
Abstract
We describe a new genus and species of woodpecker (Piciformes: Picidae), Piculoides saulcetensis, from the early Miocene (MN1–MN2) of Saulcet, in the “Saint-Gérand-le-Puy” area, central France, which is the earliest definite record of the family. The new species is represented solely by the distal end of a tarsometatarsus, which bears nonetheless diagnostic features that allowed us to place Piculoides saulcetensis in a phylogenetic context. Our results show that the fossil from Saulcet is either a stem-group representative of piculets (Picumninae) and true woodpeckers (Picinae) or of true woodpeckers only. Piculoides saulcetensis is similar to a fragmentary tarsometatarsus of a picid from the late Oligocene of southern Germany, and we thus hypothesize a close relationship between the two.
Similar content being viewed by others


Introduction
Picidae (wrynecks, piculets, and true woodpeckers) are the most species-rich family of the Piciformes, and are classified in the taxon Pici, together with Ramphastidae (barbets and toucans), and Indicatoridae (honeyguides). Monophyly of the Pici is well supported both by anatomical (Simpson and Cracraft 1981; Swierczewski and Raikow 1981; Mayr et al. 2003) and molecular character sets (e.g., Johansson and Ericson 2003; Mayr et al. 2003). Hypotheses on the relationships between the major groups within Picidae are well supported by morphological and molecular data (e.g., Goodge 1972; Swierczewski and Raikow 1981; Short 1982; Webb and Moore 2005; Benz et al. 2006). It has been the long-standing view that Picumninae (piculets) and Jynginae (wrynecks) are successive sister taxa of Picinae (true woodpeckers).
The fossil record of woodpeckers is sparse. The earliest fossil specimen of Pici recovered so far dates to the early Oligocene (MP 21) of Belgium (Mayr and Smith 2001), and is represented by a distal tarsometatarsus. Another early Oligocene species, Rupelramphastoides knopfi, was also found in southern Germany, but whether this species belongs within crown-group Pici is uncertain (Mayr 2005, 2006). Further undisputed members of the Pici, all based on incomplete tarsometatarsi, have been reported from the late Oligocene (MP 29) and late Oligocene/early Miocene of Germany (Mayr 2001, 2010). The Miocene record of Pici in Europe is mostly restricted to the Ramphastidae (toucans and barbets), and more explicitly to the extinct taxon Capitonides, from the early and middle Miocene of Germany (Ballmann 1969, 1983).
The maybe earliest record of the Picidae is a contour feather preserved in amber from the Dominican Republic, which was assigned to this family by Laybourne et al. (1994), but its precise age, though probably early Miocene at least, is nevertheless unknown. An ulna from the very late Miocene (Messinian) of Italy was described by Ballmann (1976) as belonging in this family. Picus peregrinabundus (Umanskaja 1981) from the late Miocene of the Ukraine, described from a distal tarsometatarsus, was considered a true woodpecker (Picinae) by Umanskaja (1981), although the illustrations provided do not allow for a corroboration of her hypothesis. In North America, Pliopicus brodkorbi from the late Miocene of Kansas (see Becker 1986) was described based on a distal tarsometatarsus (Feduccia and Wilson 1967). Though Feduccia and Wilson (1967) argued a close relationship between Pliopicus and the extant woodpecker genus Melanerpes, Cracraft and Morony (1969) judged the specimen to be markedly different from recent woodpeckers. The description of Palaeonerpes shorti from the middle/late Miocene of Nebraska was based on a distal tibiotarsus, and Cracraft and Morony (1969) suggested possible close relationships to melanerpine woodpeckers (tribe Dendropicini, Picinae). An ulna, from the middle Miocene of Nebraska (see Korth 2008 for stratigraphy), was described by Wetmore (1931), who highlighted a strong resemblance to colaptine woodpeckers (flickers, Picinae); these affinities have been since then seconded by Short (1965).
Despite large amounts of fossil bird material, no representatives of Picidae have so far been described from the “Saint-Gérand-le-Puy” area (Allier, France; late Oligocene to early Miocene); Picus archiaci and P. consobrinus were assigned to the Picidae by Milne-Edwards (1867–1868), but were subsequently identified as representatives of Coliiformes by Ballmann (1969). In the comparatively small collection of avian bones from Saulcet, “Saint-Gérand-le-Puy”, housed in the Natural History Museum Basel, Switzerland, we have, however, found a well-preserved distal end of a tarsometatarsus of a member of the Picidae, which we describe in this study. Despite its fragmentary nature, the unique combination of its characters justifies the introduction of a new genus and species of woodpecker. Because features of the distal end of this bone are diagnostic, we attempt to clarify the phylogenetic affinities of the fossil specimen, and compare it with the aforementioned fossil members of the Pici.
Geological setting
The localities from the Saint-Gérand-le-Puy area in the Allier Basin, including Saulcet (Fig. 1), have a long history of stratigraphic problems, since old museum collections, housed in various institutions around the world, are composed of specimens with unknown precise provenience and thus unknown precise stratigraphy, with several localities being gathered under the common name of “Saint-Gérand-le-Puy”. This is not the case for the material of Saulcet from the Natural History Museum Basel, from which the specimen described below comes. Fossils from the locality were collected by Hürzeler, a former curator of the Basel Natural History Museum, in the 1930s and formed the basis of his seminal work on cainotheres (Hürzeler 1936).

The Allier sedimentary basin is known for its late Oligocene/early Miocene calcareous sediments deposited in a fluvio-lacustrine context (Donsimoni 1975). These sediments yielded tens of localities with fossil vertebrates and invertebrates, several of these playing an important role for the European continental mammal biochronological time scale (the MN zones; Mein 1975), such as Montaigu-le-Blin, the reference locality for zone MN2. Hürzeler collected hundreds of fossils most probably from a single quarry known as Pénaud (Hugueney 1974), where the “calcaires à Phryganes” (insect sheaths-rich lacustrine limestones, Donsimoni 1975) crop out together with stromatolitic limestones. Indeed, this is where the fossils were found, trapped between the bioconstructions (Hugueney 1974; Berthet, (2003)). Despite an abundant fossil collection not much was published regarding the geology and age of Saulcet. Some uncertainty long surrounded the age of this fauna. Hürzeler (1944) proposed a late Aquitanian age, and later Hugueney (1974) suggested a very early MN2 age (“very base of the under-zone containing Montaigu-le-Blin”, i.e., MN2a). A more recent biochronological assignment points to a late MN1 to early MN2 age (Hugueney 1997; Fig. 1), the MN1 designation being accepted by most authors (e.g., Hugueney 1984; Ziegler 1999; Berthet 2003).
Materials and methods
Institutional abbreviations: NMB, Natural History Museum Basel, Switzerland; NMNH, Smithsonian Institution, National Museum of Natural History, Washington, USA; SMF, Forschungsinstitut Senckenberg, Frankfurt am Main, Germany.
The fossil specimen is deposited in NMB. Anatomical terminology follows Baumel and Witmer (1993). The following skeletons of recent Piciformes were available for comparisons: Picidae: Picinae: Colaptes auratus (SMF), Chrysocolaptes lucidus (SMF), Dendrocopos major (NMB), D. minor (SMF), Dendropicos fuscescens (SMF), Dinopium javanense (SMF), Dryocopus martius (SMF), Hemicircus concretus (SMF); Melanerpes carolinus (SMF), Picoides tridactylus (NMB), Picus canus (NMB), P. viridis (NMB), Sphyrapicus varius (SMF); Picumninae: Picumnus albosquamatus (SMF), Sasia abnormis (NMNH); Jynginae: Jynx torquilla (NMB); Indicatoridae: Indicator variegatus (SMF); Ramphastidae: Trachyphonus margaritatus (NMB); Pteroglossus viridis (NMB).
Systematic paleontology
Aves Linnaeus, 1758
Piciformes Meyer & Wolf, 1810
Pici sensu Simpson & Cracraft (1981)
Picidae Vigors, 1825
Piculoides gen. nov.
Type species: Piculoides saulcetensis, sp. nov.
Etymology: Piculoides, piculet-like.
Differential diagnosis: small member of Picidae, which differs from:
-
Ramphastidae and Indicatoridae in: (1) trochlea metatarsi II proximodistally elongated (reaching distal end of trochlea metatarsi III); (2) strongly asymmetric rims of trochlea metatarsi III with medial rim projecting further plantar than lateral rim; (3) proximodistally short trochlea metatarsi III; (4) groove in trochlea metatarsi III deep; (5) trochlea metatarsi III with slanting rims (Ballmann 1969, p. 42, fig. 11-1);
-
Ramphastidae in: (6) trochlea accessoria rests plantar of trochlea metatarsi III; (7) dorso-plantarly short trochlea metatarsi III; (8) deep groove between trochlea metatarsi IV and trochlea accessoria for tendon of M. flexor hallucis longus pars dig. IV (Ballmann 1969, p. 42, fig. 11-2); (9) medioplantar bulge just proximal of trochlea metatarsi III absent (Mayr 1998, p. 50ff, fig. 28C, D);
-
Jynginae in: (1) strongly asymmetric rims of trochlea metatarsi III with medial rim projecting further plantar than lateral rim; (2) proximodistally short trochlea metatarsi III
-
Picumninae in: (1) very wide trochlea metatarsi IV; (2) considerably larger size; (3) trochlea accessoria not as thickened.
-
Picinae in: (1) trochlea metatarsi II narrow; (2) articular surface of trochlea metatarsi IV not plantarly enlarged; (3) articular surface of trochlea metatarsi IV not set-off from rest of trochlea.
The new taxon differs from all examined Pici in the very wide notch for the tendon of M. extensor digitorum longus pars dig. IV and the distinctly splayed trochlea metatarsi IV, which are here considered autapomorphic features of the new taxon.
Piculoides saulcetensis, sp. nov.
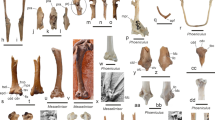
Fig. 2a, b, g and l.

Left distal tarsometatarsus of Piculoides saulcetensis gen. et sp. nov. (NMB Sau.4975) from the early Miocene of Saulcet, in comparison to extant and fossil Pici. a–b, Piculoides saulcetensis gen. et sp. nov., in plantar and dorsal views, respectively; dorsal views of the tarsometatarsi of Jynx torquilla (c), Picumnus albosquamatus (d), Picus viridis (e), and Trachyphonus margaritatus (f); distal views of the tarsometatarsi of Piculoides saulcetensis (g), Jynx torquilla (h), Picumnus albosquamatus (i), Picus viridis (j), and Trachyphonus margaritatus (k); lateral views of the tarsometatarsi of Piculoides saulcetensis (l), Jynx torquilla (m), Picumnus albosquamatus (n), Picus viridis (o), and Trachyphonus margaritatus (p); right tarsometatarsus of SMF Av. 429, from the late Oligocene of Germany, in distal (q, not to scale), and dorsal (r) views. We point out that trochlea accessoria of SMF Av. 429 is badly damaged, and that the edges of trochlea metatarsi III are worn. Abbreviations: art. surf. tr. met. IV, articular surface of trochlea metatarsi IV; sul. ten. M. ext. digit. long, sulcus for the tendon of musculus extensor digitorum longus pars dig. IV; tr. acc., trochlea accessoria; tr. met. II, trochlea metatarsi II; tr. met. III, trochlea metatarsi III; tr. met. IV, trochlea metatarsi IV. Images c, d, f, h, I, k, m, n, p have been mirrored to facilitate comparisons. Scale bars are 2 mm, except g–k and q, 1 mm. (Photographs d, i, n, q and r by Sven Tränkner, Senckenberg, Frankfurt)
Etymology: saulcetensis, named after the locality of Saulcet.
Holotype: distal end of left tarsometatarsus, NMB Sau.4975.
Locality and age: Saulcet, Allier, France; early Miocene, Aquitanian (MN1–base of MN2, ca. 22.5 Ma).
Measurements: distal width: 3.0 mm.
Diagnosis: same as for genus. Piculoides saulcetensis matches the measurements of the distal end of the tarsometatarsus of the extant small true woodpecker species Dendrocopos minor and Dendropicos fuscescens, and is only slightly smaller than Jynx torquilla. All these species are considerably larger than piculets (Table 1).
Description and comparisons
As in recent Picidae, the foramen vasculare distale is very small; a second, medial foramen, which is present in many true woodpeckers (e.g., Dendrocopos, Dryocopus, Picus), is not visible. The presence of this second foramen is nevertheless variable within modern Picidae. The dorsal surface of the distal end is flat, which is characteristic for Pici (Ballmann 1983). The distal end is not as proximodistally compressed as in extant woodpeckers (Picoides particularly exhibiting this condition), being somewhat intermediate between the condition in true woodpeckers and Jynx, in which the trochleae metatarsorum project more distally.
Although the fossa metatarsi I is distinct, it is not nearly as pronounced as in Picumnus or Trachyphonus, in which the medial rim is visible even in dorsal view. This is also the case in Jynx torquilla as well as in those Picinae which bear a comparatively large hallux, e.g., Colaptes auratus. Therefore, it seems unlikely that the hallux was greatly or even completely reduced in Piculoides, as opposed to the condition in Sasia (Picumninae), Gecinulus, and certain species of Dendrocopos, Dinopium and Picoides (Short 1982; Winkler and Christie 2002).
Contrary to the arrangement in Ramphastidae, the trochlea metatarsi II is proximodistally elongated. It departs, nonetheless, from the condition found in true woodpeckers in that it is comparatively slender, which also applies to the piculets Picumnus albosquamatus and Sasia abnormis, and to Jynx torquilla, whereas it is thick in the examined representatives of the Picinae. The new taxon is also characterized by the very deep and slender incisura intertrochlearis medialis, which is related to the greater length of the trochlea metatarsi II. This feature is also present in piculets, particularly in Sasia abnormis, and—to a lesser degree—in Jynx torquilla.
The trochlea metatarsi III is proximodistally shorter than in Jynx (Fig. 2g–h), and, as in recent Picidae, the groove in trochlea metatarsi III is deep. Both features are synapomorphic for Picumninae and Picinae. Characteristic for Picidae (and to a lesser extent Indicatoridae) is also a mediolaterally slanting trochlea metatarsi III (Ballmann 1969), which is present in the new taxon. The rims of trochlea metatarsi III are distinctly asymmetric (Fig. 2g), a feature in which Piculoides is indistinguishable from Picumnus albosquamatus and Sasia abnormis. Although the medial rim reaches farther plantar in all woodpeckers, this asymmetry is particularly noticeable in the fossil specimen and piculets, but also in some true woodpeckers as Meiglyptes tukki (Antoine Louchart, personal communication) and Chrysocolaptes lucidus. In lateral view it can be appreciated that trochlea metatarsi III is more dorsally protruding than in Picinae, and thus closely resembles Jynx (Fig. 2l–m).
In dorsal view, the trochlea metatarsi IV is peculiarly wide; its unusual shape being different from that in other Pici including Jynginae, Picumninae and Picinae. In Piculoides this trochlea spreads out and is clearly set off from the tarsometatarsus, which is not the case in the examined recent specimens. This feature could ultimately be related to the very wide, saddle-shaped notch proximal to this trochlea, namely the pulley for the tendon of M. extensor digitorum longus pars dig. IV. Unlike in Jynx, the trochlea metatarsi IV (Fig. 2g) bulges dorsally in distal view; this condition is also observed in piculets and true woodpeckers. The articular surface of trochlea metatarsi IV (Fig. 2l) is not as thickened as in Picinae, and it closely resembles that of Jynx and Picumnus. The articular facet is not set-off from the rest of the trochlea as in Picinae, thus exhibiting the plesiomorphic condition present in all other Pici.
Although the distal end of the trochlea accessoria is slightly damaged, a strong distal projection (Fig. 2l) matching the one present in Picumnus (Fig. 2n) and Sasia is noticeable. The trochlea accessoria does not project distinctly further distal than trochlea metatarsi III in Trachyphonus, Indicator, Jynx, and most Picinae (the closely related Dendrocopos, Dendropicos, Sphyrapicus are an exception, but the trochlea accessoria is still comparatively shorter than in the more basal Picidae). Trochlea accessoria and trochlea metatarsi IV are separated by a proximodistally deep groove in Picidae and Indicatoridae, which is also present in the fossil. The sulcus for the tendon of M. flexor perforatus digiti IV at the medial surface of the trochlea accessoria is noticeably notched, not differing from Indicatoridae and Picidae. This feature is less concave in ramphastids, e.g., Trachyphonus margaritatus.
Discussion
Piculoides saulcetensis can be assigned to the Picidae by the following derived features of the tarsometatarsus (Fig. 3): (1) presence of a deeply grooved trochlea metatarsi III with mediolaterally slanting rims, (2) proximodistally elongated trochlea metatarsi II, which reaches the distal end of the trochlea metatarsi III, (3) trochlea accessoria longer than trochlea metatarsi III (it is shorter outside crown-group Pici; see Mayr 1998). The fossil is distinguished from Jynginae and agrees with Picumninae and Picinae in the proximodistally short trochlea metatarsi III and in the strongly asymmetric rims of trochlea metatarsi III in distal view, which is absent from most, but not all Picinae (see description).

Phylogenetic tree based on Benz et al. (2006), showing the position of Piculoides saulcetensis in relation to extant Pici. Nodes are supported by the following osteological characters of the distal tarsometatarsus: 1 trochlea accessoria rests plantar of trochlea metatarsi III, dorso-plantarly short trochlea metatarsi III, deep groove between trochlea metatarsi IV and trochlea accessoria for tendon of m. flexor hallucis longus pars dig. IV, plantar bulge just proximal of trochlea metatarsi III absent, trochlea metatarsi III with slanting rims; 2 trochlea metatarsi II proximodistally elongated, groove in trochlea metatarsi III deep; 3 strongly asymmetric rims of trochlea metatarsi III in distal view, proximodistally short trochlea metatarsi III; 4 trochlea metatarsi II wide, articular surface of trochlea metatarsi IV plantarly enlarged, articular surface of trochlea metatarsi IV set-off from rest of trochlea
The resemblances between the fossil taxon and extant Picumninae (Picumnus and Sasia) are based on presumably plesiomorphic traits for Picumninae and Picinae, such as the strongly asymmetric rims of trochlea metatarsi III and a trochlea accessoria projecting further distally. Piculoides lacks derived traits present in Picinae, such as a thick trochlea metatarsi II, a plantarly enlarged articular surface of trochlea metatarsi IV, and a trochlea metatarsi IV with an articular surface set-off from the rest of the trochlea. For these reasons we consider it likely for Piculoides to be outside crown-group Picinae, but a larger sample of extant Picidae would need to be examined for additional support of this hypothesis.
We hypothesize that Piculoides is either a stem-lineage representative of both piculets and woodpeckers, or of true woodpeckers, which is evidenced by the mosaic of tarsometatarsal characters showing features present in both Picinae and Picumninae (Fig. 2). We nevertheless approach the evidence with caution, given that Piculoides is only known from a distal tarsometatarsus and more osteological features remain to be discovered and detailed in order to procure an unambiguous and well-supported phylogenetic placement. In any case, no true woodpeckers (Picinae) are known to have existed in Europe during the early Miocene, and on a global scale the only Miocene records of members of the Picinae are restricted to single specimens from the middle to late Miocene (Wetmore 1931; Feduccia and Wilson 1967; Cracraft and Morony 1969; Umanskaja 1981).
Piculoides saulcetensis represents the earliest definite record of the Picidae. We note, however, that a late Oligocene partial tarsometatarsus (SMF Av. 429) reported by Mayr (2001) agrees in size (see also Table 1) and comparable osteological traits with Piculoides. Particularly, they both display strongly asymmetric rims of trochlea metatarsi III, which, as previously mentioned, are also characteristic for Picumnus and Sasia. It is thus conceivable that this late Oligocene specimen is closely related to Piculoides, and may belong in the same genus. The other late Oligocene/early Miocene specimen recently reported by Mayr (2010) is slightly larger than Piculoides, and because it probably possessed a very narrow trochlea metatarsi II, it is unlikely that it represents a true woodpecker (Picinae). Due to incomplete preservation of this specimen, other comparisons with Piculoides are not practicable. The piculet-sized bird from the early Oligocene of Belgium described by Mayr and Smith (2001) possesses features consistent with its belonging outside of crown-group Picidae: it bears only slightly asymmetric trochlea metatarsi III, with a shallow furrow. Picus peregrinabundus (Umanskaja 1981) appears to be almost twice the size of Piculoides, and is morphologically quite different from the Saulcet specimen. Pliopicus brodkorbi Feduccia and Wilson 1967 is slightly larger than Piculoides, and judging from the illustrations provided by Feduccia and Wilson (1967) probably belongs in the Picinae. It lacks, however, the strongly asymmetric rims of trochlea metatarsi III which are characteristic for Piculoides and which are likely to be plesiomorphic for Picumninae and Picinae. The unusual mediolateral compression of the distal end of P. brodkorbi is not unlike that of Jynx torquilla, to which the specimen bears an overall resemblance, but not much can be ascertained without proper re-examination of the specimen.
Although Piculoides shows a combination of characters of the foot existent in both Picumninae and Picinae, the unusual, laterally splayed trochlea metatarsi IV and the wide sulcus for the tendon of M. extensor digitorum longus pars dig. IV could indicate that Piculoides was more terrestrial than most of these taxa, given that this peculiarity is also present to some extent in the extant genus of barbets Trachyphonus, which are notoriously more terrestrial than other barbets. This hypothesis, nevertheless, cannot be corroborated without a proper comparative study.
References
Ballmann, P. (1969). Die Vögel aus der altburdigalen Spaltenfüllung von Wintershof (West) bei Eichstätt in Bayern. Zitteliana, 1, 5–60.
Ballmann, P. (1976). Fossile Vögel aus dem Neogen der Halbinsel Gargano (Italien), zweiter Teil. Scripta Geologica, 38, 1–59.
Ballmann, P. (1983). A new species of fossil barbet (Aves: Piciformes) from the Middle Miocene of the Nördlinger Ries (Southern Germany). Journal of Vertebrate Paleontology, 3(1), 43–48.
Becker, J. J. (1986). Fossil birds of the Oreana local fauna (Blancan), Owyhee County, Idaho. Great Basin Naturalist, 46(4), 673–678.
Benz, W. B., Robbins, M. B., & Peterson, A. T. (2006). Evolutionary history of woodpeckers and allies (Aves: Picidae): placing key taxa on the phylogenetic tree. Molecular Phylogenetics and Evolution, 40, 389–399.
Berthet, D. (2003). Le genre Cainotherium (Mammalia, Artiodactyla). Etude morphométrique, révision systématique, implications évolutives et paléogéographiques, extinction. Documents des laboratoires de Géologie de Lyon, 159, 205.
Cracraft, J., & Morony, J. J., Jr. (1969). A new Pliocene woodpecker, with comments on the fossil Picidae. American Museum Novitates, 2400, 1–8.
Donsimoni, M. (1975). Etude des calcaires concrétionnés lacustres de l’Oligocène supérieur et de l’Aquitanien du bassin de Limagne (Massif Central, France). Unpublished thesis, University Paris VI, 201 p.
Feduccia, J. A., & Wilson, R. L. (1967). Avian fossils from the lower Pliocene of Kansas. Occasional Papers of the Museum of Zoology, University of Michigan, 655, 1–6.
Goodge, W. R. (1972). Anatomical evidence for phylogenetic relationships among woodpeckers. Auk, 89, 65–85.
Gradstein, F. M., Ogg, J. G., Smith, A. G., Bleeker, W., & Lourens, L. J. (2004). A new geologic time scale, with special reference to Precambrian and Neogene. Episodes, 27, 83–100.
Hugueney, M. (1974). Gisements de petits mammifères dans la région de Saint-Gérand-le-Puy (stratigraphie relative). Revue scientifique du Bourbonnais, 52–68.
Hugueney, M. (1984). Evolution du Paléoenvironnement dans le tertiaire de Limagne (Massif Central, France) à partir des faunes de mammifères. Geobios, mémoire spécial no. 8, 385–391.
Hugueney, M. (1997). Biochronologie mammalienne dans le Paléogène et le Miocène inférieur du centre de la France : synthèse réactualisée. In J.-P. Aguilar, S. Legendre and J. Michaux (Eds), Actes du Congrès BiochroM’97 (pp. 417–430). Mémoires et Travaux de l’institut de Montpellier, 21: E.P.H.E.
Hürzeler, J. (1936). Osteologie und Odontologie der Caenotheriden. Schweizerische Paläontologische Abhandlungen, 58.
Hürzeler, J. (1944). Beiträge zur Kenntnis der Dimylidae. Schweizerische Paläontologische Abhandlungen, 65.
Johansson, U. S., & Ericson, P. G. P. (2003). Molecular support for a sister group relationship between Pici and Galbulae (Piciformes sensu Wetmore 1960). Journal of Avian Biology, 34, 185–197.
Korth, W. W. (2008). Cranial morphology, systematics and succession of beavers from the middle miocene valentine formation of nebraska, USA. Acta Palaeontologica Polonica, 53(2), 169–182.
Laybourne, R. C., Deedrick, D. W., & Hueber, F. M. (1994). Feather in amber is earliest new world fossil of Picidae. Wilson Bulletin, 106(1), 18–25.
Mayr, G. (1998). “Coraciiforme” und “piciforme” Kleinvögel aus dem Mittel-Eozän der Grube Messel (Hessen, Deutschland). Courier Forschungsinstitut Senckenberg, 205, 1–101.
Mayr, G. (2001). The earliest fossil record of a modern-type piciform bird from the late Oligocene of Germany. Journal für Ornithologie, 142, 2–6.
Mayr, G. (2005). A tiny barbet-like bird from the Lower Oligocene of Germany: the smallest species and earliest substantial fossil record of the Pici (woodpeckers and allies). Auk, 122(4), 1055–1063.
Mayr, G. (2006). First fossil skull of a Paleogene representative of the Pici (woodpeckers and allies) and its evolutionary implications. Ibis, 148, 824–827.
Mayr, G. (2010). Mousebirds (Coliiformes), parrots (Psittaciformes), and other small birds from the late Oligocene/early Miocene of the Mainz Basin, Germany. Neues Jahrbuch für Geologie und Paläontologie–Abhandlungen, 258(2), 129–144.
Mayr, G., Manegold, A., & Johansson, U. S. (2003). Monophyletic groups within ‘higher land birds’––comparison of morphological and molecular data. Journal of Zoological Systematics and Evolutionary Research, 41, 233–248.
Mayr, G., & Smith, R. (2001). Ducks, rails, and limicoline waders (Aves: Anseriformes, Gruiformes, Charadriiformes) from the lowermost Oligocene of Belgium. Geobios, 34(5), 547–561.
Mein, P. (1975). Proposition de biozonation du Néogène méditterranéen à partir des Mammifères. Actas i coloquio internacional sobre biostratigrafia continental del Neogeno superior y cuaternario inferior, 4, 112–113.
Meulenkamp, J. E., & Sissingh, W. (2003). Tertiary palaeogeography and tectonostratigraphic evolution of the northern and southern Peri-Tethys platforms and the intermediate domains of the African-Eurasian convergent plate boundary zone. Palaeogeography, Palaeoclimatology, Palaeoecology, 196, 209–228.
Milne-Edwards, A. (1867–1868). Recherches anatomiques et paléontologiques pour servir à l’histoire des oiseaux fossiles de la France, vol II., Paris: Victor Masson et fils.
Short, L. L. (1965). Variation in West Indian flickers (Aves, Colaptes). Bulletin of the Florida State Museum, 10, 1–42.
Short, L. L. (1982). Woodpeckers of the world. Greenville, Delaware: Delaware Museum of Natural History.
Simpson, S. F., & Cracraft, J. (1981). The phylogenetic relationships of the Piciformes (class Aves). Auk, 98, 481–494.
Steininger, F. (1999). The Continental European Miocene. Chronostratigraphy, geochronology and biochronology of the “miocene european land mammal mega-zones” (ELMMZ) and the miocene “mammal-zones” (MN-Zones). In G. E. Rössner & K. Heissig (Eds.), The miocene land mammals of Europe (pp. 9–24). München: Verlag Dr. Friedrich Pfeil.
Swierczewski, E. V., & Raikow, R. J. (1981). Hind limb morphology, phylogeny, and classification of the Piciformes. Auk, 98(3), 466–480.
Umanskaja, A. S. (1981). Miocenovye pticy Zapadnogo Pričernomor’ja USSR. Soobščenie II. [Miocene birds of western Pričernomor’e. Communication II.] Vestnik Zoologii, 17, 17–21. (In Russian).
Webb, D. M., & Moore, W. S. (2005). A phylogenetic analysis of woodpeckers and their allies using 12S, Cyt b, and COI nucleotide sequences (class Aves; order Piciformes). Molecular Phylogenetics and Evolution, 36, 233–248.
Wetmore, A. (1931). Record of an unknown woodpecker from the lower Pliocene. The Condor, 33(6), 255–256.
Winkler, H., & Christie, D. A. (2002). Family Picidae (Woodpeckers). In J. del Hoyo, A. Elliott, & J. Sargatal (Eds.), Handbook of the birds of the world. Vol 7. Jacamars to Woodpeckers. (pp. 296–555). Barcelona: Lynx Edicions.
Ziegler, R. (1999). Order Insectivora. In G. E. Rössner & K. Heissig (Eds.), The Miocene land mammals of Europe (pp. 53–74). München: Verlag Dr. Friedrich Pfeil.
Acknowledgments
We thank A. Louchart (École Normale Supérieure de Lyon, Lyon, France) and C. Mourer-Chauviré (Université Claude Bernard, Lyon, France) for their thorough comments which improved this manuscript. We are grateful to James Dean and Christopher Milensky (Smithsonian Institution, National Museum of Natural History, Division of Birds, for providing us with photographic material of Sasia abnormis. We also thank Sven Tränkner (Senckenberg Forschungsinstitut Frankfurt) for taking photographs d, i, n, q, and r of Fig. 2. We express our gratitude to Christian Meyer (Natural History Museum Basel) for allowing VLD to work on the fossil collection, and Marcel Güntert (Natural History Museum Bern) for providing us with useful information on extant Piciformes. AM is supported by the German Science Foundation (DFG MA 4809/1-1).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
De Pietri, V.L., Manegold, A., Costeur, L. et al. A new species of woodpecker (Aves; Picidae) from the early Miocene of Saulcet (Allier, France). Swiss J Palaeontol 130, 307–314 (2011). https://doi.org/10.1007/s13358-011-0021-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-011-0021-8