
Abstract
Animal studies have been instrumental in providing knowledge about the molecular and neural mechanisms underlying drug addiction. Recently, the fruit fly Drosophila melanogaster has become a valuable system to model not only the acute stimulating and sedating effects of drugs but also their more complex rewarding properties. In this review, we describe the advantages of using the fly to study drug-related behavior, provide a brief overview of the behavioral assays used, and review the molecular mechanisms and neural circuits underlying drug-induced behavior in flies. Many of these mechanisms have been validated in mammals, suggesting that the fly is a useful model to understand the mechanisms underlying addiction.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Drug addiction is a disorder characterized by excessive use of a drug to the point of compulsive drug seeking and consumption. The American Psychiatric Association (DMS-IV) differentiates between substance abuse, considered an earlier stage of addiction, and substance dependence. Substance abuse is defined as continued drug use despite interpersonal problems, legal problems, failure to fulfill obligations, or physically hazardous situations. The criteria for substance dependence include physical symptoms, such as tolerance and withdrawal, as well as signs of uncontrolled use, which include giving up normal activities and continued use despite knowledge of self-harm and the desire to stop. These definitions highlight the fact that addiction is an exclusively human phenomenon. However, animal models have been used to study specific aspects of addiction, and have proved invaluable in understanding the underlying neural and molecular mechanisms.
Animal models allow the experimenter to focus on distinct components of the addiction process, ranging from simple, acute drug responses to more complex behaviors such as drug seeking, self-administration, and relapse. Each behavioral model has advantages and disadvantages. Whereas the more complex models likely have greater relevance to the human condition, assays for acute drug responses are simpler to perform and thus provide the potential for high-throughput analysis, facilitating the identification of the underlying mechanisms.
Although rodent models have provided crucial insights into the mechanisms underlying drug-related behaviors, they are not ideal for unbiased, forward genetic approaches aimed at identifying novel and unsuspected mechanisms. This is due primarily to the expense and time required for animal maintenance, breeding, and behavioral analyses. In contrast, the fruit fly Drosophila melanogaster is one of the most genetically and experimentally accessible model organisms in biology. In this review the terms Drosophila and flies will refer exclusively to this species. While for many years flies were used primarily to identify the molecular and neural mechanisms regulating acute drug responses, the recent development of assays that measure drug self-administration and reward has allowed the analysis of these more complex behaviors.
Drosophila as a model system to study behavior
Drosophila has been used to gain insight into molecular, cellular, developmental, and disease processes that are conserved in mammals, including humans, as most of these fundamental biological mechanisms are shared throughout the animal kingdom. Although mammals have two to three times as many genes as flies, they have approximately the same number of gene families (Holland 2003). About 75% of human disease genes have related sequences in Drosophila, suggesting that flies can serve as an effective model to study the function of a wide array of genes involved in human disease (Adams et al. 2000; Reiter et al. 2001). The nervous system of the fly comprises approximately 300,000 neurons including a brain, ventral nerve cord (the equivalent of the spinal cord), and peripheral nervous system. Despite their relatively small number of neurons in comparison with mammals, flies exhibit many complex behaviors such as associative learning, sensorimotor integration, and social behaviors (Quinn et al. 1974; Pick and Strauss 2005; Greenspan and Ferveur 2000; Chen et al. 2002).
The classical advantages of using Drosophila include factors such as cost, size, fecundity, and timescale. First, flies are easy and inexpensive to rear in the laboratory using small vials or bottles and a yeast-based food medium. Due to their small size, thousands of genotypes of flies can be maintained in a typical laboratory. Second, due to their high fecundity, hundreds of flies can be obtained from a single female. Third, flies have a rapid life cycle, requiring only 10 days at 25°C to develop from egg to mature adult.
For these reasons, flies have long represented an ideal organism to conduct mutagenesis screens to isolate genes regulating a particular biological process of interest (“forward genetics”, i.e. going from phenotype to gene). The advent of genetic transformation in the 1980s also allowed for “reverse genetics” (i.e. going from gene to phenotype) by allowing researchers to introduce specific genes of interest into a fly (Rubin and Spradling 1982). The subsequent sequencing and annotation of the Drosophila genome have greatly facilitated both of these approaches (Adams et al. 2000).
In recent years, the generation of large collections of publicly available mutants and other transgenic tools has allowed for the functional study of nearly any fly gene of interest. The traditional use of X-ray or chemical mutagenesis is becoming gradually supplanted by insertional mutagenesis, in which a transposable genetic element creates a mutation by inserting into a random genomic site, and the gene affected can be easily identified by sequencing the flanking DNA (Bingham et al. 1981). Several groups have now generated large mutant collections for which the insertion site in each mutant has been sequenced (Bellen et al. 2004; Thibault et al. 2004; Schuldiner et al. 2008). In addition, an RNA interference (RNAi) library has been generated in which each fly line contains an inducible RNAi construct for silencing a single fly gene, with nearly 90% of the fly genome represented (Dietzl et al. 2007).
Some of the genetic tools developed in Drosophila have particular relevance to studying the relationship between genes, the brain, and behavior. For example, genetic tools in flies allow one to manipulate the nervous system independently of other tissues in the body. Furthermore, because different neural circuits may have distinct and perhaps opposing roles in behavior, one would ideally like to target specific sets of neurons within the brain. This cellular specificity can be accomplished by the bipartite Gal4/UAS system, in which the transcriptional activator Gal4 is expressed in a spatially restricted pattern and activates any gene placed downstream of the upstream activating sequence (UAS) (Brand and Perrimon 1993; Fig. 1a). The generation and characterization of thousands of Gal4 lines expressed in various patterns allow for manipulation of specific brain regions or neuronal types (Pfeiffer et al. 2008). This technique allows one to ask in which neurons a particular gene functions to regulate a behavioral response. These patterns can be further spatially refined to very small subsets of neurons using the “split Gal4 system” in which the DNA-binding and transcriptional-activation domains of Gal4 are targeted to different neuronal subsets using different promoters; transcriptional activation of target genes occurs only in neurons expressing both domains (Luan et al. 2006). Temporal specificity can be achieved by using a temperature-sensitive Gal4 repressor called Gal80ts and shifting the flies from the permissive to the restrictive temperature during a particular time period (McGuire et al. 2003; Fig. 1b).

Common genetic tools in Drosophila. a The Gal4/UAS system (Brand and Perrimon 1993). The transcriptional activator Gal4 is expressed in a spatially restricted pattern and activates any gene placed downstream of the upstream activating sequence (UAS). b The TARGET system (McGuire et al. 2003). At the restrictive temperature (30°C), Gal80ts is inactive, Gal4 is active and UAS-driven genes are expressed. At the permissive temperature (19°C), Gal80ts is active, Gal4 is inhibited, and UAS-driven genes are not expressed. c The Shibirets system (Kitamoto 2001). At the restrictive temperature (30°C), but not the permissive temperature (19°C), Shits blocks neurotransmission by disrupting endocytosis and thereby depleting synaptic vesicles. d The TrpA1 system (Hamada et al. 2008; Pulver et al. 2009). At the restrictive temperature (27°C), but not the permissive temperature (19°C), cation flow through the temperature-gated cation channel dTRPA1 causes neuronal depolarization
In addition to studying the function of genes within the nervous system, the Gal4/UAS system is well suited to studying neural circuit function. UAS lines are available in which proteins that inducibly control neuronal activity can be expressed, allowing one to activate or silence a particular set of neurons during a specific behavioral task. For example, neurons can be silenced using Shibirets, a temperature-sensitive dynamin allele that blocks synaptic vesicle recycling (Fig. 1c), and neurons can be activated using TrpA1, a temperature-sensitive cation channel that causes neuronal depolarization (Fig. 1d) (Kitamoto 2001; Hamada et al. 2008). Furthermore, the development of a second binary system in addition to the Gal4/UAS system, the LexA/LexAop system (Lai and Lee 2006), allows for the independent manipulation of multiple neural circuits, such as activating some neurons while inhibiting others. Thus, flies have now become a leading model organism for studying not only the molecular mechanisms but also the neural circuits that underlie behavior.
Models to study ethanol-related behaviors in flies
Ethanol is the drug that has been by far the most intensively studied in Drosophila, and will therefore be the main focus of this review. Ethanol is a commonly abused psychoactive drug that can produce both short-term behavioral impairment as well as long-term addiction. Unlike other drugs such as cocaine and nicotine, ethanol does not act on a single molecular target but instead is thought to affect a variety of molecules, including multiple ion channels (Koob 2004). Fruit flies encounter ethanol in their natural environment since one of the main metabolites in fermenting fruit is ethanol. Ethanol can act as a long-distance signal to draw flies to rotting fruit, as flies are attracted to low concentrations of ethanol vapor (Dudley 2002; Hoffmann and Parsons 1984). Female flies prefer to lay their eggs in media containing up to 5% ethanol (McKenzie and Parsons 1972), and larvae efficiently metabolize ethanol and use it as a food source (Geer et al. 1993).
Despite this long-standing relationship between Drosophila and ethanol, the molecular underpinnings of the effects of ethanol on fly behavior were not investigated until relatively recently. Several types of ethanol-related behaviors have now been characterized in flies, with the goal of using the abundant genetic tools in Drosophila to understand the underlying mechanisms. These behaviors range from simple to complex: (1) acute locomotor responses to ethanol, (2) ethanol tolerance following an initial exposure, and (3) ethanol preference and conditioned preference behaviors that model specific facets of addiction.
Acute ethanol sensitivity
Flies exhibit acute responses to ethanol exposure that are quite similar to those of mammals, including humans (Morean and Corbin 2010). There is evidence in humans as well as mammalian models that sensitivity to acute ethanol-induced motor impairment correlates inversely with ethanol consumption and risk of abuse, and that the same genes can influence both types of behavior (Schuckit 1994; Morean and Corbin 2010; Kurtz et al. 1996; Thiele et al. 1998; Hodge et al. 1999). Studying these simpler ethanol responses, which are often easier to test in the laboratory, is therefore likely to provide insight into the mechanisms regulating more complex addiction-related behaviors as well.
To measure acute ethanol responses in flies, ethanol is typically administered in the form of pure ethanol vapor mixed with air at a specified ratio, allowing one to control the ethanol concentration that the flies receive (Wolf et al. 2002). Ethanol can also be administered to flies by injection, though few studies have employed this technique (Dzitoyeva et al. 2003). Low to moderate concentrations of ethanol induce locomotor hyperactivity, which can be measured by filming the flies and using tracking software to identify the flies and calculate their locomotor speed (Wolf et al. 2002).
In contrast, high concentrations of ethanol elicit loss of postural control and eventually sedation (Moore et al. 1998; Rothenfluh et al. 2006; Corl et al. 2009). Loss of postural control was initially assayed in the inebriometer, a vertical column containing mesh baffles (Weber 1988; Cohan and Graf 1985; Moore et al. 1998; Fig. 2a). Flies naturally exhibit negative geotaxis and therefore tend to remain at the top of the column, but as they lose postural control they gradually fall from one baffle to the next. Ethanol sensitivity can therefore be measured as the time required for the flies to reach the bottom of the column. Negative geotaxis has also been directly assayed as a measure of ethanol sensitivity by quantifying the vertical distance that flies climb after being knocked to the bottom of a vial (Bhandari et al. 2009). More recently, ethanol-induced loss of postural control (referred to more simply as “sedation”) has been assayed manually using a loss-of-righting reflex assay, in which one counts the number of flies that fail to regain upright posture after being knocked over (Fig. 2b) (Rothenfluh et al. 2006; Corl et al. 2009).

Assays to measure alcohol-induced behavior in Drosophila. a The inebriometer measures ethanol-induced loss of postural control by measuring the time required for flies to fall down the mesh baffles from the top to the bottom of the column (Weber 1988; Moore et al. 1998). b The booz-o-mat allows for the measurement of ethanol-induced hyperactivity and sedation while streaming vaporized ethanol into horizontal tubes containing groups of flies. Hyperactivity is measured by filming the flies and using tracking software to calculate their locomotor speed. Sedation is measured by recording the time required for flies to exhibit the loss-of-righting reflex (Wolf et al. 2002). c The two-choice CAFE assay measures consumption preference for food containing ethanol compared to normal food (Ja et al. 2007; Devineni and Heberlein 2009). d Conditioned ethanol preference is measured by training the flies in a sealed container to associate a neutral odor with the presence of an intoxicating dose of ethanol, and later testing preference for that odor in the absence of ethanol using a Y-maze (Kaun et al. 2011)
Ethanol tolerance
In flies, as in mammals, repeated exposure to ethanol induces tolerance, which is defined as an acquired resistance to the effects of the drug. Tolerance is one of the DSM-IV criteria for alcohol dependence (American Psychiatric Association 1994) and has been correlated with heavy drinking and alcohol abuse (Schuckit et al. 2008). In flies, ethanol tolerance is assayed by measuring the decrease in sensitivity to ethanol-induced motor impairment after ethanol pre-exposure. This change in sensitivity can be assayed using the inebriometer (Scholz et al. 2000), the sedation assay (Berger et al. 2004), or negative geotaxis (Bhandari et al. 2009), and can be measured as a change in either the proportion of impaired flies (Urizar et al. 2007), latency or duration of impairment (Dzitoyeva et al. 2003; Devineni et al. 2011), or the recovery time after sedation (Berger et al. 2004; Cowmeadow et al. 2005). Ethanol tolerance appears to be robust to variations in the tolerance protocol, as the studies cited above vary significantly in the timing and concentration of ethanol exposure.
Two types of ethanol tolerance have been characterized in flies, termed rapid and chronic. Rapid tolerance is induced by relatively brief exposure to a sedating concentration of ethanol, while chronic tolerance is induced by prolonged (~24 h) exposure to a low ethanol concentration that does not produce overt intoxication (Berger et al. 2004). Rapid and chronic tolerance are mediated by distinct mechanisms; for example, only chronic tolerance is dependent on protein synthesis (Berger et al. 2004).
Voluntary ethanol consumption
The behavioral assays described above were designed to study relatively simple behaviors associated with ethanol exposure, and much has been learned from these assays regarding the molecular mechanisms underlying the effects of ethanol. However, in order to relate these discoveries to addiction-related processes, more complex assays that more closely mimic mammalian behavior are needed. A recently developed ethanol self-administration assay demonstrates that flies prefer to consume ethanol-containing food over regular food and that this preference exhibits several features reminiscent of compulsive alcohol consumption (Ja et al. 2007; Devineni and Heberlein 2009).
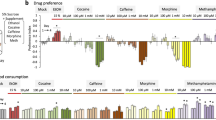
Ethanol consumption can be measured in flies using a two-choice assay similar to the two bottle choice assay used in rodent studies (Fig. 2c) (Devineni and Heberlein 2009). Flies exhibit a robust, dose-dependent preference for food containing 5–25% ethanol. This ethanol preference cannot be entirely explained by either chemosensory or caloric attraction to ethanol; ethanol preference persists in the absence of olfactory or gustatory input, and preference is not altered by varying the relative caloric content of the solutions (Devineni and Heberlein 2009). Flies also show increased ethanol consumption over time, and, when food deprived, voluntarily consume ethanol to pharmacologically relevant concentrations. Importantly, flies demonstrate two additional criteria of addiction-like behavior; they will overcome an aversive stimulus, the bitter-tasting compound quinine, in order to consume ethanol, and they rapidly return to high levels of ethanol consumption after ethanol deprivation, modeling a relapse-like effect (Devineni and Heberlein 2009).
Conditioned preference for ethanol reward
To test directly whether intoxicating doses of ethanol are rewarding to flies, a conditioned ethanol preference assay was recently developed (Kaun et al. 2011; Fig. 2d). In this assay, flies are initially exposed to two neutral odor cues, one of which is paired with a moderately intoxicating exposure to ethanol vapor. Flies are later offered a choice between the two odors, and preference for the ethanol-associated odor is measured. Similar to mammalian conditioned place preference (CPP) models, this assay uses conditioned preference to assess the rewarding properties of ethanol intoxication. This assay has some advantages over the ethanol self-administration assay: (1) the ethanol concentration that the flies experience can be controlled by the experimenter and (2) the ethanol stimulus is removed during the test, allowing measurement of the rewarding value of the drug rather than immediate preference for the drug.
When flies have been trained to associate an odor cue with ethanol intoxication, they show initial aversion to the cue, which, within 12–15 h, transforms into a long-lasting preference (Kaun et al. 2011). The development of conditioned preference is dependent on the ethanol concentration; preference is induced only by exposure to moderate ethanol doses that induce locomotor hyperactivity. Conditioned preference is not induced by lower ethanol concentrations that fail to elicit behavioral changes or higher concentrations that cause sedation. Thus, flies seem to require an intoxicating, but not sedating, dose of ethanol for it to be remembered as rewarding. Remarkably, flies will endure electric shock in order to attain the cue associated with ethanol, indicating that they are willing to tolerate punishment to seek the drug (or, in this case, a cue that predicts the presence of the drug) (Kaun et al. 2011). This response is reminiscent of compulsive behavior such as impaired response inhibition observed in mammalian studies of drug reward. Furthermore, flies will endure a stronger shock intensity to attain a cue associated with ethanol than a cue associated with sugar, suggesting that the preference for ethanol is distinct from a preference for food reward (Kaun et al. 2011).
Molecular mechanisms underlying ethanol-induced behavior in Drosophila
As described above, an array of assays has been established to study various aspects of ethanol-induced behavior in Drosophila. All of these assays are relatively simple, robust, and high-throughput, allowing researchers to conduct forward genetic screens to identify the underlying mechanisms. The genes identified in these screens have led to the characterization of diverse molecular and cellular processes that mediate ethanol-induced behavior in flies.
Molecular pathways
Several classical molecular signaling pathways have been implicated in regulating sensitivity to ethanol-induced motor impairment in flies, including the epidermal growth factor receptor (EGFR), phosphoinositide 3-kinase (PI3K)/Akt, and cyclic adenosine monophosphate (cAMP) pathways. Genetic and pharmacological evidence indicates that the EGFR and cAMP pathways promote resistance to ethanol sedation (Corl et al. 2009; Moore et al. 1998), while the PI3K/Akt pathway enhances ethanol sedation (Eddison et al. 2011). However, these pathways are likely to regulate ethanol sensitivity in more complex ways depending on factors such as the cell types in which they are expressed and the presence of multiple protein isoforms. For example, a mutation disrupting the function of the type II regulatory subunit of protein kinase A (PKA), a key effector of cAMP signaling, causes the opposite effect on ethanol sedation as expected from previous manipulations that disrupt overall PKA signaling (Park et al. 2000; Moore et al. 1998).
A genetic screen revealed that scabrous (sca), encoding a secreted protein that negatively regulates the Notch signaling pathway (Baker et al. 1990; Powell et al. 2001), is required for ethanol reward memory. Notch signaling is required for long-term memory formation in flies, suggesting that sca may contribute generally to long-term memory processes (Presente et al. 2004; Ge et al. 2004). However, a mutation of sca that affects ethanol reward memory does not affect short-term memory (LaFerriere et al. 2008). Notch signaling has been shown to regulate migration, morphology, synaptic plasticity and survival of immature and mature neurons (Ables et al. 2011). It will be interesting to examine how sca and the Notch pathway affect the neural plasticity underlying memory for ethanol reward.
Transcription and translation have been implicated in ethanol-induced behaviors. Two genes encoding putative transcriptional regulators, Drosophila LIM-domain only (dLmo) and hangover (hang), were identified as regulators of ethanol sedation and ethanol tolerance, respectively (Lasek et al. 2011a; Scholz et al. 2005). One likely target gene whose expression is regulated by dLmo, the Drosophila homolog of anaplastic lymphoma kinase (dAlk), has been identified and shown to regulate ethanol sedation (Lasek et al. 2011b).
A mutation in krasavietz (kra), which encodes a predicted translation initiation factor that inhibits protein translation in vitro (Lee et al. 2007), causes decreased sensitivity to ethanol-induced sedation, decreased rapid and chronic ethanol tolerance, and decreased ethanol consumption (Berger et al. 2008; Devineni and Heberlein 2009). Although protein synthesis is required for chronic tolerance, it is not required for the development of rapid tolerance and is unlikely to occur during the brief timescale of acute ethanol intoxication (~30 min; Berger et al. 2004). However, treating flies with a protein synthesis inhibitor prior to ethanol exposure caused pronounced resistance to ethanol impairment (Berger et al. 2004), suggesting that some proteins that are constitutively synthesized in the absence of ethanol mediate naive ethanol sensitivity. The role of protein synthesis in ethanol consumption has not been directly tested.
Finally, molecular pathways involved in cellular stress responses have been implicated in ethanol tolerance. hang mutants, which show decreased ethanol tolerance, also show sensitivity to oxidative stress and decreased heat-ethanol cross-tolerance (i.e. tolerance to ethanol stimulated by heat shock stress instead of ethanol exposure; Scholz et al. 2005). The gene jwa, a retinoic acid-responsive gene whose product associates with the cytoskeleton, mediates oxidative and heat stress responses and also promotes ethanol tolerance (Li et al. 2008). Since high doses of ethanol induce cellular stress, which in some ways mimics oxidative and heat stress, it may not be surprising that common molecular pathways respond to ethanol as well as other stressors (e.g. Wu and Cederbaum 2009; Wilke et al. 1994; Piper 1995).
Cellular mechanisms
One of the key cellular processes that has been implicated in ethanol-induced behaviors in flies is cytoskeletal dynamics. thousand and one (tao), which was identified as a critical regulator of ethanol-induced hyperactivity, implicated microtubule dynamics in the hyperactivity response (King et al. 2011). tao was shown to function through the conserved kinase PAR-1 (also called MARK in mammals) to regulate the microtubule-binding protein Tau during fly brain development (Matenia and Mandelkow 2009; King et al. 2011). The mouse homolog of jwa, which promotes ethanol tolerance in flies (see above), is also a microtubule-associated protein (Li et al. 2008; Chen et al. 2007).
In addition to microtubule organization, the regulation of actin has been implicated in ethanol responses. Rho GTPase activator protein 18B (RhoGAP18B) regulates sensitivity to both ethanol-induced sedation and hyperactivity through different protein isoforms (Rothenfluh et al. 2006). RhoGAP18B is a GTPase activating protein that regulates ethanol sensitivity by functioning through Rho family GTPases, which are key regulators of actin dynamics (Rothenfluh et al. 2006). Additionally, Kra (described above) interacts with the crosslinking protein Short stop to regulate actin organization, suggesting that that actin regulation may underlie some of its diverse effects on ethanol-induced behavior (Lee et al. 2007; Sanchez-Soriano et al. 2009). However, it is important to note that changes in neither microtubule nor actin organization have been directly linked to altered ethanol responses in these mutants.
Finally, the integrin class of cell adhesion molecules has been implicated in ethanol-induced behaviors. Mutations in the alpha-integrin gene scab (scb) or the β-integrin gene myospheroid (mys) cause increased ethanol sensitivity as well as increased tolerance (Bhandari et al. 2009). It will be interesting to determine the mechanisms by which disruptions in cytoskeletal organization or cell adhesion lead to altered ethanol responses.
Synaptic function and neuronal excitability
Synapse number has recently been implicated in ethanol sensitivity in flies. Several genetic manipulations that lead to increased ethanol sedation sensitivity, such as mutations in arouser (aru) or amnesiac (amn) and overexpression of PI3K or Ras homolog enriched in brain (Rheb), also increase synapse number at the larval neuromuscular junction (NMJ) and/or the adult central brain (Eddison et al. 2011). An environmental manipulation, adult social isolation, which is known to reduce the number of synapses of a specific set of fly brain neurons (Donlea et al. 2009), also reduces ethanol sensitivity (Eddison et al. 2011). Furthermore, social isolation concurrently restores normal synapse number and ethanol sensitivity to the ethanol-sensitive mutant aru (Eddison et al. 2011). This correlation between synapse number and ethanol sedation sensitivity using multiple independent genetic and environmental manipulations suggests that increased synapse number may directly promote increased sensitivity to ethanol sedation.
In addition to providing a novel cellular mechanism by which ethanol behaviors can be regulated, these findings suggest the hypothesis that ethanol tolerance may result from a compensatory decrease in synapse number induced by the initial ethanol exposure. While this hypothesis has not been directly tested, hang, which promotes ethanol tolerance (see above), negatively regulates synapse number at the larval NMJ (Schwenkert et al. 2008). For both ethanol sensitivity and tolerance, it remains to be investigated whether increased synapse number translates into increased postsynaptic excitation (or inhibition), and whether ethanol sensitivity depends on increased synapse number generally throughout the nervous system or in specific neurons.
There is abundant evidence that synaptic transmission regulates ethanol-induced behaviors in flies. Flies carrying a mutation in Syntaxin 1A (Syx1A) or shibire (shi, encoding Drosophila dynamin), which are, respectively, required for synaptic vesicle docking and recycling, show defects in ethanol sedation tolerance (Krishnan et al. 2011). Use of conditional mutations revealed that normal synaptic vesicle release is required immediately after initial ethanol exposure rather than after recovery from intoxication to promote tolerance (Krishnan et al. 2011). A different study showed that flies lacking Synapsin (Syn), encoding a presynaptic vesicle scaffolding protein, unexpectedly show increased ethanol tolerance (Godenschwege et al. 2004). This result may reflect the fact that Synapsin is involved not only in regulating neurotransmitter release but also in neurite growth, synaptic formation and maturation, and in segregating the reserve and readily releasable pools of vesicles (Cesca et al. 2010). Finally, Homer, a protein that interacts with postsynaptic scaffolding and signaling proteins, including metabotropic glutamate receptors, regulates both initial ethanol sensitivity and ethanol tolerance (Urizar et al. 2007).
The major regulators of neuronal excitability that have been implicated in ethanol-induced behaviors in flies are the γ-aminobutyric acid B (GABAB) receptors and the large conductance calcium-activated potassium (BK) channels. GABAB receptor activity promotes sensitivity to ethanol sedation but reduces rapid ethanol tolerance (Dzitoyeva et al. 2003). As in mammals, Drosophila GABAB receptors are metabotropically coupled to potassium channels, thereby inhibiting neuronal excitability due to potassium efflux (Mezler et al. 2001). The BK channel encoded by the gene slowpoke (slo) has also been implicated in rapid ethanol tolerance, but in the opposite direction. Expression of the fly BK channel is upregulated by ethanol exposure and its function is required for the development of rapid ethanol tolerance; induction of BK channel expression is in fact sufficient to induce ethanol resistance, mimicking the tolerant state (Cowmeadow et al. 2005, 2006). The fact that GABAB receptors and BK channels likely affect neuronal excitability in the same direction, but regulate tolerance in opposite ways, suggests that they may function in different subsets of neurons that exert opposing effects on behavior. Alternatively, it has been proposed that BK channels may in fact enhance neuronal excitability by reducing the refractory period or enhancing firing rates, allowing neurons to compensate for the depressant effect of ethanol during sedation (Atkinson 2009).
In addition to classical neurotransmitters such as GABA, neuromodulators, including biogenic amines and neuropeptides, also regulate ethanol-induced behavior in flies. Dopamine promotes ethanol hyperactivity through the D1-like receptor DopR (Bainton et al. 2000; Kong et al. 2010b) and is also required for conditioned ethanol preference (Kaun et al. 2011). Octopamine, a biogenic amine thought to be the invertebrate analog of norepinephrine, is essential for the development of rapid but not chronic ethanol tolerance (Scholz 2000; Berger et al. 2004). Two neuropeptides produced in the fly brain, neuropeptide F (NPF) and insulin, have been shown to regulate ethanol sedation. NPF, the fly homolog of neuropeptide Y, enhances ethanol sedation (Wen et al. 2005). Mutations in the insulin receptor (InR) cause increased sedation sensitivity, as does overexpression of the adaptor protein p60 to inhibit the coupling between the insulin receptor (InR) and PI3K, the main effector of insulin signaling (Corl et al. 2005). These results indicate that insulin acts through PI3K to promote sedation resistance. However, a different study (discussed earlier) using several more direct manipulations of the PI3K/Akt pathway demonstrated that this pathway promotes sedation sensitivity (Eddison et al. 2011). PI3K may therefore have opposing roles in regulating ethanol sedation depending on the upstream molecule to which it is coupled and the cell type in which it is expressed. In general, the mechanisms by which these neuromodulators affect postsynaptic and/or presynaptic cells have not yet been characterized. It thus remains an open question whether they directly affect postsynaptic excitability or modulate other pre- or postsynaptic properties.
Genome-wide studies
The majority of the genes discussed above were identified using genetic screens in which mutants exhibiting abnormal behavior were isolated. However, an alternative approach is to use transcriptional profiling to compare gene expression under different conditions. For example, one study identified genes differentially expressed in fly strains selected for increased versus decreased sensitivity to ethanol, and confirmed that mutations in many of these genes cause altered ethanol sensitivity (Morozova et al. 2007).
Three studies have identified genes whose expression is regulated by ethanol exposure, making them good candidates for mediating the development of tolerance. These studies used varying exposure protocols and collectively identified 1,669 candidate genes, 29 of which were common to all three studies and 229 of which were common to at least two out of three studies (Morozova et al. 2006; Urizar et al. 2007; Kong et al. 2010a). Many of these genes were functionally validated using mutant analysis (Morozova et al. 2006; Kong et al. 2010a), but in most cases the molecular and cellular mechanisms by which these genes function have not been determined.
Neural circuits underlying ethanol-induced behavior in Drosophila
Although the neural circuits mediating ethanol-induced behaviors in Drosophila have not been as extensively studied as the molecular mechanisms, new tools such as Gal4 lines to target particular neurons and transgenes to manipulate neuronal activity have made the study of circuits more accessible.
In mammals, dopamine is an important regulator of many ethanol-related behaviors (Soderpalm et al. 2009). In the fly, dopamine is expressed in several clusters of neurons that project to a variety of brain regions (Nassel and Elekes 1992). As in mammals, many of these dopaminergic cells have been shown to play a role in ethanol-related behaviors. The function of dopamine in regulating ethanol hyperactivity was localized to a pair of dopaminergic neurons projecting to DopR-expressing neurons in the ellipsoid body of the central complex (Kong et al. 2010b), a region known to regulate visual and locomotor behavior, arousal, and memory (Martin et al. 1999; Wu et al. 2007; Neuser et al. 2008; Ofstad et al. 2011). The ellipsoid body is also the site of Homer function in the regulation of ethanol sedation sensitivity and tolerance (Urizar et al. 2007), though it is unknown whether Homer functions via DopR signaling, or in DopR-expressing neurons.
Dopamine neurons also mediate conditioned ethanol preference. Ethanol reward memory, like other forms of memory, can be divided into three phases: acquisition (memory formation during training), consolidation (the period between training and testing), and retrieval (expression of the memory during testing) (Krashes et al. 2007). Interestingly, silencing dopaminergic neurotransmission impairs retrieval, but not acquisition or consolidation, of ethanol reward memory (Kaun et al. 2011).
While some dopaminergic neurons innervate the ellipsoid body, others terminate in the mushroom body, a brain structure implicated in olfactory processing and learning (Davis 2011). Neurotransmission of mushroom body neurons is required for both ethanol-induced hyperactivity and conditioned ethanol preference (King et al. 2011; Kaun et al. 2011). Both behaviors are mediated by neurons in specific subregions within this structure, and distinct phases of conditioned ethanol preference are in fact localized to different mushroom body neurons (King et al. 2011; Kaun et al. 2011). Together, these studies demonstrate that different ethanol-induced behaviors can be mapped to distinct neural loci, and that some brain structures, such as the mushroom body, are important for multiple behaviors.
Mammalian validation of mechanisms underlying ethanol-induced behavior
Now that years of research have implicated many different molecular and cellular pathways in mediating fly responses to ethanol, it is important to ask whether these mechanisms function in mammals as well. In fact, many of the genes and molecular pathways implicated in Drosophila ethanol responses play a similar role in mammals (see Table 1). For example, the cAMP, EGFR, and NPF/NPY pathways all regulate ethanol sensitivity similarly in flies and rodents (Moore et al. 1998; Wand et al. 2001; Corl et al. 2009; Wen et al. 2005; Thiele et al. 1998). Furthermore, these pathways regulate not only ethanol sensitivity but also ethanol consumption in rodents (Wand et al. 2001; Corl et al. 2009; Thiele et al. 1998). Thus, simple behavioral assays that are readily used for genetic screening in flies can yield candidate genes that have homologous roles in rodent models. Moreover, an FDA-approved drug that inhibits the function of EGFR, a molecule first shown to regulate ethanol-related behavior in the fly, has been shown to be effective in a preclinical rat model of ethanol addiction (Corl et al. 2009).
While most of the genes affecting ethanol-induced behavior in flies have not yet been tested for a role in humans, a few have already been associated with human ethanol-related behavior. Polymorphisms in the human ALK gene are correlated with multiple measures of ethanol sensitivity (Lasek et al. 2011b), and polymorphisms in one human homolog of hang, ZNF699, were found to be associated with alcohol dependence (Riley et al. 2006). Recently, a genome-wide meta-analysis revealed that polymorphisms in autism susceptibility candidate 2 (AUTS2) are associated with alcohol consumption (Schumann et al. 2011). Mice selected for high versus low alcohol consumption differ in expression of AUTS2, and downregulation of the fly homolog of AUTS2 leads to reduced ethanol sensitivity (Schumann et al. 2011). Given the significant conservation of genes affecting ethanol responses in flies and rodents, it is likely that additional genes identified in flies will be validated in rodents and humans, and vice versa.
In addition to molecular pathways, some of the cellular mechanisms implicated in Drosophila ethanol responses have also been studied in mammals. For example, the role of synapse function in ethanol-induced behavior is still an emerging field of study in flies, while the effects of ethanol at the synapse have been well studied in mammals. Ethanol acts on a variety of postsynaptic receptors, most notably GABAA and N-methyl-d-aspartic acid (NMDA) receptors, and also exerts presynaptic effects on neurotransmitter release (Siggins et al. 2005). Whether changes in synapse number are associated with altered ethanol behaviors in mammals, as is the case in flies (Eddison et al. 2011), has not yet been studied.
While at the molecular level flies and mammals share many features (Littleton and Ganetzky 2000; Lloyd et al. 2000), the anatomical organization of fly and mammalian nervous systems is quite distinct. It is therefore difficult to draw parallels between the neural circuits that regulate ethanol-induced behavior in flies and mammals. In flies, brain structures such as the ellipsoid body and the mushroom body have been implicated in various ethanol responses; it is unclear what the equivalent structures are in the mammalian brain. Nevertheless, certain conserved neurochemical systems function similarly in flies and mammals. The mammalian mesolimbic dopamine pathway, including its target regions, is perhaps the most intensely studied neural circuit in the context of alcohol reward and addiction (Soderpalm et al. 2009). Dopamine neurons were similarly found to be required for ethanol hyperactivity and reward in Drosophila (Kong et al. 2010b; Kaun et al. 2011). Neuropeptidergic systems, such as the NPY/NPF system, also regulate ethanol responses similarly in flies and rodents, as discussed above. Thus, the functions of neurochemically defined neural pathways, rather than morphologically defined brain regions, are likely to be conserved in regulating ethanol behaviors.
Study of other drugs of abuse in Drosophila
Drugs of abuse other than ethanol have not yet been studied extensively in flies. This may be due in part to the fact that ethanol, in vapor or liquid form, can be delivered to flies quite readily and in a reproducible manner; the same is not true for drugs such as cocaine and nicotine (McClung and Hirsh 1998; Bainton et al. 2000). Although there are assays to measure the locomotor effects of some of these other drugs, there are currently no assays to investigate their rewarding and addiction-like properties. Nevertheless, several genes and molecular mechanisms regulating drug-induced behaviors have been discovered using simple locomotion assays (Table 2).
Cocaine
Cocaine is an addictive psychostimulant that causes enhanced locomotor activity and stereotypy (repetitive behavior) in mammals (Satel et al. 1991). When flies are exposed to volatilized cocaine, they show similar behavioral effects including continuous grooming at low doses, circling and aberrant walking behavior at intermediate doses, and fast, uncontrolled movements followed by body tremors and akinesia at high doses (McClung and Hirsh 1998). Repeated cocaine exposure causes flies to become increasingly sensitive to the behavioral effects of the drug, a process referred to as sensitization (McClung and Hirsh 1998). In addition to direct observation, other assays have been developed for greater control of drug delivery and simpler behavioral analysis. For example, the “crackometer” quantifies the loss of negative geotaxis and positive phototaxis (two robust innate behaviors in flies) under the influence of volatilized cocaine (Bainton et al. 2000). Semi-automation of this assay allows for high-throughput behavioral analysis (George et al. 2005). Finally, locomotor tracking systems allow for quantification of locomotor speed and pattern (Bainton et al. 2000; Dimitrijevic et al. 2004).
Unlike ethanol, cocaine acts primarily on a single class of molecular targets: it inhibits monoamine transporters, thereby increasing synaptic levels of monoamines including dopamine, serotonin, epinephrine, and norepinephrine. Inhibition of the dopamine transporter (DAT), in particular, is largely responsible for cocaine-induced locomotor hyperactivity in mammals (Giros et al. 1996). It is therefore not surprising that dopamine signaling is also required for cocaine-induced hyperactivity in flies. A key role for the dopaminergic system in mediating the effect of cocaine has been demonstrated through both pharmacological and genetic methods. Pharmacological reduction of dopamine levels or dopamine receptor function causes decreased locomotor hyperactivation by cocaine, suggesting that cocaine induces hyperactivity by increasing dopaminergic transmission (Bainton et al. 2000; Torres and Horowitz 1998; Yellman et al. 1998). However, genetic studies provide conflicting results: (1) overexpression of one isoform of the vesicular monoamine transporter (VMAT-A), which is expected to increase dopamine levels, decreases cocaine sensitivity (Chang et al. 2006), and (2) constitutive inhibition of dopaminergic transmission, predicted to block the effects of cocaine, causes cocaine hypersensitivity (Li et al. 2000). In both cases these initially counterintuitive results could be explained by compensatory adaptations in postsynaptic dopamine signaling (Chang et al. 2006; Li et al. 2000).
Drosophila studies have identified unanticipated genes and pathways regulating cocaine-induced behavior. For example, mutations in the circadian genes period (per), clock (Clk), cycle (cyc), and doubletime/discs overgrown (dco) reduce behavioral sensitization to cocaine (Andretic et al. 1999). These genes may regulate cocaine sensitization by affecting dopaminergic signaling since cocaine-treated per mutants, unlike cocaine-treated wild-type flies, do not increase locomotion in response to a dopamine receptor agonist (Andretic et al. 1999). Circadian genes similarly mediate cocaine-induced behaviors in mammals. The mammalian homologs of per have been shown to regulate cocaine sensitization and CPP in mice (Abarca et al. 2002). In addition, mice lacking a functional Clock gene display increased cocaine reward and dopamine neuron excitability in the midbrain (McClung et al. 2005).
moody, which encodes two G protein-coupled receptors (GPCRs), was identified in a genetic screen for cocaine-induced loss of negative geotaxis (Bainton et al. 2005). moody mutant flies show enhanced cocaine sensitivity, and moody was shown to function in glia to regulate blood–brain barrier permeability (Bainton et al. 2005; Schwabe et al. 2005). loco, which encodes a regulator of G protein signaling (RGS) that terminates GPCR signaling, functions along with moody to regulate blood–brain barrier permeability (Schwabe et al. 2005). As predicted from its molecular function, loco regulates cocaine sensitivity in the opposite direction as moody (Granderath et al. 1999; Bainton et al. 2005). The mammalian homolog of loco, RGS4, has been implicated in psychostimulant use (Bishop et al. 2002; Gold et al. 2003; Schwendt et al. 2006, 2007). These studies suggest that blood–brain barrier permeability may play an important role in drug sensitivity and potentially addiction.
Genetic screens have revealed overlap between genes that regulate ethanol and cocaine sensitivity. RhoGAP18B, tao, and dLmo, three genes that were identified in genetic screens and shown to regulate ethanol sensitivity (see previous section), also regulate cocaine sensitivity (Rothenfluh et al. 2006; King et al. 2011; Tsai et al. 2004). Both RhoGAP18B and tao mutants exhibit cocaine resistance, while dLmo mutants show increased cocaine sensitivity. dLmo regulation of cocaine sensitivity has been mapped to a subset of the pigment-dispersing factor (PDF) neurons (Tsai et al. 2004), the primary circadian pacemaker cells in flies (Renn et al. 1999). The function of dLmo has been validated in mammals, as downregulation of one of the mammalian homologs, Lmo4, causes increased cocaine sensitivity and sensitization in mice (Lasek et al. 2010). Lmo4 may regulate cocaine sensitivity by acting through the dopamine D2 receptor (Drd2) or the GluR1 subunit of the AMPA receptor, both of which show decreased expression upon Lmo4 downregulation (Heberlein et al. 2009).
Amphetamines
Amphetamines are psychostimulants that, like cocaine, increase monoaminergic transmission. Amphetamines cause monoamine transporters to function in reverse, transporting these transmitters into the synapse (Koob and Nestler 1997). A small number of studies have characterized the effects of amphetamines in Drosophila, focusing on methamphetamine and 3,4-methylenedioxy-methamphetamine (MDMA, also known as ecstasy).
One study investigated the behavioral effects of orally administering methamphetamine to adult flies. Consistent with its stimulant effects in mammals, methamphetamine increased locomotor activity of flies, interrupted sleep, and also affected male courtship (Andretic et al. 2005). Methamphetamine administration was also associated with changes in visually evoked neural activity, suggesting that it may affect visual perception (Andretic et al. 2005). Methamphetamine showed interactions with the dopaminergic system in affecting this type of neural activity, consistent with its mode of action in mammals (Andretic et al. 2005).
In a different study, MDMA was orally administered to Drosophila larvae, which caused reduced feeding and locomotion (Dasari et al. 2007). It has not been determined whether MDMA also produces stimulant effects on Drosophila behavior. Larvae fed MDMA did not show an acute difference in levels of dopamine or serotonin, but did contain higher levels of both transmitters as adults, suggesting that monoamine synthesis may be upregulated over time in response to MDMA-induced monoamine depletion (Dasari et al. 2007).
Nicotine
Nicotine, the major addictive component of tobacco, affects mammalian behavior by activating nicotinic acetylcholine receptors (Nestler 2005). When exposed to volatilized nicotine, flies exhibit locomotor hyperactivity and spasmodic movements leading to grooming at low doses and hypokinesis and akinesia at higher doses (Bainton et al. 2000). Similar to cocaine, nicotine exposure dose-dependently impairs negative geotaxis in flies (Bainton et al. 2000). In mammals, the addictive properties of nicotine are thought to be mediated by both direct and indirect activation of dopaminergic neurons (Nestler 2005). The locomotor effects of nicotine in flies are similarly dependent on dopamine, as pharmacological depletion of dopamine reduces nicotine sensitivity (Bainton et al. 2000). Aside from dopamine, little is known about the molecular mechanisms mediating nicotine sensitivity in flies. However, several genes known to mediate cocaine sensitivity in flies have also been shown to regulate nicotine sensitivity: moody mutant flies are sensitive to the effects of both drugs, whereas RhoGAP18B and tao mutants are resistant (Bainton et al. 2005; Rothenfluh et al. 2006; King et al. 2011). These genes suggest that certain shared mechanisms may regulate multiple types of drug addiction in flies.
Conclusions and future directions
For many years flies have been used as a model to study acute drug responses, with a particular focus on ethanol, and the mechanisms underlying these behaviors have turned out to be remarkably conserved from flies to mammals. More recently, the development of new assays, in particular the voluntary ethanol consumption and conditioned ethanol preference assays, has demonstrated that flies exhibit addiction-like behavior. Flies fulfill several of the criteria proposed for an animal model of alcohol addiction (Cicero 1979; McBride and Li 1998). The ethanol consumption assay reveals that (1) flies voluntarily consume ethanol and can achieve pharmacologically relevant internal ethanol levels, (2) their consumption is not dependent on caloric or sensory properties of ethanol, and (3) they exhibit a relapse-like effect (Devineni and Heberlein 2009). The conditioned preference assay further demonstrates that flies find ethanol intoxication rewarding (Kaun et al. 2011). In both assays flies were willing to overcome negative stimuli (bitter-tasting compound or electric shock, respectively) in order to obtain ethanol or the ethanol-associated cue, suggesting compulsive-like behavior toward ethanol.
However, certain important criteria for addiction have not yet been met in flies. For example, it has not been shown that voluntary ethanol consumption leads to ethanol tolerance, that ethanol removal causes withdrawal symptoms, or that flies are willing to “work” in order to obtain ethanol. The latter criterion could be demonstrated by showing that flies exhibit operant responding for ethanol, a paradigm that has not yet been developed in Drosophila.
In addition to developing new and more complex behavioral assays, many questions remain to be tested using the existing assays for ethanol consumption and reward. For example, flies are typically trained in the conditioned preference assay for only 1 day; it is unknown whether this preference would be altered if flies were trained intermittently over many days. Preliminary data suggest that such long-term ethanol exposures are still perceived as rewarding (K. Kaun and U. Heberlein, unpublished data). In addition, it is unknown whether flies in the conditioned preference assay show behavior indicative of a relapse-like effect, which could be demonstrated by showing that flies exhibit reinstatement to the ethanol-associated cue following extinction.
While more complex assays improve the validity of Drosophila as a model system to study drug addiction, these assays are necessarily more cumbersome and time-consuming, thus making large-scale genetic screening difficult. It is therefore important to understand how the simple and more complex assays for drug-induced behavior are related. Evidence in humans and rodent models is generally consistent with the notion that resistance to the acute effects of ethanol predicts increased ethanol consumption and risk of abuse (Schuckit 1994; Schuckit and Smith 1996; Morean and Corbin 2010). While this association suggests that an overlapping set of genes regulates both simple and complex drug behaviors, several exceptions to this correlation exist (e.g. Colombo et al. 2000; Phillips et al. 1998; Boehm et al. 2004), indicating that factors other than acute drug sensitivity modulate drug intake.
A recent study analyzed the relationships between initial ethanol sensitivity, ethanol tolerance, and voluntary ethanol consumption in flies. Ethanol consumption was positively correlated with the development of tolerance, but not with naive sensitivity to the sedating or hyperactivating effects of ethanol (Devineni et al. 2011). These results suggest that complex behaviors such as voluntary ethanol consumption are not simply readouts of acute responses; they likely incorporate acute sensitivity and the development of tolerance in addition to other factors, such as experience and learning. Some genes have been found to regulate multiple ethanol-induced behaviors: for example kra regulates sedation sensitivity, tolerance, and voluntary ethanol consumption (Berger et al. 2008; Devineni and Heberlein 2009). In contrast, other genes regulate individual ethanol-induced behaviors: for instance sca mediates conditioned ethanol preference, but not acute ethanol sensitivity (Kaun et al. 2011; LaFerriere et al. 2008). Thus, while the simpler assays are ideal for rapidly identifying ethanol-related genes and have some predictive value for more complex behaviors, the development of high-throughput versions of the more complex assays will be necessary in order to apply the power of Drosophila genetics to the study of addiction-like behaviors.
Intriguingly, forward genetic screens in flies have identified many different and unexpected molecular mechanisms that regulate ethanol-related behavior. These mechanisms include signaling pathways such as the cAMP and PI3K pathways that have broad roles in regulating many diverse processes such as development, cell signaling, and neuronal plasticity. Many of the signaling pathways that have been identified can affect each other in complex ways, so it is likely that some pathways may have a more direct role in regulating ethanol responses than others. In most cases it is unknown whether the molecules that have been implicated are direct targets of ethanol or are required downstream of ethanol binding.
One theme that emerges from this large number of pathways is the involvement of molecules affecting molecular or cellular plasticity (e.g. cytoskeletal regulators, ion channels, synaptic molecules), which appear to be recruited to induce behavioral changes in response to ethanol. Much work remains to be done in uncovering how different mechanisms interact to regulate ethanol-related behavior, and how the molecular and cellular changes induced by acute ethanol exposure are translated into addiction-like responses. For example, cytoskeletal changes caused by acute ethanol exposure may be required to induce changes in synapse number, which consequently may mediate ethanol tolerance and preference behaviors. Understanding the relationship between mechanisms mediating acute and long-term responses to ethanol is key to understanding the addictive properties of the drug. The fly is ideally suited to this task due to the availability of tools to investigate these mechanisms with high spatial and temporal resolution.
The next decade should witness the discovery of many novel mechanisms underlying addiction-related behaviors in flies as the number of tools available to study molecular and neural processes is expanding at a rapid rate. Based on what we have learned in the last 15 years from Drosophila addiction research, we expect that these novel mechanisms will be relevant to mammalian models and provide novel targets for the development of pharmacotherapies for drug addiction.
References
Abarca C, Albrecht U, Spanagel R (2002) Cocaine sensitization and reward are under the influence of circadian genes and rhythm. Proc Natl Acad Sci USA 99:9026–9030
Ables JL, Breunig JJ, Eisch AJ, Rakic P (2011) Not(ch) just development: Notch signalling in the adult brain. Nat Rev Neurosci 12:269–283
Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, Amanatides PG, Scherer SE, Li PW, Hoskins RA, Galle RF et al (2000) The genome sequence of Drosophila melanogaster. Science 287:2185–2195
American Psychiatric Association (1994) Diagnostic and statistical manual of mental disorders, 4th edn. Washington DC
Andretic R, Chaney S, Hirsh J (1999) Requirement of circadian genes for cocaine sensitization in Drosophila. Science 285:1066–1068
Andretic R, van Swinderen B, Greenspan RJ (2005) Dopaminergic modulation of arousal in Drosophila. Curr Biol 15:1165–1175
Atkinson NS (2009) Tolerance in Drosophila. J Neurogenet 23:293–302
Bainton RJ, Tsai LT, Singh CM, Moore MS, Neckameyer WS, Heberlein U (2000) Dopamine modulates acute responses to cocaine, nicotine and ethanol in Drosophila. Curr Biol 10:187–194
Bainton RJ, Tsai LT, Schwabe T, DeSalva M, Gaul U, Heberlein U (2005) moody encodes two GPCRs that regulate cocaine behaviors and blood–brain barrier permeability in Drosophila. Cell 123:145–156
Baker NE, Mlodzik M, Rubin GM (1990) Spacing differentiation in the developing Drosophila eye: a fibrinogen-related lateral inhibitor encoded by scabrous. Science 250:1370–1377
Bellen HJ, Levis RW, Liao G, He Y, Carlson JW, Tsang G, Evans-Holm M, Hiesinger PR, Schulze KL, Rubin GM, Hoskins RA, Spradling AC (2004) The BDGP gene disruption project: single transposon insertions associated with 40% of Drosophila genes. Genetics 167:761–781
Berger KH, Heberlein U, Moore MS (2004) Rapid and chronic: two distinct forms of ethanol tolerance in Drosophila. Alcohol Clin Exp Res 28:1469–1480
Berger KH, Kong EC, Dubnau J, Tully T, Moore MS, Heberlein U (2008) Ethanol sensitivity and tolerance in long-term memory mutants of Drosophila melanogaster. Alcohol Clin Exp Res 32:895–908
Bhandari P, Kendler KS, Bettinger JC, Davies AG, Grotewiel M (2009) An assay for evoked locomotor behavior in Drosophila reveals a role for integrins in ethanol sensitivity and rapid ethanol tolerance. Alcohol Clin Exp Res 33:1794–1805
Bingham PM, Lewis R, Rubin GM (1981) Cloning of DNA sequences from the white locus of D. melanogaster by a novel and general method. Cell 25:693–704
Bishop GB, Cullinan WE, Curran E, Gutstein HB (2002) Abused drugs modulate RGS4 mRNA levels in the rat brain: comparison between acute drug treatment and a drug challenge after chronic treatment. Neurobiol Dis 10:334–343
Boehm SL, Peden L, Jennings AW, Kojima N, Harris RA, Blednov YA (2004) Overexpression of the fyn-kinase gene reduces hypnotic sensitivity to ethanol in mice. Neurosci Lett 372:6–11
Brand AH, Perrimon N (1993) Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118:401–415
Brown JM, Hanson GR, Fleckenstein AE (2001) Regulation of the vesicular monoamine transporter-2: a novel mechanism for cocaine and other psychostimulants. J Pharmacol Exp Ther 296:726–727
Cesca F, Baldelli P, Valtorta F, Benfenati F (2010) The synapsins: key actors of synapse function and plasticity. Prog Neurobiol 91:313–348
Chang HY, Grygoruk A, Brooks ES, Ackerson LC, Maidment NT, Bainton RJ, Krantz DE (2006) Overexpression of the Drosophila vesicular monoamine transporter increases motor activity and courtship but decreases the behavioral response to cocaine. Mol Psychiatry 11:99–113
Chen S, Lee AY, Bowens NM, Huber R, Kravitz EA (2002) Fighting fruit flies: a model system for the study of aggression. Proc Natl Acad Sci USA 99:5664–5668
Chen H, Bai J, Ye J, Liu Z, Chen R, Mao W, Li A, Zhou J (2007) JWA as a functional molecule to regulate cancer cells migration via MAPK cascades and F-actin cytoskeleton. Cell Signal 19:1315–1327
Cicero TJ (1979) Biochemistry and Pharmacology of Ethanol, vol 2. Plenum Press, New York
Cohan FM, Graf J-D (1985) Latitudinal cline in Drosophila melanogaster for knockdown resistance to ethanol fumes and for rates of response to selection for further resistance. Evolution 39:278–293
Colombo G, Agabio R, Carai MA, Lobina C, Pani M, Reali R et al (2000) Different sensitivity to ethanol in alcohol preferring sP and non-preferring sNP rats. Alcohol Clin Exp Res 24:1602–1608
Corl AB, Rodan AR, Heberlein U (2005) Insulin signaling in the nervous system regulates ethanol intoxication in Drosophila melanogaster. Nat Neurosci 8:18–19
Corl AB, Berger KH, Ophir-Shohat G, Gesch J, Simms JA, Bartlett S, Heberlein U (2009) Happyhour, a Ste20 family kinase, implicates EGFR signaling in ethanol-induced behaviors. Cell 137:949–960
Cowmeadow RB, Krishnan HR, Atkinson NS (2005) The slowpoke gene is necessary for rapid ethanol tolerance in Drosophila. Alcohol Clin Exp Res 29:1777–1786
Cowmeadow RB, Krishnan HR, Ghezzi A, Al’Hasan YM, Wang YZ, Atkinson NS (2006) Ethanol tolerance caused by slowpoke induction in Drosophila. Alcohol Clin Exp Res 30:745–753
Dasari S, Viele K, Turner AC, Cooper RL (2007) Influence of PCPA and MDMA (ecstasy) on physiology, development and behavior in Drosophila melanogaster. Eur J Neurosci 26:424–438
Davis R (2011) Traces of Drosophila memory. Neuron 70:8–19
Devineni AV, Heberlein U (2009) Preferential ethanol consumption in Drosophila models features of addiction. Curr Biol 19:2126–2132
Devineni AV, McClure K, Guarnieri D, Corl A, Wolf F, Eddison M, Heberlein U (2011) The genetic relationships between ethanol preference, acute ethanol sensitivity and ethanol tolerance in Drosophila melanogaster. Fly 5:191–199
Dietzl G, Chen D, Schnorrer F, Su KC, Barinova Y, Fellner M, Gasser B, Kinsey K, Oppel S, Scheiblauer S, Couto A, Marra V, Keleman K, Dickson BJ (2007) A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature 448:151–156
Dimitrijevic N, Dzitoyeva S, Manev H (2004) An automated assay of the behavioral effects of cocaine injections in adult Drosophila. J Neurosci Methods 137:181–184
Donlea JM, Ramanan N, Shaw PJ (2009) Use-dependent plasticity in clock neurons regulates sleep need in Drosophila. Science 324:105–108
Dudley R (2002) Fermenting fruit and the historical ecology of ethanol ingestion: is alcoholism in modern humans an evolutionary hangover? Addiction 97:381–388
Dzitoyeva S, Dimitrijevic N, Manev H (2003) γ-Aminobutyric acid B receptor 1 mediates behavior-impairing actions of alcohol in Drosophila: adult RNA interference and pharmacological evidence. Proc Natl Acad Sci USA 100:5485–5490
Eddison M, Guarnieri DJ, Cheng L, Liu C-H, Moffat KG, Davis G, Heberlein U (2011) arouser reveals a role for synapse number in the regulation of ethanol sensitivity. Neuron 70:979–990
El Ghundi M, George SR, Drago J, Fletcher PJ, Fan T, Nguyen T, Liu C, Sibley DR, Westphal H, O’Dowd BF (1998) Disruption of dopamine D1 receptor gene expression attenuates alcohol-seeking behavior. Eur J Pharmacol 353:149–158
Friedhoff AJ, Miller J (1973) Effect of ethanol on biosynthesis of dopamine. Ann N Y Acad Sci 215:183–186
Ge X, Hannan F, Xie Z, Feng C, Tully T, Zhou H, Xie Z, Zhong Y (2004) Notch signaling in Drosophila long-term memory formation. Proc Natl Acad Sci USA 101:10172–10176
Geer BW, Heinstra PW, McKechnie SW (1993) The biological basis of ethanol tolerance in Drosophila. Comp Biochem Physiol B 105:203–229
George R, Lease K, Burnette J, Hirsh J (2005) A “bottom-counting” video system for measuring cocaine-induced behaviors in Drosophila. Methods Enzymol 393:841–851
Giros B, Jaber M, Jones ST, Wightman RM, Caron MG (1996) Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature 379:606–612
Godenschwege TA, Reisch D, Diegelmann S, Eberle K, Funk N, Heisenberg M, Hoppe V, Hoppe J, Klagges BRE, Martin J-R, Nikitina EA, Putz G, Reifegerste R, Reisch N, Rister J, Schaupp M, Scholz H, Schwarzel M, Werner U, Zars TD, Buchner S, Buchner E (2004) Flies lacking all synapsins are unexpectedly healthy but are impaired in complex behaviour. Eur J Neurosci 20:611–622
Gold SJ, Han MH, Herman AE, Ni YG, Pudiak CM, Aghajanian GK, Liu RJ, Potts BW, Mumby SM, Nestler EJ (2003) Regulation of RGS proteins by chronic morphine in rat locus coeruleus. Eur J Neurosci 17:971–980
Granderath S, Stollework A, Greig S, Goodman CS, O’Kane CJ, Klambt C (1999) loco encodes an RGS protein required for Drosophila glial differentiation. Development 126:1781–1791
Greenspan RJ, Ferveur J-F (2000) Courtship in Drosophila. Annu Rev Genet 34:205–232
Hamada FN, Rosenzweig M, Kang K, Pulver SR, Ghezzi A, Jegla TJ, Garrity PA (2008) An internal thermal sensor controlling temperature preference in Drosophila. Nature 454:217–220
Heberlein U, Tsai LT, Kapfhamer D, Lasek AW (2009) Drosophila, a genetic model system to study cocaine-related behaviors: a review with focus on LIM-only proteins. Neuropharmacology 56(Suppl 1):97–106
Hodge CW, Mehmert KK, Kelley SP, McMahon T, Haywood A, Olive MF, Wang D, Sanchez-Perez AM, Messing RO (1999) Supersensitivity to allosteric GABA(A) receptor modulators and alcohol in mice lacking PKCepsilon. Nat Neurosci 2:997–1002
Hoffmann AA, Parsons PA (1984) Olfactory response and resource utilization in Drosophila—interspecific comparison. Biol J Linnean Soc 22:43–53
Holland PW (2003) More genes in vertebrates? J Struct Funct Genomics 3:75–84
Ja WW, Carvalho GB, Mak EM, de la Rosa NN, Fang AY, Liong JC, Brummel T, Benzer S (2007) Prandiology of Drosophila and the CAFÉ assay. Proc Natl Acad Sci USA 104:8253–8256
Kaun KR, Azanchi R, Maung Z, Hirsh J, Heberlein U (2011) A Drosophila model for ethanol reward. Nat Neurosci 14:612–619
King I, Tsai LT, Pflanz R, Voigt A, Lee S, Jackle H, Lu B, Heberlein U (2011) Drosophila tao controls mushroom body development and ethanol-stimulated behavior through par-1. J Neurosci 31:1139–1148
Kitamoto T (2001) Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. J Neurobiol 47:81–92
Knott TK, Dopico AM, Dayanithi G, Lemos J, Treistman SN (2002) Integrated channel plasticity contributes to alcohol tolerance in neurohypophysial terminals. Mol Pharmacol 62:135–142
Kong EC, Allouche L, Chapot PA, Vranizan K, Moore MS, Heberlein U, Wolf FW (2010a) Ethanol-regulated genes that contribute to ethanol sensitivity and rapid tolerance in Drosophila. Alcohol Clin Exp Res 34:302–316
Kong EC, Woo K, Haiyan L, Lebestky T, Mayer N, Sniffen MR, Heberlein U, Bainton RJ, Hirsh J, Wolf FW (2010b) A pair of dopamine neurons target the D1-like dopamine receptor DopR in the central complex to promote ethanol-stimulated locomotion in Drosophila. PLoS ONE 5:e9954
Koob GF (2004) A role for GABA mechanisms in the motivational effects of alcohol. Biochem Pharmacol 68:1515–1525
Koob GF, Nestler EJ (1997) The neurobiology of drug addiction. J Neurophychiatry Clin Neurosci 9:482–497
Krashes MJ, Keene AC, Leung B, Armstrong JD, Waddell S (2007) Sequential use of mushroom body neuron subsets during Drosophila odor memory processing. Neuron 53:103–115
Krishnan HR, Al-Hasan YM, Pohl JB, Ghezzi A, Atkinson NS (2011) A role for dynamin in triggering ethanol tolerance. Alcohol Clin Exp Res. doi:10.1111/j.1530-0277.2011.01587.x [Epub ahead of print]
Kurtz DL, Stewart RB, Zweifel M, Li T-K, Froehlich JC (1996) Genetic differences in tolerance and sensitization to the sedative/hypnotic effects of alcohol. Pharmacol Biochem Behav 53:585–591
LaFerriere H, Guarnieri DJ, Sitaraman D, Diegelmann D, Heberlein U, Zars T (2008) Genetic dissociation of ethanol sensitivity and memory formation in Drosophila melanogaster. Genetics 178:1895–1902
Lai SL, Lee T (2006) Genetic mosaic with dual binary transcriptional systems in Drosophila. Nat Neurosci 9:703–709
Lasek AW, Kapfhamer D, Kharazia V, Gesch J, Giorgetti F, Heberlein U (2010) Lmo4 in the nucleus accumbens regulates cocaine sensitivity. Genes Brain Behav 9:817–824
Lasek AW, Giorgetti F, Berger KH, Tayor S, Heberlein U (2011a) Lmo genes regulate behavioral responses to ethanol in Drosophila melanogaster and the mouse. Alcohol Clin Exp Res 35:1600–1606
Lasek AW, Lim J, Kliethermes CL, Berger KH, Joslyn G, Brush G, Xue L, Robertson M, Moore MS, Vranizan K, Morris SW, Schuckit MA, White RL, Heberlein U (2011b) An evolutionary conserved role for anaplastic lymphoma kinase in behavioral responses to ethanol. PLoS ONE 6:e22636
Lee S, Nahm M, Lee M, Kwon M, Kim E, Zadeh AS, Cao H, Kim HJ, Lee ZH, Oh SB, Kolodziej PA, Lee S (2007) The F-actin-microtubule crosslinker Shot is a platform for Krasavietz-mediated translational regulation of midline axon repulsion. Development 134:1767–1777
Li H, Chaney S, Roberts IJ, Forte M, Hirsh J (2000) Ectopic G-protein expression in dopamine and serotonin neurons blocks cocaine sensitization in Drosophila melanogaster. Curr Biol 10:211–214
Li C, Zhao X, Xingjiang C, Chu D, Chen J, Zhou J (2008) The Drosophila homolog of jwa is required for ethanol tolerance. Alcohol Alcohol 43:529–536
Littleton JT, Ganetzky B (2000) Ion channels and synaptic organization: analysis of the Drosophila genome. Neuron 26:35–43
Lloyd TE, Verstreken P, Ostrin EJ, Phillippi A, Lichtarge O et al (2000) A genome-wide search for synaptic vesicle proteins in Drosophila. Neuron 26:45–50
Luan H, Peabody NC, Vinson CR, White BJ (2006) Refining spatial manipulation of neuronal function by combinatorial restriction of transgene expression. Neuron 52:425–436
Martin JR, Raabe T, Heisenberg M (1999) Central complex structures are required for the maintenance of locomotor activity in Drosophila melanogaster. J Comp Physiol A 185:277–288
Matenia D, Mandelkow EM (2009) The tau of MARK: a polarized view of the cytoskeleton. Trends Biochem Sci 34:332–342
McBride WJ, Li TK (1998) Animal models of alcoholism: neurobiology of high alcohol-drinking behavior in rodents. Crit Rev Neurobiol 12:339–369
McClung C, Hirsh J (1998) Stereotypic behavioral responses to free-base cocaine and the development of behavioral sensitization in Drosophila. Curr Biol 8:109–112
McClung CA, Sidiropoulou K, Vitaterna M, Takahashi JS, White FJ, Cooper DC, Nesstler EJ (2005) Regulation of dopaminergic transmission and cocaine reward by the Clock gene. Proc Natl Acad Sci USA 102:9377–9381
McGuire SE, Le PT, Osborn AJ, Matsumoto K, Davis RL (2003) Spatiotemporal rescue of memory dysfunction in Drosophila. Science 302:1765–1768
McKenzie JA, Parsons P (1972) Alcohol tolerance—ecological parameter in relative success of Drosophila melanogaster and Drosphila simulans. Oecologia 10:373–388
Mezler M, Muller T, Raming K (2001) Cloning and functional expression of GABAB receptors from Drosophila. Eur J Neurosci 13:477–486
Moore MS, DeZazzo J, Luk AY, Tully T, Singh CM, Heberlein U (1998) Ethanol intoxication in Drosophila: genetic and pharmacological evidence for regulation by the cAMP signaling pathway. Cell 93:997–1007
Morean ME, Corbin WR (2010) Subjective response to alcohol: a critical review of the literature. Alcohol Clin Exp Res 34:385–395
Morozova TV, Anholt RR, Mackay TF (2006) Transcriptional response to alcohol exposure in Drosophila melanogaster. Genome Biol 7:R95
Morozova TV, Anholt RR, Mackay TF (2007) Phenotypic and transcriptional response to selection for alcohol sensitivity in Drosophila melanogaster. Genome Biol 8:R231
Nassel DR, Elekes K (1992) Aminergic neurons in the brain of blowflies and Drosophila: dopamine- and tyrosine hydroxylase-immunoreactive neurons and their relationship with putative histaminergic neurons. Cell Tissue Res 267:147–167
Nestler EJ (2005) Is there a common molecular pathway for addiction? Nat Neurosci 8:1445–1449
Neuser K, Triphan T, Mronz M, Poeck B, Strauss R (2008) Analysis of a spatial orientation memory in Drosophila. Nature 453:1244–1247
Ofstad TA, Zuker CS, Reiser MB (2011) Visual place learning in Drosophila melanogaster. Nature 474:204–207
Park SK, Sedore SA, Cronmiller C, Hirsh J (2000) Type II cAMP-dependent protein kinase-deficient Drosophila are viable but show developmental, circadian, and drug response phenotypes. J Biol Chem 275:20588–20596
Pfeiffer BD, Jenett A, Hammonds AS, Ngo TT, Misra S, Murphy C, Scully A, Carlson JW, Wan KH, Laverty TR, Mungall C, Svirskas R, Kadonaga JT, Doe CQ, Eisen MB, Celniker SE, Rubin GM (2008) Tools for neuroanatomy and neurogenetics in Drosophila. Proc Natl Acad Sci USA 105:9715–9720
Phillips TJ, Brown KJ, Burkhard-Kasch S, Wenger CD, Kelly MS, Rubinstein M et al (1998) Alcohol preference and sensitivity are markedly reduced in mice lacking dopamine D2 receptors. Nat Neurosci 1:610–615
Pick S, Strauss R (2005) Goal-driven behavioral adaptations in gap-climbing Drosophila. Curr Biol 15:1473–1478
Piper PW (1995) The heat shock and ethanol stress responses of yeast exhibit extensive similarity and functional overlap. FEMS Microbiol Lett 134:121–127
Powell PA, Wesley C, Spencer S, Cagan RL (2001) Scabrous complexes with Notch to mediate boundary formation. Nature 409:626–630
Pradhan S (1983) Effect of cocaine on rat brain enzymes. Arch Int Pharmacodyn Ther 266:221–228
Presente A, Boyles RS, Serway CN, deBelle S, Andres A (2004) Notch is required for long-term memory in Drosophila. Proc Natl Acad Sci USA 101:1746–1768
Pulver SR, Pashkovski SL, Hornstein NJ, Garrity PA, Griffith LC (2009) Temporal dynamics of neuronal activation by Channelrhodopsin-2 and TRPA1 determine behavioral output in Drosophila larvae. J Neurophysiol 101:3075–3088
Quinn WG, Harris WA, Benzer S (1974) Conditioned behavior in Drosophila melanogaster. Proc Natl Acad Sci USA 71:708–712
Reiter LT, Potocki L, Chien S, Gribskov M, Bier E (2001) A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res 11:1114–1125
Renn SC, Park JH, Rosbash M, Hall JC, Taghert PH (1999) A pdf neuropeptide gene mutation and ablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms in Drosophila. Cell 99:791–802
Riley BP, Kalsi G, Kuo PH, Vladimirov V, Thiselton DL, Vittum J, Wormley B, Grotewiel MS, Patterson DG, Sullivan PF, van den Oord E, Walsh D, Kendler KS, Prescott CA (2006) Alcohol dependence is associated with the ZNF699 gene, a human locus related to Drosophila hangover, in the Irish Affected Sib Pair Study of Alcohol Dependence (IASPSAD) sample. Mol Psychiatry 11:1025–1031
Rothenfluh A, Threlkeld RJ, Bainton RJ, Tsai LT, Lasek AW, Heberlein U (2006) Distinct behavioral responses to ethanol are regulated by alternate RhoGAP18B isoforms. Cell 127:199–211
Rubin GM, Spradling AC (1982) Genetic transformation of Drosophila with transposable element vectors. Science 218:348–353
Sanchez-Soriano N, Travis M, Dajas-Bailador F, Goncalves-Pimental C, Whitmarsh AJ, Prokop A (2009) Mouse ACF7 and drosophila short stop modulate filopodia formation and microtubule organization during neuronal growth. J Cell Sci 122:2534–2542
Satel SL, Southwick SM, Gawin FH (1991) Clinical features of cocaine-induced paranoia. Am J Psychiatry 148:495–498
Scholz H, Ramond J, Singh CM, Heberlein U (2000) Functional ethanol tolerance in Drosophila. Neuron 28:261–271
Scholz H, Franz M, Heberlein U (2005) The hangover gene defines a stress pathway required for ethanol tolerance development. Nature 436:845–847
Schuckit MA (1994) Low level of response to alcohol as a predictor of future alcoholism. Am J Psychiatry 151:184–189
Schuckit MA, Smith TL (1996) An 8-year follow-up of 450 sons of alcoholic and control subjects. Arch Gen Psychiatry 53:202–210
Schuckit MA, Smith TL, Hesselbrock V, Bucholz KK, Bierut L, Edenberg H, Kramer J, Longacre E, Fukura T, Kalmign J, Danko GP, Trim R (2008) Clinical implications of tolerance to alcohol in nondependent young drinkers. Am J Drug Alcohol Abuse 34:133–149
Schuldiner O, Berdnik D, Levy JM, Wu JS, Luginbuhl D, Gontang AC, Luo L (2008) piggyBac-based mosaic screen identifies a postmitotic function for cohesin in regulating developmental axon pruning. Dev Cell 14:227–238
Schumann G, Coin LJ, Lourdusamy A, Charoen P, Berger KH et al (2011) Genome-wide association and genetic functional studies identify autism susceptibility candidate 2 gene (AUTS2) in the regulation of alcohol consumption. Proc Natl Acad Sci 108:7119–7124
Schwabe T, Bainton RJ, Fetter RD, Heberlein U, Gaul U (2005) GPCR signaling is required for blood-brain barrier formation in Drosophila. Cell 123:133–134
Schwendt M, Gold SJ, McGinty JF (2006) Acute amphetamine down-regulates RGS4 mRNA and protein expression in rat forebrain: distinct roles of D1 and D2 dopamine receptors. J Neurochem 96:1606–1615
Schwendt M, Hearing MC, See RE, McGinty JF (2007) Chronic cocaine reduced RGS4 mRNA in rat prefrontal cortex and dorsal striatum. NeuroReport 18:1261–1265
Schwenkert I, Eltrop R, Funk N, Steinert JR, Schuster CM, Scholz H (2008) The hangover gene negatively regulates bouton addition at the Drosophila neuromuscular junction. Mech Dev 125:700–711
Siggins GR, Roberto M, Nie Z (2005) The tipsy terminal: presynaptic effects of ethanol. Pharmacol Ther 107:80–98
Soderpalm B, Lof E, Ericson M (2009) Mechanistic studies of ethanol’s interaction with the mesolimbic dopamine reward system. Pharmacopsychiatry 42:S87–S94
Szumlinski KK, Lominac KD, Oleson EB, Walker JK, Mason A, Dehoff MH, Klugmann M, Cagle S, Welt K, During M, Worley PF, Middaugh LD, Kalivas PW (2005) Homer2 is necessary for EtOH-induced neuroplasticity. J Neurosci 25:7054–7061
Tabakoff B, Ritzmann RF (1977) The effects of 6-hydroxydopamine on tolerance and dependence on ethanol. J Pharmacol Exp Ther 203:319–331
Thibault ST, Singer MA, Miyazaki WY, Milash B, Dompe NA, Singh CM, Buchholz R, Demsky M, Fawcett R, Francis-Lang HL, Ryner L, Cheung LM, Chong A, Erickson C, Fisher WW, Greer K, Hartouni SR, Howie E, Jakkula L, Joo D, Killpack K, Laufer A, Mazzotta J, Smith RD, Stevens LM, Stuber C, Tan LR, Ventura R, Woo A, Zakrajsek I, Zhao L, Chen F, Swimmer C, Kopczynski C, Duyk G, Winberg ML, Margolis J (2004) A complementary transposon tool kit for Drosophila melanogaster using P and piggyBac. Nat Genet 36:283–287
Thiele TE, Marsh DJ, Ste Marie L, Bernstein IL, Palmiter RD (1998) Ethanol consumption and resistance are inversely related to neuropeptide Y levels. Nature 396:366–369
Torres G, Horowitz JM (1998) Activating properties of coaine and cocaethylene in a behavioral preparation of Drosophila melanogaster. Synapse 29:148–161
Tsai LT, Bainton RJ, Blau J, Heberlein U (2004) Lmo mutants reveal a novel role for circadian pacemaker neurons in cocaine-induced behaviors. PLoS Biol 2:e408
Urizar NL, Yang Z, Edenberg HJ, Davis RL (2007) Drosophila Homer is required in a small set of neurons including the ellipsoid body for normal ethanol sensitivity and tolerance. J Neurosci 27:4541–4551
Wand G, Levine M, Zweifel L, Schwindinger W, Abel T (2001) The cAMP–protein kinase A signal transduction pathway modulates ethanol consumption and sedative effects of ethanol. J Neurosci 21:5297–5303
Weber KE (1988) An apparatus for measurement of resistance to gas-phase reagents. Drosophila Inform Serv 67:90–92
Wen T, Parrish CA, Xu D, Wu Q, Shen P (2005) Drosophila neuropeptide F and its receptor, NPFR1, define a signaling pathway that acutely modulates alcohol sensitivity. Proc Natl Acad Sci USA 102:2141–2146
Wilke N, Sganga M, Barhite S, Miles MF (1994) Effects of alcohol on gene expression in neural cells. EXS 71:49–59
Wolf FW, Rodan AR, Tsai LT-Y, Heberlein U (2002) High-resolution analysis of ethanol-induced locomotor stimulation in Drosophila. J Neurosci 22:11035–11044
Wu D, Cederbaum AI (2009) Oxidative stress and alcoholic liver disease. Semin Liver Dis 29:141–154
Wu CL, Xia S, Fu TF, Wang H, Chen YH, Leong D, Chiang AS, Tully T (2007) Specific requirement of NMDA receptors for long-term memory consolidation in Drosophila ellipsoid body. Nat Neurosci 10:1578–1586
Yellman C, Tao H, He B, Hirsh J (1998) Conserved and sexually dimorphic behavioral responses to biogenic amines in decapitated Drosophila. Proc Natl Acad Sci USA 94:4131–4136
Zaleski MJ, Nunes Filho JR, Lemos T, Morato GS (2001) GABA(B) receptors play a role in the development of tolerance to ethanol in mice. Psychopharmacology 153:415–424
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
K.R. Kaun and A.V. Devineni contributed equally to this work.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Kaun, K.R., Devineni, A.V. & Heberlein, U. Drosophila melanogaster as a model to study drug addiction. Hum Genet 131, 959–975 (2012). https://doi.org/10.1007/s00439-012-1146-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00439-012-1146-6