
Abstract
The history of biological systematics documents a continuing tension between classifications in terms of nested hierarchies congruent with branching diagrams (the ‘Tree of Life’) versus reticulated relations. The recognition of conflicting character distribution led to the dissolution of the scala naturae into reticulated systems, which were then transformed into phylogenetic trees by the addition of a vertical axis. The cladistic revolution in systematics resulted in a representation of phylogeny as a strictly bifurcating pattern (cladogram). Due to the ubiquity of character conflict—at the genetic or morphological level, or at any level in between—some characters will necessarily have to be discarded (qua noise) in favor of others in support of a strictly bifurcating phylogenetic tree. Pattern analysts will seek maximal congruence in the distribution of characters (ultimately of any kind) relative to a branching tree-topology; process explainers will call such tree-topologies into question by reference to incompatible evolutionary processes. Pattern analysts will argue that process explanations must not be brought to bear on pattern reconstruction; process explainers will insist that the reconstructed pattern requires a process explanation to become scientifically relevant, i.e., relevant to evolutionary theory. The core question driving the current debate about the adequacy of the ‘Tree of Life’ metaphor seems to be whether the systematic dichotomization of the living world is an adequate representation of the complex evolutionary history of global biodiversity. In ‘Questioning the Tree of Life’, it seems beneficial to draw at least four conceptual distinctions: pattern reconstruction versus process explanation as different epistemological approaches to the study of phylogeny; open versus closed systems as expressions of different kinds of population (species) structures; phylogenetic trees versus cladograms as representations of evolutionary processes versus patterns of relationships; and genes versus species as expressions of different levels of causal integration and evolutionary transformation.
Similar content being viewed by others


Notes
Notebook B, p. 36, Cambridge University Library, #DAR121.
Bonnet was introduced to Locke’s philosophy by his teacher, the Genevan mathematician and philosopher Gabriel Cramer (Savioz 1948).
Patterson’s talk, delivered at the 2nd Annual Willi Hennig Society Meeting on October 3, 1981, in Ann Arbor, MI, was transcribed and made available by D.M. Williams, Dept. of Botany, The Natural History Museum, London.
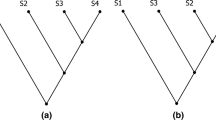
The test was based on the assumption that characters used to infer species relationships are unique (no convergence occurs) and un-reversed (character transformation is irreversible)—two desiderata that are both unwarranted.
For the ‘Questioning the Tree of Life’ initiative, see http://centres.exeter.ac.uk/eugenis/research/QuestioningtheTreeofLife.htm.
References
Abel O (1929) Paläobiologie und Stammesgeschichte. G. Fischer, Jena
Agassiz L (1859) An essay on classification. Longman, Brown, Green, Longmans and Roberts, London
Archibald JM, Rogers MB, Toop M, Ishida K, Keeling PJ (2003) Lateral gene transfer and the evolution of plastid-targeted proteins in the secondary plastid-containing alga Bigelowiella natans. PNAS 100:7678–7683
Armstrong DM (1997) A world of states of affairs. Cambridge University Press, Cambridge
Baer KE (1828) Ueber Entwickelungsgeschichte der Thiere. Beobachtung und Reflexion, Theil I. Gebr. Bornträger, Königsberg
Bapteste E, Boucher Y (2008) Lateral gene transfer challenges principles of microbial systematics. Trends Microbiol 16:200–207
Bapteste E, Susko E, Leigh J, MacLeod D, Charlebois RL, Doolittle WF (2005) Do orthologous gene phylogenies really support tree-thinking? BMC Evol Biol 5:33. doi:10.1186/1471-2148-5-33
Barsanti G (1992) La scala, la mappa, l’albero. Imagini e classificazioni della natura fra Sei e Ottocento. Sansoni, Firenze
Beatty J (1982) Classes and cladists. Syst Zool 31:25–34
Beatty J (1995) The evolutionary contingency thesis. In: Wolters G, Lennox JG (eds) Concepts, theories, and rationality in the biological sciences. University of Pittsburgh Press, Pittsburgh, pp 45–81
Bergthorsson U, Richardson AO, Young GLJ, Goertzen LR, Palmer JD (2004) Massive horizontal transfer of mitochondrial genes from diverse land plant donors to the basal angiosperm Amborella. PNAS 101:17747–17752
Bloch K (1956) Zur Theorie der naturwissenschaftlichen Systematik, unter besonderer Berücksichtigung der Biologie. Brill, Leiden
Bonde N (1974) [Review of] Interrelationships of fishes, Greenwood, P.H., R.S. Miles, C. Patterson (eds.). Syst Zool 23:562–569
Bonnet Ch (1745) Traîté d’Insectologie. Première Partie. Durand Librairie, Paris
Bonnet Ch (1764) Contemplation de la Nature, vol 1. Marc-Michel Rey, Amsterdam
Bonnet Ch (1768) Considérations sur les Corps Organisés, vol 1, 2nd edn. Marc-Michel Rey, Amsterdam
Bonnet Ch (1769) La Palingénésie Philosophique, vol 1. C. Philibert and B. Chirol, Geneva
Boucher Y, Bapteste E (2009) Revisiting the concept of lineage in prokaryotes: a phylogenetic perspective. Bioessays 31:526–536
Bowler PJ (1990) Charles Darwin. The man and his influence. Basil Blackwell, Oxford
Bowler C, Allen AE, Badger JH, Grimwood J, Jabbari K, Kue A, Maheswari U, Martens C, Maumus F, Otillar RP, Rayko E, Salamov A, Vandepoele K, Beszteri B, Gruber A, Heijde M, Katinka M, Mock T, Valentin K, Verret F, Berges JA, Brownlee C, Cadoret JP, Chiovitti A, Choi CJ, Coesel S, De Martino A, Detter JC, Durkin C, Falciatore A, Fournet J, Haruta M, Huysman MJJ, Jenkins BD, Jiroutova K, Jorgensen RE, Joubert Y, Kaplan A, Kröger N, Kroth PG, La Roche J, Lindquist E, Lommer M, Martin-Jézéquel V, Lopez PJ, Lucas S, Mangogna M, McGinnis K, Medlin LK, Montsant A, Oudot-Le Secq MP, Napoli C, Obornik M, Schnitzler Parker M, Petit JLL, Porcel BM, Poulsen N, Robinson M, Rychlewski L, Rynearson TA, Schmutz J, Shapiro H, Siaut M, Stanley M, Sussman MR, Taylor AR, Vardi A, von Dassow P, Vyverman W, Willis A, Wyrwicz LS, Rokhsar DS, Weissenbach J, Armbrust EV, Green BR, Van de Peer Y, Grigoriev IVV (2008) The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 456:239–244
Boyd R (1999) Homeostasis, species, and higher taxa. In: Wilson RA (ed) Species. New interdisciplinary essays. MIT Press, Cambridge, pp 141–185
Brady RH (1985) On the independence of systematics. Cladistics 1:113–126
Bredekamp H (2005) Darwins Korallen. Wagenbach Verlag, Berlin
Brigandt I (2009) Natural kinds in evolution and systematics. Acta Biotheor 57:77–97
Brooks DR, McLennan DA (1991) Phylogeny, ecology, and behavior. University of Chicago Press, Chicago
Brower AVZ (2000) Evolution is not a necessary assumption of cladistics. Cladistics 16:143–154
Brower AVZ, DeSalle R, Vogler A (1996) Gene trees, species trees, and systematics. Annu Rev Ecol Syst 27:423–450
Brundin L (1966) Transantarctic relationships and their significance, as evidenced by chironomid midges. K svenska VetenskAkad Handl 4(11):1–472
Buffon GLL (1749) Histoire Naturelle, Générale et Particulière, vol 1. Imprimerie Royale, Paris
Chambers R (1844) Vestiges of the natural history of creation. John Churchill, London
Craw R (1992) Margins of cladistics: identity, difference and place in the emergence of phylogenetic systematics, 1864–1975. In: Griffiths P (ed) Trees of life: essays in philosophy of biology. Kluwer, Dordrecht, pp 65–107
Cuvier G (1817) Le Règne Animal distribué d’après son Organisation, vol 1. Deterville, Paris
Daudin H (1926a) Cuvier et Lamarck. Les Classes Zoologiques et l’Idée de la Série Animale (1790–1830). Félix Alcan, Paris
Daudin H (1926b) De Linné à Jussieu. Méthode de la Classification et l’Idée de la Série en Botanique et en Zoologie (1740–1790). Félix Alcan, Paris
Davis CC, Wurdack KJ (2004) Host-to-parasite gene transfer in flowering plants: phylogenetic evidence from Malpighiales. Science 305:676–678
deBeer G (1960) Darwin’s notebooks on transmutation of species. Bull Br Mus Nat Hist (Hist Ser) 2:23–73
Denker E, Bapteste E, Le Guyader H, Manuel M, Rabet N (2008) Horizontal gene transfer and the evolution of the cnidarian stinging cells. Curr Biol 18(18):R858–R859
deQueiroz K (1999) The general lineage concept of species and the defining properties of the species category. In: Wilson RA (ed) Species. New interdisciplinary essays. MIT Press, Cambridge, pp 49–89
Dollo L (1895) Sur la phylogénie des dipneustes. Bull Soc Belge Géol Paléont Hydrol 9:97–128
Doolittle WF (1999) Phylogenetic classification and the universal tree. Science 284:2124–2128
Doolittle WF (2009) The practice of classification and the theory of evolution, and what the demise of Charles Darwin’s tree of life hypothesis means for both of them. Philos Trans R Soc Lond B364:2221–2228
Doolittle WF, Bapteste E (2007) Pattern pluralism and the Tree of Life Hypothesis. PNAS 104:2043–2049
Duméril AMC (1806) Zoologie Analytique, ou Méthode Naturelle de Classification, 8 vols. Allais Librairie, Paris
Dunning Hotopp JC, Clark ME, Oliveira DCSG, Foster JM, Fischer P, Munoz Torres MC, Giebel JD, Kumar N, Ishmael N, Wang S, Ingram J, Nene RV, Shepard J, Tomkins J, Richards S, Spiro DJ, Ghedin E, Slatko BE, Tettelin H, Werren JH (2007) Widespread lateral gene transfer from intracellular bacteria to multicellular eukaryotes. Science 317:1753–1756
Dupuis C (1984) Willi Hennig’s impact on taxonomic thought. Annu Rev Ecol Syst 15:1–24
Ereshefsky M (2010) Microbiology and the species problem. Biol Philos (this issue—details to follow)
Funk V (1985) Phylogenetics, patterns, and hybridization. Ann Missouri Bot Gard 72:596–635
Gaffney ES (1979) An introduction to the logic of phylogeny reconstruction. In: Cracraft J, Eldredge N (eds) Phylogenetic analysis and paleontology. Columbia University Press, New York, pp 79–111
Gardner S (1999) Kant and the critique of pure reason. Routledge, London
Ghiselin M (1974) A radical solution to the species problem. Syst Zool 23:536–544
Ghiselin M (1997) Metaphysics and the Origin of Species. SUNY Press, Albany
Gladyshev EA, Meselson M, Arkhipova IR (2008) Massive horizontal gene transfer in bdelloid rotifers. Science 320:1210–1213
Gogarten P (2000) Horizontal gene transfer: a new paradigm for biology. Esalen Center for Theory and Research Conference. http://www.esalenctr.org/display/confpage.cfm?confid=10&pageid=105&pgtype=1. Retrieved on 03-24-2009
Gogarten P, Townsend JP (2005) Horizontal gene transfer, genome innovation and evolution. Nat Rev Microbiol 3:679–687
Gogarten P, Doolittle WF, Lawrence JG (2002) Prokaryotic evolution in light of gene transfer. Mol Biol Evol 19:2226–2238
Gould SJ (1977) Ontogeny and phylogeny. Harvard University Press, Cambridge
Grant PR, Grant BR (2002) Unpredictable evolution in a 30-year study of Darwin’s finches. Science 296:707–711
Grant PR, Grant BR, Markert JA, Keller LF, Petren K (2004) Convergent evolution of Darwin’s finches caused by introgressive hybridization and selection. Evolution 58:1588–1599
Greenwood PH, Miles RS, Patterson C (1973) Interrelationships of fishes. Academic Press, London
Griffiths PE (1999) Squaring the circle: natural kinds with historical essences. In: Wilson RA (ed) Species. New interdisciplinary essays. MIT Press, Cambridge, pp 209–228
Günther K (1956) Systemlehre und Stammesgeschichte. Fortschr Zool NF 10:33–278
Günther K (1962) Systematik und Stammengeschichte der Thiere, 1954–1959. Fortschr Zool NF 14:268–547
Hennig W (1950) Grundzüge einer Theorie der Phylogenetischen Systematik. Deutscher Zentralverlag, Berlin
Hennig W (1957) Systematik und Phylogenese. In: Hannemann HJ (ed) Bericht über die Hundertjahrfeier der Deutschen Entomologischen Gesellschaft Berlin. Akademie Verlag, Berlin, pp 50–71
Hennig W (1965) Phylogenetic systematics. Annu Rev Entomol 10:97–116
Hennig W (1966) Phylogenetic systematics. University of Illinois Press, Urbana
Hull DL (1976) Are species really individuals. Syst Zool 25:174–191
Hull DL (1979) The limits of cladism. Syst Zool 28:416–440
Hull DL (1988) Science as a process. An evolutionary account of the social and conceptual development of science. The University of Chicago Press, Chicago
Hull DL (1989) The metaphysics of evolution. State University of New York Press, Albany
Hull DL (1999) On the plurality of species: questioning the party line. In: Wilson RA (ed) Species. New interdisciplinary essays. MIT Press, Cambridge, pp 23–48
Humphries CJ (1983) Primary data in hybrid analysis. In: Platnick NI, Funk VA (eds) Advances in cladistics, vol 2. Columbia University Press, New York, pp 89–103
Kluge AG (2003) The repugnant and the mature in phylogenetic inference: atemporal similarity and historical identity. Cladistics 19:356–368
Konstantinidis KT, Ramette A, Tiedje JM (2006) The bacterial species definition in the genomic era. Philos Trans R Soc Lond B361:1929–1940
Kuhn TS (1974) Logic of discovery or psychology of research. In: Lakatos I, Musgrave A (eds) Criticism and the growth of knowledge. Cambridge University Press, Cambridge, pp 1–23
Lauder GV (1990) Functional morphology and systematics: studying functional patterns in an historical context. Annu Rev Ecol Syst 21:317–340
Laurin M, Bryant HN (2009) The third meeting on the International Society for Phylogenetic Nomenclature: a report. Zool Scr 38:333–337
Lienau EK, DeSalle R (2009) Evidence, content and corroboration and the tree of life. Acta Biotheor 57:187–199
Loftus BL, Anderson I, Davies R, Alsmark UCM, Samuelson J, Amedeo P, Roncaglia P, Berriman M, Hirt RP, Mann BJ, Nozaki T, Suh B, Pop M, Duchene M, Ackers J, Tannich E, Leippe M, Hofer M, Bruchhaus I, Willhoeft U, Bhatttacharya A, Chillingworth T, Churcher C, Hance Z, Harris B, Harris D, Jagels K, Moule S, Mungall K, Ormond D, Squares R, Whitehead S, Quail MA, Rabbinowitsch E, Norbertczak H, Price C, Wang Z, Gullién N, Gilchrist C, Stroup SE, Bhattacharya S, Lohia A, Foster PG, Sicheritz-Ponten T, Weber C, Singh U, Mukherjee C, El-Sayed NM, Petri WA Jr, Clark CG, Embley TM, Barrell B, Fraser CM, Hall N (2005) The genome of the protist parasite Entamoeba histolytica. Nature 433:865–868
Lorenz K (1941a) Kants Lehre vom Apriorischen im Lichte gegenwärtiger Biologie. Blätter Deutsch Philos 15:94–125
Lorenz K (1941b) Vergleichende Bewegungsstudien an Anatinen. J Ornithol 89(Suppl):194–293
Lovejoy AO (1936) The great chain of being. Harvard University Press, Cambridge
Lovejoy AO (1959) Recent criticism of the Darwinian theory of recapitulation: its grounds and its initiator. In: Glass B, Temkin O, Straus WK Jr (eds) Forerunners of Darwin, 1745–1859. The Johns Hopkins University Press, Baltimore, pp 438–458
Maddison WP (1997) Gene trees in species trees. Syst Biol 46:523–536
Magee B (1973) Popper. Fontana, New York
Mayr E (1963) Animal species and evolution. Belknap Press at Harvard University Press, Cambridge
Mayr E (1969) Principles of systematic zoology. McGraw-Hill, New York
Mayr E (1974) Cladistic analysis or cladistic classification. Zeitschr Zool Syst Evolutforsch 12:94–128
Mayr E (1982) The growth of biological thought. The Belknap Press at Harvard University Press, Cambridge
Miles RS (1973) Relationships of acanthodians. In: Greenwood PH, Miles RS, Patterson C (eds) Interrelationships of fishes. Academic Press, London, pp 63–103
Miles RS (1975) The relationships of the Dipnoi. In: Lehman JP (ed) Problèmes Actuels de Paléontologie (Evolution des Verttébrés). Colloques Internationaux du Centre National de la Rercherche Scientifique, Paris, pp 133–148
Miller H (1849) Footprints of the Creator, or the Asterolepis of Stromness. Johnstone and Hunter, London
Millhauser M (1959) Just before Darwin. Wesleyan University Press, Middletown
Müller-Wille S (2007) Collection and collation: theory and practice of Linnean botany. Stud Hist Philos Biol Biomed Sci 38:541–562
Nedelcu AM, Miles IH, Fagir AM, Karol K (2008) Adaptive eukaryote-to-eukaryote lateral gene transfer: stress-related genes of algal origin in the closest unicellular relatives of animals. J Evol Biol 21:1852–1860
Nelson G (2004) Cladistics: its arrested development. In: Williams DM, Forey PL (eds) Milestones in systematics. CRC Press, Boca Raton, pp 127–147
Nelson G (2007) Patterson, Colin. In: Koertge N (ed) New dictionary of scientific biography, vol 6, 2nd edn. Gale Cengage, Farmington Hills, pp 30–34
Okasha S (2006) Evolution and the levels of selection. Clarendon Press, Oxford
Ospovat D (1981) The development of Darwin’s theory. Cambridge University Press, Cambridge
Panchen AL (1992) Classification, evolution, and the nature of biology. Cambridge University Press, Cambridge
Patterson C (1981) Significance of fossils in determining evolutionary relationships. Annu Rev Ecol Syst 12:195–223
Patterson C (1982) Morphological characters and homology. In: Joysey K, Friday AE (eds) Problems of phylogenetic reconstruction. Academic Press, London, pp 21–74
Patterson C (2002) Evolution and creationism. The Linnean 18:15–32
Platnick NI (1977) Cladograms, phylogenetic trees, and hypothesis testing. Syst Zool 26:438–442
Popper KR (1959 [1992]) The logic of scientific discovery. Routledge and Kegan Paul, London
Popper KR (1963) Conjectures and refutations: the growth of scientific knowledge. Routledge and Kegan Paul, London
Rensch B (1947) Neuere Probleme der Abstammungslehre: Die Transspezifische Evolution. Enke, Stuttgart
Rensch B (1968) Biophilosophie auf erkenntnistheoretischer Grundlage (Panpsychistischer Identismus). Fischer, Stuttgart
Rensch B (1979) Lebensweg eines Biologen in einem turbulenten Jahrhundert. Fischer, Stuttgart
Retchless AC, Lawrence JG (2007) Temporal fragmentation of species in bacteria. Science 317:1093–1096
Richards RJ (1992) The meaning of evolution. The morphological construction and ideological reconstruction of Darwin’s theory. University of Chicago Press, Chicago
Richards RJ (2002) The romantic conception of life. Science and philosophy in the age of Goethe. University of Chicago Press, Chicago
Richards RJ (2008) The tragic sense of life. Ernst Haeckel and the struggle over evolutionary thought. University of Chicago Press, Chicago
Rieppel O (1986) Species are individuals: a review and critique of the argument. Evol Biol 20:283–317
Rieppel O (2004) The language of systematics, and the philosophy of ‘total evidence’. Syst Biodivers 2:9–19
Rieppel O (2005) The philosophy of total evidence and its relevance for phylogenetic inference. Pap Avulsos Zool 45:77–89
Rieppel O (2006a) On concept formation in systematics. Cladistics 22:474–492
Rieppel O (2006b) Willi Hennig on transformation series: metaphysics and epistemology. Taxon 55:377–385
Rieppel O (2007a) Parsimony, likelihood, and instrumentalism in systematics. Biol Philos 22:141–144
Rieppel O (2007b) The metaphysics of Hennig’s phylogenetic systematics: substance, events and laws of nature. Syst Biodivers 5:345–360
Rieppel O (2007c) Species: kinds of individuals or individuals of a kind. Cladistics 23:373–384
Rieppel O (2008a) Total evidence in phylogenetic systematics. Biol Philos. doi:10.1007/s10539-008-9122-1
Rieppel O (2008b) Re-writing Popper’s philosophy of science for systematics. Hist Philos Life Sci 30:317–340
Rieppel O (2008c) Hypothetico-deductivism in systematics: fact or fiction? Pap Avulsos Zool 48:263–273
Rieppel O (2009) Species as a process. Acta Biotheor 57:33–49
Rieppel O, Rieppel M, Rieppel L (2006) Logic in systematics. J Zool Syst Evol Res 44:186–192
Russell ES (1916) Form and function. A contribution to the history of animal morphology. John Murray, London
Savioz R (1948) Mémoires autobiographiques de Charles Bonnet de Genève. Librairie Philosophique J. Vrin, Paris
Serres E (1824) Explication du système nerveux des animaux invertébrés. Ann Sci Nat Paris 3:377–380
Simpson GG (1961) Principles of animal taxonomy. Columbia University Press, New York
Stamos DN (2007) Popper, laws, and the exclusion of biology from genuine science. Acta Biotheor 55:357–375
Steiner H (1946) Sinai Tschulok (16. April 1875 bis 6. Dezember 1945). Vierteljahresschr Natforsch Ges Zürich 91:70–72
Stevens PF (1994) The development of biological systematics. Columbia University Pess, New York
Striepen B, Pruijssers AJP, Huang J, Li C, Gubbels MJ, Umejiego NN, Hedstrom L, Kissinger JC (2004) Gene transfer in the evolution of parasite nucleotide biosynthesis. PNAS 101:3154–3159
Takhtajan A (1959) Die Evolution der Angiospermen. Fischer, Jena
Trembley A (1744) Mémoires Pour Servir à l’Histoire d’un Genre de Polypes d’Eau Douce. Jean & Herman Verbeek, Leiden
Tschulok S (1910) Das System der Biologie in Forschung und Lehre. Fischer, Jena
Tschulok S (1922) Deszendenzlehre. Fischer, Jena
Wagner WH (1983) Reticulistics: the recognition of hybrids and their role in cladistics and classification. In: Platnick NI, Funk VA (eds) Advances in cladistics, vol 2. Columbia University Press, New York, pp 63–79
Wagner PJ, Erwin DH (1995) Phylogenetic patterns as tests of speciation models. In: Erwin DH, Anstey RL (eds) New approaches to speciation in the fossil record. Columbia University Press, New York, pp 87–122
Wheeler QD, Meier R (2000) Species concepts and phylogenetic theory. A debate. Columbia University Press, New York
Williams PA (1992) Confusion in cladism. Synthese 91:135–152
Williams DM, Ebach MC (2007) Foundations of systematics and biogeography. Springer, Berlin
Williams DM, Scotland RW, Humphries CJ, Siebert DJ (1996) Confusion in philosophy: a comment on Williams (1992). Synthese 108:127–136
Wilson EO (1965) A consistency test for phylogenies based on contemporaneous species. Syst Zool 14:214–220
Wilson J (1999) Biological individuality. The identity and persistence of living entities. Cambridge University Press, Cambridge
Woodger JH (1952) From biology to mathematics. Br J Philos Sci 3:1–21
Ziehen Th (1934) Erkenntnistheorie. Zweite Auflage. Erster Teil. Allgemeine Grundlegung der Erkennnistheorie. Spezielle Erkenntnistheorie der Empfindungstatsachen einschliesslich Raumtheorie. Fischer, Jena
Acknowledgments
This paper was first presented at the workshop, Perspectives on the Tree of Life, sponsored by the Leverhulme Trust and held in Halifax, Nova Scotia, July, 2009. I thank Eric Bapteste for invaluable guidance to the literature; he and two anonymous reviewers offered much appreciated comments and criticism of an earlier version of this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rieppel, O. The series, the network, and the tree: changing metaphors of order in nature. Biol Philos 25, 475–496 (2010). https://doi.org/10.1007/s10539-010-9216-4
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10539-010-9216-4