
Abstract
Local species populations that are more numerically abundant and occupy a greater proportion of the landscape relative to other species often recover more quickly (i.e., are more resilient) following local-scale environmental perturbations. In a companion study, we found that seatrout juvenile populations were distributed more broadly across Tampa Bay, Florida, and numerical abundance was generally much higher than those of similar-sized red drum, suggesting that spotted seatrout may also be generally more resilient to population declines than red drum. Following major population declines over a 12-year period (1996–2008), we found that larger juveniles of seatrout (51–100 mm standard length) simultaneously gained numerical abundance and broadened their spatial distribution generally within the next year. Population recovery in the same size of juvenile red drum generally took multiple years, and distribution and abundance increases were not always concurrent during the recovery period. Despite their overall higher abundance and broader spatial distribution, the smaller-sized juveniles of spotted seatrout (15–50 mm standard length (SL)) did not always recover more quickly from population declines compared with similar-sized red drum. Populations of the smaller-sized juveniles of both species often took multiple years to recover and showed non-concurrent increases in distribution and abundance during recovery. Despite their relatively narrow spatial distribution, juveniles of red drum may increase their overall resilience to local environmental perturbations by occupying multiple isolated patches across the Tampa Bay landscape. Monitoring dynamics of numerical abundance and spatial distribution may be helpful in gauging relative population resilience to facilitate overall management of these fishery populations.
Similar content being viewed by others
Introduction
Resilience is the ability of a population to recover after environmental perturbations (Holling 1973). Both low abundance and a relatively small spatial distribution can independently decrease the resilience of local species populations (Gaston and Curnutt 1998). Low population abundance increases the likelihood that demographic and environmental variability will eliminate populations. A local population with a restricted spatial distribution may be more vulnerable to environmental perturbations resulting in major declines or extinction (Lawton 1999; Gaston et al. 2000; Gunderson 2000; Purvis et al. 2000; Roberts et al. 2002; Williams et al. 2008). In a companion paper, Whaley et al. (2015, this issue) found that two size classes of spotted seatrout (15–50 mm and 51–100 mm) were distributed more broadly and abundance was generally much higher than those of similar-sized red drum. The more restricted distribution of red drum coupled with their lower abundance suggests they may be more vulnerable to environmental perturbations than spotted seatrout. Examining temporal dynamics of the spatial distribution-abundance relationship relative to environmental perturbations can help to identify relative resilience of juveniles of fishery species which may be useful for conservation planning (see review by Fisher et al. 2010).
We examined the dynamics of spatial distribution and population abundance of juveniles of two economically important fishery species (spotted seatrout, Cynoscion nebulosus and red drum, Sciaenops ocellatus) to examine population recovery to several major environmental perturbations that occurred in Tampa Bay, Florida during a 12-year period (1996–2008). This paper focuses on the influence of a major acid spill, a drought, and a major red tide event on the spatial distribution and abundance of juvenile populations in Tampa Bay. General spatial distribution and abundance patterns and the influence of freshwater inflow on these species populations are examined in our companion paper (Whaley et al. 2015).
Materials and Methods
Study Area
Tampa Bay (Fig. 1), located on Florida’s peninsular Gulf Coast, is surrounded by a highly urbanized watershed. Man-made and natural environmental perturbations (e.g., chemical spills, harmful algal blooms) periodically affect portions of the bay, temporarily reducing the quality of important estuarine nursery habitats for economically and culturally important fishery species. Despite these periodic events, Tampa Bay remains a popular recreational boating and fishing destination for visitors and the local population (Greene et al. 1997).

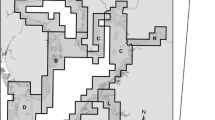
Map of eight sampling zones (shaded area) covering Tampa Bay (A–E), Florida, USA, and three major rivers (K–M) feeding into the bay
Data Collection
Juvenile fish data were collected monthly from 1997 to 2008 by the State of Florida’s Fisheries-Independent Monitoring Program (FIM; Florida Fish and Wildlife Conservation Commission, Fish and Wildlife Research Institute). Juveniles (generally 15–100 mm standard length) of red drum (Sciaenops ocellatus) and spotted seatrout (Cynoscion nebulosus) were collected with a 21.3-m center-bag seine (3.2-mm stretched mesh) targeting in shallow waters (depth ≤1.5 m) of the lower portions of three major rivers (Alafia, Little Manatee, and Manatee Rivers) and Tampa Bay proper (depth ≤1.8 m). Details of the field sample collection are described in our companion paper (Whaley et al. 2015) and so are not repeated here.
Data Analysis
By analyzing temporal abundance of juveniles, we defined peak recruitment season as June–September for smaller juveniles of spotted seatrout (15–50 mm), and August–November for larger juveniles (51–100 mm). Peak season of juvenile red drum abundance was defined as October–December for smaller juveniles (15–50 mm) and January–March for larger juveniles (51–100 mm). These months were combined to calculate an annual relative abundance for each zone and bay-wide. Each month, grid cells in each of eight zones were randomly selected for sampling (Fig. 1). To describe spatial distribution, we calculated the proportion of sampled grid cells with a positive species occurrence during the months of peak season of recruitment for each year of the study. Details regarding the estimation of annual abundance for each species and size class are described in our companion paper (Whaley et al. 2015).
We plotted the annual abundance and spatial distribution of each species (spotted seatrout and red drum) and size range (15–50 mm standard length (SL) and 51–100 mm SL) to examine year-to-year temporal trends in abundance, distribution, and abundance-distribution relationships bay-wide and among zones, particularly before and after several environmental perturbations (a major acid spill, a drought, and a major red tide) that occurred during the study period.
Results
Occurrence of Major Environmental Perturbations
The study period began in 1996, a period of moderate rainfall. During December of 1997, a heavy rainfall event caused a major acid spill in the Alafia River resulting in the instantaneous death of fish, crabs, and shrimp along 68 km of the river as well as a die-off of 136 ha of freshwater marshes. The spill also caused an extremely large influx of nitrogen and phosphorus into Tampa Bay, lasting for several months (Environmental Protection Commission of Hillsborough County 2000). Higher freshwater inflow persisted through 1998, followed by a prolonged drought associated with La Niña from 1999 to 2001. El Niño conditions again prevailed in 2002–2004 causing another period of high flow. Beginning in January 2005, a large red tide plume drifted into Tampa Bay and persisted into January 2006 causing a mass kill-off of estuarine nekton (Hu et al. 2006). This red tide event was followed by another prolonged drought beginning in 2006 and continued through to the end of the study period in 2008.
The 1997 Alafia Acid Spill and Juvenile Abundance and Distribution
In our companion study, we found the majority of the juvenile red drum population (both size classes) to occur within the major rivers of the Tampa Bay study area (Hillsborough, Alafia, Little Manatee, and Manatee Rivers), and were particularly concentrated in the Alafia River during the majority of the study (Whaley et al. 2015). Despite the degradation of this important nursery area caused by the Alafia acid spill, the overall abundance of both sizes of red drum across the Tampa Bay study area did not decline. Only a few months after the spill, 19 % of the larger juveniles and 14 % of the smaller juveniles of red drum occurred in the Alafia River. The distribution of the smaller-sized red drum population actually expanded broadly throughout several bay zones in 1998 with the largest proportions occurring in Old Tampa Bay (31 %) and in the Manatee River (zone M, 21 %, Fig. 2). The Manatee River also contained the largest proportion of larger red drum (45 %). In 1999, Hillsborough Bay (zone C), located directly downstream from the Alafia River, contained 59 % of the larger red drum. This zone contained less than 10 % of the population of larger red drum during most years of the study.

Yearly trends in spatial distribution and population abundance of juveniles of spotted seatrout and red drum over the study period (1997–2008). The ranges of axes are unique for each species and size class. Distribution and abundance of smaller-sized (15–50 mm SL) spotted seatrout changed in opposite directions during and following the drought of 2000 (a), as well as after the 2005 red tide event while distribution and abundance of larger (51–100 mm SL) spotted seatrout (b) consistently changed in the same direction throughout the study. Distribution and abundance of smaller (15–50 mm SL) (c), and larger (51–100 mm SL) (d) red drum changed in opposite directions directly after the Alafia acid spill, during the 2000 drought and during the drought following the red tide, a period of very low population abundance and distribution
Compared with similar-sized red drum, smaller- and larger-sized juveniles of spotted seatrout had higher overall population abundance and were more broadly distributed overall (Fig. 2), with the majority of the seatrout population located within the estuary proper (Whaley et al. 2015). The smaller-sized spotted seatrout (15–50 mm SL) reached their maximum abundance and spatial extent (Fig. 2) just after the Alafia acid spill. During this period, the spatial distribution of the smaller seatrout population occurred mainly in Old Tampa Bay, Hillsborough Bay, and Lower East portions of the bay (zones A, C, and E) which was not uncommon. Only 8 months after the Alafia acid spill, however, 43 % of the larger juveniles were located adjacent to the Alafia River in Hillsborough Bay (zone C), a similar and equally unusual pattern to that seen in the larger red drum in 1999. Across all other years, zone C averaged 10 % of the larger seatrout juvenile population.
The 2000 Drought and Juvenile Abundance and Distribution
Freshwater inflow was positively related to the abundance of red drum (both sizes) and the smaller-sized seatrout in our companion study (Whaley et al. 2015). Consequently, the drought (generally from 1999 to 2001) was a period of relatively low juvenile distribution and abundance. As populations recovered, increases in distribution and abundance were often asynchronous. For the smaller-sized juveniles of both red drum and seatrout, the first sign of recovery occurred with initial increases in abundance, yet their distribution was reduced in 2001. As inflow continued to increase, populations of smaller juveniles of both species continued to recover, increasing in distribution by 2003. In contrast to the smaller juveniles, larger red drum expanded their distribution, first in 2001, with considerable increases in abundance 2 years later. Although population levels of the larger-sized seatrout were not significantly related to freshwater inflow overall in our companion study, their abundance and distribution were also relatively low during the drought. Unlike red drum and the smaller seatrout, distribution and abundance of these larger seatrout changed in the same direction from year to year.
The 2005 Major Red Tide and Juvenile Abundance and Distribution
During the study period, the distribution and abundance of red drum juveniles (both sizes) declined most severely during the red tide of 2005. In the years following the red tide, the Tampa Bay region entered into another drought period and juveniles of red drum (both sizes) showed little signs of recovery. Distribution and abundance of juveniles of red drum fluctuated only slightly, often changing in opposite directions through the end of the study period. The abundance of the smaller red drum declined to levels around five times lower than prior to the red tide event and remained low with only minor increases through 2008. More dramatic declines occurred in the larger red drum population, as abundance remained an order of magnitude lower than that prior to the red tide. Both sizes of red drum exhibited the most restricted distribution in 2008, 3 years after the red tide (11 and 2 % of grid cells, respectively). Although the majority of both sizes of red drum were typically found in the rivers during most years, an unusual spatial pattern emerged in 2007–2008. During this period of extremely reduced population abundance, 64 % of the population of smaller red drum occurred in Old Tampa Bay (zone A). Equally unusual, the greatest proportion of the larger-sized red drum population occurred in Hillsborough Bay (zone C) during 2007 and 2008.
Similar to red drum, the red tide caused dramatic declines in the distribution and abundance of both sizes of spotted seatrout. Distribution of smaller seatrout declined from occurrence in over half of grid cells (54 %) in 2004 to only in 34–39 % during 2005–2007. The abundance of these smaller seatrout declined to levels around five times lower than prior to the red tide event and remained low until 2008, when abundance and distribution increased considerably. The larger juveniles of seatrout exhibited the most restricted distribution in the study during the major red tide event of 2005, when they occurred in only 14 % of grid cells, only half their maximum distribution seen in 1996. However, populations of the larger seatrout appeared to recover quickly with considerable gains in distribution and abundance in 2006, but declined again in 2007, and remained low through the end of the study period. Unusually high proportions of both sizes of seatrout (33 % of smaller and 30 % of larger juveniles) occurred in Old Tampa Bay (zone A) during the 2005 red tide, and this pattern continued for 3 years to the end of the study period.
Discussion
Abundance-Distribution Relationships Following Environmental Perturbations
We found significant abundance-distribution relationships for both size classes of spotted seatrout and red drum in our companion study, a relationship typically found in many taxa (Brown 1984; Gaston et al. 2000; Foggo et al. 2003). However, a detailed examination of annual changes revealed that distribution and abundance of both sizes of red drum and the smaller spotted seatrout did not always change in the same direction from year to year. These discordant changes occurred during multi-year recovery periods following dramatic declines in population abundance and/or distribution. In contrast to red drum and smaller juveniles of seatrout, the larger juveniles of spotted seatrout recovered relatively quickly following population declines (generally within the next year), and distribution and abundance of these larger seatrout consistently changed in the same direction from year to year. Dynamic changes in distribution and abundance of fishes have rarely been examined (Fisher et al. 2010). However, Fisher and Frank (2004) also found positive abundance-distribution relationships in half the fishery stocks they examined off the Scotian Shelf and Bay of Fundy, Canada, with notable exceptions following ecological perturbations (e.g., over fishing). More research is needed to determine if distribution and abundance changing in opposite directions, as we saw in this study, might be an indicator of population recovery from environmental perturbations. This would be helpful in situations where perturbations are not well known, unlike the acid spill, drought, and major red tide in this study.
Relationships Between Spatial Distribution and Species Resilience
We found that during the majority of the study, juvenile red drum were generally less abundant and occupied a smaller proportion of the study area compared with similar-sized spotted seatrout. This suggests that juvenile red drum may be generally more vulnerable to environmental perturbations compared to spotted seatrout. We found some evidence to support this as the larger juvenile seatrout appeared to recover quickly (usually within the next year) from major population declines throughout the study, yet larger juveniles of red drum took multiple years to recover from the same major disturbances. In contrast, relative recovery time for the smaller-sized juveniles of both species was not consistent and depended on the nature of the perturbation. Smaller-sized spotted seatrout, with their larger populations and broader overall distribution, appeared to begin recovery sooner than similar-sized red drum after the red tide event. However, smaller red drum showed signs of recovery from the drought of 2000 sooner than seatrout.
If resilience of a local population increases with spatial distribution and abundance, then the dynamics of juvenile distribution and abundance suggests that resilience of these populations may also be dynamic. For those juveniles we found to have positive inflow-distribution and inflow-abundance relationships (in our companion study), resilience to environmental perturbations may also vary positively with freshwater inflow. Alternatively, an event, such as a red tide or a chemical spill that causes a decline in population abundance and/or spatial distribution, may reduce the population’s resilience to other stressors, including droughts. Although resilience can be difficult to quantify, the population responses of juveniles of red drum and spotted seatrout after the Alafia acid spill in 1997, drought of 2000, and the major red tide event in 2005 provide some insight into the potential influence of freshwater inflow on population resilience. In the high inflow period following the 1997 acid spill, red drum juveniles appeared to rebound relatively quickly despite drastic habitat degradation of the Alafia River, an important nursery area. The most drastic effects of the spill were within the river, which was not an area of high concentration for juveniles of spotted seatrout. In addition, seatrout had a much broader distribution across the study area than red drum. Both species appeared to be resilient to the effects of the acid spill during this period of high freshwater inflow. Hillsborough Bay (zone C), directly downstream from the Alafia acid spill, contained elevated levels of nutrients in 1998 after this spill (Johansson 2002; Environmental Protection Commission of Hillsborough County 2000). Elevated nutrient levels may have caused the unusually large increases in abundance of larger juveniles of spotted seatrout (in 1998) and red drum (in 1999) in this zone. Yet, water clarity was also reduced which may have reduced predation on these juveniles. In contrast, a drought period following the major red tide event of 2005 was coincident with a sustained reduction in distribution and abundance of red drum juveniles (both sizes) and smaller spotted seatrout over several years. However, a sustained decline in spawning adults caused by the red tide fish kill may have been the main cause of the continued decline of juveniles, and this may have been exacerbated by low freshwater inflow conditions. These events, though anecdotal, suggest that for those populations influenced by freshwater (smaller juveniles of seatrout and both size classes of juvenile red drum), resilience (time to population recovery) may also be positively related to freshwater inflow. Therefore, occurrence of environmental perturbations during or followed by heavy inflow periods may lead to shorter recovery periods for these juvenile populations than those events occurring during or followed by prolonged drought conditions, particularly with regard to red drum.
Similar to the influences of relative abundance and spatial distribution, the number of occupied patches is likely positively related to the resilience and persistence of these juvenile populations in Tampa Bay. The number of isolated patches of species occupation has been positively related to the persistence of metapopulations of some species (Hanski et al. 1996; Bascompte et al. 2002). The distribution of a number of occupied patches of juveniles throughout Tampa Bay likely contributed to population resilience to the 1997 acid spill and 2005 red tide. Vulnerability of red drum to environmental perturbations affecting one river, such as the Alafia acid spill, was reduced because juvenile red drum were distributed over several individual rivers. A large proportion of the population occurred in the Manatee River in the years after the Alafia acid spill. Similarly, unusually large proportions of juvenile populations of spotted seatrout were found in Old Tampa Bay during and after the red tide event. Hydrography of the bay isolates this area from the rest of the bay with regard to circulation, and salinities can be typically lower than the requirements of the red tide organism (Flaherty and Landsberg 2011). Therefore, Old Tampa Bay appears to be a refuge for juveniles of spotted seatrout, and potentially for other estuarine animals as well, during red tide events occurring in Tampa Bay. Walters et al. (2013) found Old Tampa Bay to also be a refuge for spawning sand seatrout (Cynoscion arenarius) during the 2005 red tide. Generally, it appears that distributing juveniles over multiple areas within the estuarine/riverine landscape may improve resilience of these populations to local-scale perturbations that affect a proportion of the landscape. However, this may not improve resilience to regional-scale perturbations, such as droughts, that affect the entire local population.
Some research suggests that the use of density-based measures, such as catch-per-unit-effort, as an index of population abundance could be improved with information on spatial distribution (e.g., Brown 1984; Petitgas 1998; Gaston et al. 2000). Mean density of adult fish stocks may not decline, yet they may occupy a smaller area and thus have a lower overall population size. Monitoring both distribution and abundance of juveniles of fishery species may also provide more information on status of declining and recovering populations. Therefore, both abundance and spatial distribution of a species or a vulnerable life stage can provide unique information regarding its relative resilience to environmental perturbations. For example, an increase in distribution may occur first, as the larger juveniles of red drum did after the 1997 Alafia acid spill and the 2000 drought, and be an early indicator of population recovery prior to increasing in population abundance. Where sufficient long-term monitoring programs exist, similar examinations of intraspecific and interspecific abundance and distribution dynamics may prove to be key in gauging sources of environmental stress and relative resilience of estuarine-dependent fishery species (see Fisher et al. 2010). In addition, more analysis is needed to examine the influence of number of patches on population resilience and persistence of juveniles of fishery species. The number of occupied patches may affect population survival and productivity at other life stages as well, particularly for those fishery species that aggregate on specific and limited spawning grounds (Bascompte et al. 2002; Walters et al. 2013). When identifying important habitat areas for conservation, certain areas (e.g., Old Tampa Bay) may provide an “insurance policy” for juvenile populations when environmental perturbations, such as red tide and acid spills, reduce habitat quality in their typical area of occupation.
References
Bascompte, J., H. Possingham, and J. Roughgarden. 2002. Patchy populations in stochastic environments: critical number of patches for persistence. The American Naturalist 159(2): 128–137.
Brown, J.H. 1984. On the relationship between abundance and distribution of species. The American Naturalist 124(2): 255–279.
Environmental Protection Commission of Hillsborough County. 2000. Final damage assessment and restoration plan and environmental assessment for the December 7, 1997 Alafia River spill. http://www.gc.noaa.gov/gc-rp/muldarp2.pdf
Fisher, J.A.D., and K.T. Frank. 2004. Abundance–distribution relationships and conservation of exploited marine fishes. Marine Ecology Progress Series 279: 201–213.
Fisher, J.A.D., K.T. Frank, and W.C. Leggett. 2010. Dynamic macroecology on ecological time-scales. Global Ecology and Biogeography 19: 1–15.
Flaherty, K.E., and J.H. Landsberg. 2011. Effects of a persistent red tide (Karenia brevis) bloom on community structure and species-specific relative abundance of nekton in a Gulf of Mexico estuary. Estuaries and Coasts 34: 417–439.
Foggo, A., M.T. Frost, and M.J. Attrill. 2003. Abundance-occupancy patterns in British estuarine macroinvertebrates. Marine Ecology Progress Series 265: 297–302.
Gaston, K.J., and J.L. Curnutt. 1998. The dynamics of abundance-range size relationships. Oikos 81: 38–44.
Gaston, K.J., T.M. Blackburn, J.J.D. Greenwood, R.D. Gregory, R.M. Quinn, and J.H. Lawton. 2000. Abundance–occupancy relationships. Journal of Applied Ecology 37(Suppl. 1): 39–59.
Greene, G., C.B. Moss, and T.H. Spreen. 1997. Demand for recreational fishing in Tampa Bay, Florida: A random utility approach. Marine Resource Economics 293–305.
Gunderson, L.H. 2000. Ecological resilience—in theory and application. Annual Review of Ecology and Systematics 31: 425–439.
Hanski, I., A. Moilanen, and M. Gyllenberg. 1996. Minimum viable metapopulation size. American Naturalist 147: 527–541.
Holling, C. 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4: 1–23.
Hu, C., F.E. Muller-Karger, and P.W. Swarzenski. 2006. Hurricanes, submarine groundwater discharge, and Florida’s red tides. Geophysical Research Letters 33(L11601): 5.
Johansson, J.O. R. 2002. Historical overview of Tampa Bay water quality and seagrass issues and trends. Reports. Paper 147. http://scholarcommons.usf.edu/basgp_report/147
Lawton, J.H. 1999. Are there general laws in ecology? Oikos 84: 177–192.
Petitgas, P. 1998. Biomass-dependent dynamics of fish spatial distributions characterized by geostatistical aggregation curves. ICES Journal of Marine Science 55: 443–453.
Purvis, A., J.L. Gittleman, G. Cowlishaw, and G.M. Mace. 2000. Predicting extinction risk in declining species. Proceedings of the royal society of London. Series B: Biological Sciences 267(1456): 1947–1952.
Roberts, C.M., C.J. McClean, J.E. Veron, J.P. Hawkins, G.R. Allen, D.E. McAllister, C.G. Mittermeier, et al. 2002. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295(5558): 1280–1284.
Walters, S., S. Lowerre-Barbieri, J. Bickford, J. Tustison, and J.H. Landsberg. 2013. Effects of Karenia brevis red tide on the spatial distribution of spawning aggregations of sand seatrout Cynoscion arenarius in Tampa Bay, Florida. Marine Ecology Progress Series 479: 191–202.
Whaley, S.D., M.C. Christman, J.J. Burd Jr. 2015. Spatial distribution-abundance relationships in juvenile (Age-0) red drum (Sciaenops ocellatus) and spotted seatrout (Cynoscion nebulosus). I: Influence of Freshwater Inflow. (This issue).
Williams, S.E., L.P. Shoo, J.L. Isaac, A.A. Hoffmann, and G. Langham. 2008. Towards an integrated framework for assessing the vulnerability of species to climate change. PLoS Biology 6(12): e325.
Acknowledgments
We are grateful to the Florida Fish and Wildlife Research Institute’s Fisheries-Independent Monitoring program for collecting the field data used in this analysis. This sampling program is partially supported by the US Fish and Wildlife Service (Sport Fish Restoration Grant F-43). In addition, we greatly appreciate the support by senior staff (H. Norris, R. Flamm, and K. O’Keife) and the entire Center for Spatial Analysis team at FWRI (partially supported by the US Fish and Wildlife Service, Sport Fish Restoration Grant F-66). This manuscript benefited from valuable comments from W. Cooper, M. Barrett, R. Butryn, C. Harmak, and several anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Josianne G. Støttrup
Rights and permissions
About this article

Cite this article
Whaley, S.D., Christman, M.C. & Burd, J.J. Spatial Distribution-Abundance Relationships in Juvenile (Age-0) Red Drum (Sciaenops Ocellatus) and Spotted Seatrout (Cynoscion Nebulosus). II: Influence of Major Disturbances. Estuaries and Coasts 39, 752–758 (2016). https://doi.org/10.1007/s12237-015-0035-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-015-0035-3