 7057-22, no. 22, 110 μm; 21 Arcanicapsa cf. leiostraca (Foreman), 7057-22, no. 3, 140 μm; 22 Zhamoidellum cf. calamin O’Dogherty, Gorican and Dumitrica, 7057-15, no. 7, 120 μm; 23 Williriedellum sp. 1 (O’Dogherty et al. 2006), 7057-22, no. 9, 150 μm; 24 Podobursa cf. spinosa Ozvoldova, 7057-14, no. 5, 200 μm; 25 ?Fultocapsa sp., 7057-22, no. 23, 130 μm; 26 Acaeniotyle diaphorogona Foreman, 7057-14, no. 1, 200 μm; 27 Emiluvia cf. chica Foreman, 7057-22, no. 15, 200 μm; 28 Homoeoparonaella argolidensis Baumgartner, 7057-22, no. 1, 200 μm; 29 Bernoullius cf. dicera (Baumgartner), 7057-20, no. 6, 150 μm; 30 Tritrabs rhododactylus Baumgartner, 7057-22, no. 18, 200 μm; 31 Mirifusus sp., 7057-14, no. 3, 250 μm
Biochronology and ages are based on the following radiolarian occurrences:
-
7057-3 contains Archeospongoprunum cf. imlayi Pessagno, Orbiculiformella sp., Stichocapsa robusta Matsuoka, and Stichocapsa sp. The age is provided by the occurrence of S. robusta Matsuoka known from UAZ 5 to UAZ 7 corresponding to the Late Bajocian–Early Callovian interval.
-
7057-8 comprises Cinguloturris carpatica Dumitrica, Praeconocaryomma sp., S. aff. robusta Matsuoka (Fig. 4, no. 18) and ?Stichomitra sp. (Fig. 4, no. 14). This assemblage is assigned to biozones UAZ 7-11 of Late Bathonian–Early Callovian to Late Kimmeridgian–Early Tithonian age.
-
7057-14 contains Acaeniotyle diaphorogona Foreman (Fig. 4, no. 26), Archeodictyomitra apiarum (Rüst), Archeodictyomitra sp. (Fig. 4, no. 7), C. carpatica Dumitrica (Fig. 4, no. 4), Emiluvia ordinaria Ozvoldova, Mirifusus sp. (Fig. 4, no. 31), Parahsuum carpathicum Widz & De Wever (Fig. 4, no. 11), ?Parvicingula mashitaensis Mizutani (Fig. 4, no. 10), Podobursa cf. spinosa Ozvoldova (Fig. 4, no. 24), Protunuma sp. (Fig. 4, no. 13), Pseudoeucyrtis sp. (Fig. 4, no. 1), Zhamoidellum ovum Dumitrica (Fig. 4, no. 17) and Zhamoidellum sp. (Fig. 4, no. 19). This assemblage is attributed to biozones UAZ 9-11 of Middle Oxfordian to Early Tithonian age.
-
7057-15 contains a peculiar association which is not reported in Baumgartner et al. (1995) biozonation but is documented by O’Dogherty et al. (2006): Yaocapsa sp. (Fig. 4, no. 12), C. carpatica Dumitrica, Zhamoidellum cf. argandi O’Dogherty, Gorican & Dumitrica (Fig. 4, no. 16) and Zhamoidellum cf. calamin O’Dogherty, Gorican & Dumitrica (Fig. 4, no. 22). This assemblage is tentatively assigned to UAZ 6–7 (O’Dogherty et al. 2006).
-
7057-20 contains A. apiarum (Rüst), Bernoullius cf. dicera Baumgartner (Fig. 4, no. 29), Praeconocaryomma sp. and Stichocapsa sp. It provides a Middle Callovian–Early Kimmeridgian age (UAZ 8–10). As the occurrence of species B. dicera Baumgartner is uncertain, the upper age limit of this association is tentative.
-
7057-22 correlation to biozone UAZ 7 of Late Bathonian–Early Callovian age is provided by co-occurrence of C. carpatica Dumitrica (Fig. 4, no. 4) and Pseudoristola tsunoensis (Aita) (Fig. 4, no. 20). Other radiolarians compatible with this age determination are Williriedellum sp. 1 (Fig. 4, no. 23) only documented in Les Gets nappe along with morphotypes from UAZ 6 and 7 (O’Dogherty et al. 2006) and Archeospongoprunum elegans Wu (Fig. 4, no. 2) also known in this time interval. Other morphotypes present in 7057-22 are Arcanicapsa cf. leiostraca (Foreman) (Fig. 4, no. 21), Archeodictyomitra sp. (Fig. 4, no. 8), Cinguloturris sp. (Fig. 4, no. 5), Emiluvia cf. chica Foreman (Fig. 4, no. 27), ?Fultocapsa sp. (Fig. 4, no. 25), Homoeoparonaella argolidensis Baumgartner (Fig. 4, no. 28), Mirifusus sp., Pseudodictyomitrella cf. tuscanica Chiari, Cortese & Marcucci (Fig. 4, no. 15), and Tritrabs rhododactylus Baumgartner (Fig. 4, no. 30).
-
7064-E1 contains A. apiarum (Rüst) (Fig. 4, no. 6), A. cf. imlayi Pessagno and C. carpatica Dumitrica (Fig. 4, no. 4). This assemblage is assigned to UAZ 8–11 of Middle Callovian to Early Tithonian age.
In summary:
-
1.
Our chert pebbles range in age from Late Bajocian–Early Callovian (UAZ 5–7) to Middle Oxfordian–Early Tithonian (UAZ 8-11).
-
2.
The oldest pebble could be as old as Late Bajocian (base of UAZ 5) but also as young as Early Callovian (top of UAZ 7).
-
3.
The youngest pebble could be as old as Middle Oxfordian (7057-14) and as young as Early Tithonian (7057-14 or 7064-E1).
-
4.
Three pebbles are Middle Jurassic in age. Among them, 7057-22 is restricted to UAZ 7 of Late Bathonian–Early Callovian age. The two others 7057-3 and 7057-15 are possibly of the same age but could also be younger (UAZ 5–7 and UAZ 6–7, respectively).
-
5.
Only one pebble (7057-14) is with certainty Late Jurassic in age. The three remaining pebbles are either Middle or Late Jurassic (7057-8, 7057-20, and 7064-E1).
4 Comparisons with previous radiolarian localities of the Piemont Schistes Lustrés zone
4.1 Biochronology
As mentioned previously, only three localities of supraophiolitic radiolarites are dated so far in the Schistes Lustrés complex of the French-Italian Alps. They range in age from Middle Bathonian to Middle–Late Oxfordian:
-
1.
The Cascavelier Peak radiolarite near Saint-Véran was originally dated as Late Oxfordian–Middle Kimmeridgian (De Wever and Caby 1981). This age was later revised to the Middle or Late Oxfordian (UAZ 9) (De Wever and Baumgartner 1995).
-
2.
The Traversiera massif locality was originally found by R. Polino (Torino), with radiolarians occurring in phosphate nodules at the base of the sedimentary cover of the ophiolite (Y. Lagabrielle, personal communication, 2006). It was dated as Late Oxfordian–Early Kimmeridgian (Schaaf et al. 1985), an age later revised as Late Bathonian–Early Callovian (UAZ 7) (De Wever et al. 1987; De Wever and Baumgartner 1995).
-
3.
The “Rocher de la Perdrix” radiolarites from the Chenaillet massif near Montgenèvre are exposed in the Lago Nero-Replatte thrust sheet located structurally beneath the Chenaillet ophiolite thrust sheet s.s. It was tentatively assigned to UAZ 6 of Middle Bathonian age. It could be the oldest radiolarite exposure of the Piemont zone (Cordey and Bailly 2007).
In comparing our results with these biochronological data, we find that the ages of radiolarite pebbles overlap almost entirely the age range known from in situ Piemont Schistes Lustrés radiolarite localities. Our results also lead to the following observations:
-
1.
In our sampling, biozone UAZ 7 seems well represented in 2 pebbles, and possibly even in 4. It matches the age established in the Traversiera massif (Late Bathonian–Early Callovian).
-
2.
The occurrence of biozone UAZ 6 known at the Chenaillet is not confirmed but could be present in 2 of the 7 pebbles.
-
3.
Our youngest pebble could be coeval to the Pic Cascavelier locality of Middle–Late Oxfordian age (UAZ 9) but could also be younger (Kimmeridgian or Early Tithonian).
4.2 Faunal content
Some radiolarian morphotypes present in our pebbles were not reported before in the Piemont Schistes Lustrés in situ radiolarites. The Pic Cascavelier locality (UAZ 7) has an assemblage of 16 species in which only H. argolidensis Baumgartner occurs in one pebble (7057-22). The St-Véran section (UAZ 9) comprises 19 species in which only A. diaphorogona Foreman and potentially B. dicera (Baumgartner) are present in our assemblages. Finally, the Rocher de la Perdrix locality has only five morphotypes with two species of the genus Kilinora, which does not occur elsewhere. Overall, our pebbles seem richer in nassellarians, including morphotypes of Cinguloturris, Fultocapsa, Protunuma, Pseudodictyomitrella, Pseudoristola, Pseudoeucyrtis, Stichocapsa, Yaocapsa and Zhamoidellum, which were not reported in other localities of the Piemont Schistes Lustrés zone (Bill et al. 2001). On one hand, these discrepancies may not be paleontologically significant as they could result from taphonomical biases. On the other hand, they could suggest that radiolarite beds stored as pebbles in molasse conglomerates have no in situ equivalents, or that these equivalents have been entirely destroyed or metamorphosed. We have presently no argument for preferring one or the other of these two hypotheses.
5 Discussion and conclusions
5.1 The Alpine Tethys birth and spreading scenario
The oldest radiolarite sample documented in our study is Late Bajocian or Early Callovian in age. If we consider that in situ radiolarites of the Western Alps are associated with ophiolite basements, this age remains compatible with the classical scenario of synchronous opening for both the Central Atlantic and the Alpine Tethys in the Middle Jurassic (Lemoine et al. 2000; Bill et al. 2001; Cordey and Bailly 2007; de Graciansky et al. 2010). However, we do not know if the sampled molasse units contain older oceanic remnants. In the outer Piemont zone, the three thrust-sheets bearing diagnostic supraophiolitic radiolarites are imbricated with thrust-sheets derived from the European distal margin (Caby et al. 1971; Tricart et al. 2003). The simplest hypothesis is that our pebbles result from the erosion of ophiolite-bearing thrust-sheets of a similar Piemont paleogeographic origin, i.e. the part of the ocean located at the foot of the European margin. However, one cannot rule out a source in one or several now entirely eroded units that originated from the other side of the ocean at the foot of the African margin (Adria).
With the contribution of these fossiliferous pebbles, the number of radiolarites dated so far in the Piemont zone of the Western Alps increases from 3 to 10. If the occurrence of Kimmeridgian or Tithonian cherts were confirmed by future sampling, it would extend the time span represented in these deep-sea sediments from 16 to 20 Myr. For an ocean with a very low spreading rate of ~2 cm/year as expected for the Alpine Tethys (Lagabrielle and Cannat 1990; Lagabrielle and Lemoine 1997; Lagabrielle 2009; Manatschal and Müntener 2009), this time span could correspond to 400 km in width. Of course this estimate is to be considered with caution, as pebbles could come from any level of an original radiolarite succession resting on a slightly older crust.
We do not have any evidence for the occurrence of radiolarite sedimentation in the Early Cretaceous and consequently an even wider oceanic domain. However, most Tethyan basin facies grade into pelagic limestones or shale/limestone alternation in the Late Tithonian (Bill et al. 2001). All direct and indirect fossiliferous elements of the ocean found in ophiolite covers and molasses are from the oldest parts of this domain, corresponding to remnants of the so-called “narrow ocean” stage. As already observed elsewhere in the Alpine–Apennine domain, radiolarite units are Middle and Late Jurassic without any evidence of Cretaceous chert sedimentation (Bill et al. 2001). Apart from the Balma and Chiavenna units in the Valais oceanic domain (93 Ma U/Pb ages; Liati and Froitzheim 2006), all other radiometric ages overlap the age of radiolarian cherts (Manatschal and Müntener 2009). In the hypothesis of ongoing Cretaceous rifting, elements of a younger and more axial part of this ocean remain to be identified. If such elements are no longer exposed, they could remain hidden at depth. They could also have been eroded after exhumation. If this is the case, further investigation on foreland detrital formations is needed. However, the most probable explanation is that during the Cretaceous, the Alpine Tethyan Ocean was no more active and the oceanic spreading was mostly accommodated in the Central Atlantic to the west and the Neotethys to the east.
5.2 The Alpine orogenic wedge building scenario
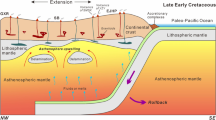
During Early Oligocene times, the present-day Alpine internal zones inherited part of the Eocene subduction-related accretionary wedge that had just undergone syncollisional polyphase shortening (Tricart et al. 2006, Morag et al. 2008, Bernet and Tricart 2011). The erosion of the new-born mountain range with exposed margin- and ocean-derived thrust sheets fed rather abruptly the foreland molasses which show a great variety of lithologies, not only radiolarites but also magmatic pebbles such as basalts and gabbros with greenschist to blueschist facies, as well as serpentinized peridotites with blueschist to eclogite facies (de Graciansky et al. 1971; Schwartz et al. 2012). As this foreland was progressively affected by folding and forethrusting, Oligocene molasse basins born along the Alpine front evolved as transported basins before being closed and incorporated to the Alpine wedge (present-day external zone: see review in Ford et al. 2006).
Although in situ fossiliferous radiolarian cherts of the outer Piemont zone underwent high greenschist or blueschist facies metamorphism, our study shows that some eroded radiolarite units buried as molasse pebbles experienced low P–T conditions. This is supported by the following observations: (1) low silica recrystallization of pebbles’s radiolarite matrices, (2) absence of phengites in the pebbles microfacies; these white micas are commonly observed in the HP blueschist siliceous facies of the Piemont zone, (3) fair preservation of radiolarian microfossils. It is also noteworthy that Mesozoic radiolarite pebbles of the Subalpine molasse display much higher rates of microfossil recovery than in situ Mesozoic radiolarites of the Piemont zone. Interestingly, the same low-grade metamorphism characterizes the chert pebbles extracted from synorogenic conglomerates derived from the Mesozoic accretionary complexes of western North America (Seiders and Blome 1984; Cordey 1992a, b, 1998). There, some radiolarite pebbles have revealed the existence of ocean units whose equivalent correlatives have never been found in situ (Cordey 1998). Our study suggests that this is also the case in the Alps.
The occurrence of low-grade radiolarite pebbles indicates that some eroded thrust-sheets originating from the oldest parts of the ocean escaped any serious tectonic burial during the Alpine convergence (Fig. 5). The 20 Myr potential age range represented by our radiolarite pebbles does not favour a single source. However, we do not know if these radiolarites derived from the European side or from both sides of the ocean spreading-ridge. Yet they show that by 30 Ma, the Alpine wedge had probably sampled the entire oceanic sediments accreted at very contrasted structural levels in the accretionary wedge. The question whether there is a link between the European or African paleogeographic origin of these remnants and their structural history remains open. Future models for the building of the Alpine wedge during early convergence will have to test the processes of a selective tectonic sampling of these paleogeographic realms during accretion.

Crustal cross-section model proposing an evolution for the southern Western Alps during the Early Oligocene. At that time, the Piemont thrust sheets, already stacked in the upper Cretaceous–Paleocene subduction accretionary wedge, underwent severe syncollision shortening. This resulted in a major orogenesis so that erosion fed abruptly the peripheral molasse basins in margin-derived and ocean-derived clasts whose metamorphic signatures range from non or very low-grade to high-grade blueschist facies (Lz lizardite, Atg antigorite)
References
Artoni, A., & Meckel, L. D. III., (1998). History and deformation rates of a thrust sheet top basin: The Barrême basin, western Alps, SE France. Geological Society Special Publication, 134, 213–237.
Barféty, J. C., Lemoine, M., de Graciansky, P. C., Tricart, P., & Mercier, D. (1995). Notice explicative, Carte géol. France (1/50 000), feuille Briançon (823), Orléans. Bureau de Recherches Géologiques et Minières. Geological map by J.C. Barféty, M. Lemoine, D. Mercier, R. Polino, P. Nievergelt, J. Bertrand, T. Dumont, S. Amaudric du Chaffaut, A. Pêcher, G. Montjuvent.
Baumgartner, P. O., Bartolini, A., Carter, E. S., Conti, M., Cortese, G., Danelian, T., De Wever, P., Dumitrica, P., Dumitrica-Jud, R., Gorican, S., Guex, J., Hull, D. M., Kito, N., Marcucci, M., Matsuoka, A., Murchey, B., O’Dogherty, L., Savary, J., Vishnevskaya, V., Widz, D., Yao, A. (1995). Middle Jurassic to Early Cretaceous radiolarian biochronology of Tethys based on unitary associations. In Baumgartner, P. O., et al. (Eds.), Middle Jurassic to Lower Cretaceous Radiolaria of Tethys: Occurrences, systematics, biochronology (Vol. 23, pp. 1013–1043). Lausanne: Mémoires de Géologie.
Bernet, M., & Tricart, P. (2011). The Oligocene orogenic pulse in the Southern Penninic Arc (Western Alps): Structural, sedimentary and thermochronological constraints. Bulletin de la Société Géologique de France, 182, 25–36. doi:10.2113/gssgfbull.182.1.25.
Bernoulli, D., & Jenkyns, H. (2009). Ancient oceans and continental margins of the Alpine–Mediterranean Tethys: Deciphering clues from Mesozoic pelagic sediments and ophiolites. Sedimentology, 56, 149–190.
Bill, M., O’Dogherty, L., Guex, J., Baumgartner, P. O., & Masson, H. (2001). Radiolarite ages in Alpine-Mediterranean ophiolites. Constraints on the oceanic spreading and the Tethys–Atlantic connection. Geological Society of America Bulletin, 113, 129–143.
Bousquet, R., Oberhänsli, R., Goffé, B., Wiederkehr, M., Koller, F., Schmid, S. M., Schuster, R., Engi, M., Berger, A., & Martinotti, G. (2008). Metamorphism of metasediments at the scale of an orogen: A key to the Tertiary geodynamic evolution of the Alps. In S. Siegesmund, B. Fügenschuh & N. Froitzheim (Eds.), Tectonic aspects of the Alpine–Dinaride–Carpathian System (pp. 393–411). The Geological Society of London.
Boussac, J. (1912). Etudes statigraphiques sur le Nummulitique alpin. Mémoire explicatif de la carte géologique détaillée de la France.
Caby, R., Michard, A., & Tricart, P. (1971). Découverte d’une brèche polygénique à éléments granitoïdes dans les ophiolites métamorphiques piémontaises (Schistes lustrés du Queyras, Alpes françaises). Comptes Rendus de l’Académie des Sciences Paris, 273, sér. D, 999–1002.
Chauveau, J. C., & Lemoine, M. (1961). Contribution à l’étude géologique du synclinal tertiaire de Barrême (moitié nord). Bulletin de la Carte géologique de France, 264, 147–178.
Cordey, F. (1992a). Radiolarians and Terrane analysis in the Canadian Cordillera: The “clastic approach”. In J. Aitchison & B. Murchey (Eds.), The significance and application of Radiolaria to Terrane analysis (Vol. 96, pp. 155–159). Palaeogeography Palaeoclimatology Palaeoecology.
Cordey, F. (1992b). Radiolarian ages from chert pebbles of the Tantalus Formation, Carmacks area, Yukon Territory. Geological Survey of Canada Paper, 92-1E, 53–59.
Cordey, F. (1998). Radiolaires des complexes d’accrétion cordillérains. Geological Survey of Canada Bulletin, 509.
Cordey, F., & Bailly, A. (2007). Alpine ocean seafloor spreading and onset of pelagic sedimentation: New radiolarian data from the Chenaillet-Montgenèvre ophiolite (French-Italian Alps). Geodinamica Acta, 20, 131–138.
de Graciansky, P. C. (1972). Le bassin tertiaire de Barrême (Alpes de Haute-Provence): Relations entre déformation et sédimentation; chronologie des plissements. Comptes Rendus de l’Académie des Sciences, Paris, 275, 2825–2828.
de Graciansky, P. C., Lemoine, M., & Saliot, P. (1971). Remarques sur la présence de minéraux et de paragénèses du métamorphisme alpin dans les galets des conglomérats oligocènes du synclinal de Barrême (Alpes de Haute-Provence). Comptes Rendus de l’Académie des Sciences, Paris, 272, 3243–3245.
de Graciansky, P. C., Roberts, D., & Tricart, P. (2010). The Western Alps from rift to passive margin to orogenic belt: An integrated geosciences overview. Amsterdam: Elsevier.
De Wever, P., & Baumgartner, P. O. (1995). Radiolarians from the base of the supra-ophiolitic Schistes Lustrés Formation in the Alps (Saint-Véran, France and Traversiera Massif, Italy). In P. O. Baumgartner, et al. (Eds.), Middle Jurassic to Lower Cretaceous Radiolaria of Tethys: Occurrences, systematics, biochronology (Vol. 23, pp. 725–730). Lausanne: Mémoires de Géologie.
De Wever, P., & Caby, R. (1981). Datation de la base des Schistes lustrés postophiolitiques par des radiolaires (Oxfordien supérieur-Kimméridgien moyen) dans les Alpes Cottiennes (Saint-Véran, France). Comptes Rendus de l’Académie des Sciences Paris, 292, 467–472.
De Wever, P., Danelian, T., Durand-Delga, M., Cordey, F., & Kito, N. (1987). Datations des radiolarites post-ophiolitiques de Corse alpine à l’aide des Radiolaires. Comptes Rendus de l’Académie des Sciences Paris, 305, 893–900.
Evans, M. J., & Elliott, T. (1999). Evolution of a thrust-sheet-top basin: The Tertiary Barrême basin, Alpes de Haute Provence, France. Geological Society of America Bulletin, 111, 1617–1643.
Ferry, S., Tremblay, S., Guiraud, M., & Cordey, F. (2005). Les molasses rouges paléogènes du bassin subalpin français. Corrélations avec le bassin helvétique. ASF Association Sédimentologie France, Octobre 2005 (abstract).
Ford, M., Duchene, S., Gasquet, D., & Vanderhaeghe, O. (2006). Two-phase orogenic convergence in the external and internal SW Alps. Journal of the Geological Society, 163, 815–826.
Ford, M., & Lickorish, W. H. (2004). Foreland basin evolution around the western Alpine arc. In P. Joseph & S. A. Lomas (Eds.), Deep-water sedimentation in the Alpine basin of SE France: new perspectives on the grès d’Annot and related systems (Vol. 221, pp. 39–63). Geological Society Special Publication.
Ford, M., Lickorish, W. H., & Kusznir, N. J. (1999). Tertiary foreland sedimentation in the Southern Subalpine Chains, SE France: A geodynamic appraisal. Basin Research, 11, 315–336.
Gidon, M. (1971). Gap, geological map of France 1:50 000. Bureau de Recherches Géologiques et Minières.
Gidon, M., Kerckhove, C., Michard, A., Tricart, P., Gotteland, P., Gout, C., Leblanc, D., Lefevre, R., Le Guernic, J., Mégard-Galli, J., & Michel-Noël, G. (1994). Aiguille de Chambeyron, Geological Map of France 1/50 000. Bureau de Recherches Géologiques et Minières.
Gubler, Y. (1958). Etude critique des sources du matériel constituant certaines séries dans le Tertiaire des Alpes françaises du Sud: Formations détritiques de Barrême, Flysch “Grés d’Annot”. Eclogae Geologicae Helvetiae, 51, 942–977.
Lagabrielle, Y. (2009). Mantle exhumation and lithospheric spreading: An historical perspective from investigations in the oceans and in the Alps–Appennines ophiolites. Italian Journal of Geosciences, 128, 279–293.
Lagabrielle, Y., & Cannat, M. (1990). Alpine Jurassic ophiolites resemble the modern central Atlantic basement. Geology, 18, 319–322.
Lagabrielle, Y., & Lemoine, M. (1997). Alpine, Corsican and Apennine ophiolites: The slow-spreading ridge model. Comptes Rendus de l’Académie des Sciences, Paris, 325, sér. II, 909–920.
Lemoine, M. (1983). Rifting and early drifting: Mesozoic Central Atlantic and Ligurian Tethys. Initial reports Deep Sea Drilling Project, US Government Printing Office, Washington DC, 76, 885–895.
Lemoine, M., Bas, T., Arnaud-Vanneau, A., Arnaud, A., Dumont, T., Gidon, M., Bourbon, M., de Graciansky, P.-C., Rudkiewicz, J.-L., Mégard-Galli, J., & Tricart, P. (1986). The continental margin of the Mesozoic Tethys in the Western Alps. Marine and Petroleum Geology, 3, 179–199.
Lemoine, M., de Graciansky, P. C., & Tricart, P. (2000). De l’océan à la chaîne de montagnes: Tectonique des plaques dans les Alpes. Paris: Gordon and Breach Science.
Lemoine, M., Steen, D., & Vuagnat, M. (1970). Sur le problème stratigraphique des ophiolites piémontaises et des roches sédimentaires associées: Observations dans le massif de Chabrière en Haute-Ubaye (Basses-Alpes, France). Archives des Sciences, Genève, 5, 44–59.
Liati, A., & Froitzheim, N. (2006). Assessing the Valais Ocean, Western Alps: U-Pb SHRIMP zircon geochronology of eclogite in the Balma unit, on top of the Monte Rosa nappe. European Journal of Mineralogy, 18, 299–308.
Manatschal, G., & Müntener, O. (2009). A type sequence across an ancient magma-poor ocean-continent transition: The example of the western Alpine Tethys ophiolites. Tectonophysics, 437, 4–19.
Meckel, L. D. III, Ford, M., & Bernoulli, D. (1996). Tectonic and sedimentary evolution of the Dévoluy Basin, a remnant of the Tertiary western Alpine foreland basin, SE France. Géologie de la France, 2, 3–26.
Mevel, C., Caby, R., & Kienast, J.-R. (1978). Lower amphibolite facies conditions in the oceanic cruts: Example of amphibolitized flaser-gabbro and amphibolites from an alpine ophiolitic massif (Chenaillet massif, Hautes Alpes, France). Earth and Planetary Science Letters, 39, 98–108.
Morag, N., Avigad, D., Harlavan, Y., McWilliams, O., & Michard, A. (2008). Rapid exhumation and mountain building in the Western Alps: Petrology and 40Ar/39Ar geochronology of detritus from Tertiary basins of southeastern France. Tectonics, 27. doi:10.1029/2007TC002142.
O’Dogherty, L., Bill, M., Gorican, S., Dumitrica, P., & Masson, H. (2006). Bathonian radiolarians from an ophiolitic mélange of the Alpine Tethys (Gets Nappe, Swiss-French Alps). Micropaleontology, 51(6), 425–485.
Rüst, D. (1885). Beiträge zur Kenntniss der fossilen Radiolarien aus Gesteinen des Jura und der Kreide. Palaeontographica, 31, 269–321.
Schaaf, A., Polino, R., & Lagabrielle, Y. (1985). Nouvelle découverte de radiolaires d’âge Oxfordien supérieur-Kimméridgien inférieur à la base d’une série supra-ophiolitique des schistes lustrés piémontais (Massif de Traversiera, Haut Val Maïra, Italie). Comptes Rendus de l’Académie des Sciences, Paris, 14, sér. II, 1079–1084.
Schmid, S. M., Fügenschuh, B., Kissling, E., & Schuster, R. (2004). Tectonic map and overall architecture of the Alpine orogen. Eclogae Geologicae Helvetiae, 97, 93–117.
Schwartz, S., Tricart, P., Guillot, S., Bernet, M., Chamorro-Perrez, E. M., Montagnac, G., & Jourdan, S. (2012). Source tracing of detrital serpentinite in the Oligocene molasse deposits from the western Alps (Barrême basin): Implications for relief formation in the internal zone. Geological Magazine. doi:10.1017/S0016756811001105
Seiders, V. M., & Blome, C. D. (1984). Clast compositions of Upper Mesozoic conglomerates of the California Coast Ranges and their tectonic significance. In M.C. Blake Jr. (Ed.), Franciscan geology of Northern California (Vol. 43, pp. 135–148). Pacific Section of Economic Paleontologists and Mineralogists.
Sissingh, W. (2001). Tectonostratigraphy of the West Alpine Foreland: Correlation of Tertiary sedimentary sequences, changes in eustatic sea-level and stress regimes. Tectonophysics, 333, 361–400.
Tricart, P. (1984). From passive margin to continental collision: A tectonic scenario for the Western Alps. American Journal of Science, 284, 97–120.
Tricart, P., Amaudric du Chaffaut, S., Ayoub, C., Ballèvre, M., Caby, R., Gout, C., Lagabrielle, Y., Leblanc, D., Le Mer, O., Philippot, P., & Saby, P. (2003). Aiguilles-Col Saint Martin Geological Map of France 1/50 000. Bureau de Recherches Géologiques et Minières.
Tricart, P., Lardeaux, J.-M., Schwartz, S., & Sue, C. (2006). The late extension in the inner western Alps: A synthesis along the south-Pelvoux transect. Bulletin de la Société Géologique de France, 177, 299–310.
Acknowledgments
We thank Pierre-Charles de Graciansky (Paris) and Peter O. Baumgartner (Lausanne) for their thorough and insightful reviews. This work was supported by the CNRS-UMR 5276 and the ANR program “ERD-Alps” (Erosion and Relief Development in the Western Alps). Chert pebbles were processed in the radiolarian extraction laboratory of Université Lyon 1 (CNRS-UMR 5276). We also thank Arlette Armand (Lyon) for SEM assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial Handling: D. Marty.
The studied pebbles are housed at the Department of Earth Sciences of University Claude Bernard Lyon 1 (France).
Rights and permissions
About this article
Cite this article
Cordey, F., Tricart, P., Guillot, S. et al. Dating the Tethyan Ocean in the Western Alps with radiolarite pebbles from synorogenic Oligocene molasse basins (southeast France). Swiss J Geosci 105, 39–48 (2012). https://doi.org/10.1007/s00015-012-0090-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00015-012-0090-8