
Abstract
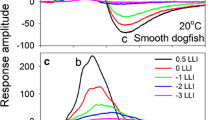
Sharks are apex predators, and their evolutionary success is in part due to an impressive array of sensory systems, including vision. The eyes of sharks are well developed and function over a wide range of light levels. However, whilst close relatives of the sharks—the rays and chimaeras—are known to have the potential for colour vision, an evolutionary trait thought to provide distinct survival advantages, evidence for colour vision in sharks remains equivocal. Using single-receptor microspectrophotometry, we measured the absorbance spectra of visual pigments located in the retinal photoreceptors of 17 species of shark. We show that, while the spectral tuning of the rod (wavelength of maximum absorbance, λmax 484–518 nm) and cone (λmax 532–561 nm) visual pigments varies between species, each shark has only a single long-wavelength-sensitive cone type. This suggests that sharks may be cone monochromats and, therefore, potentially colour blind. Whilst cone monochromacy on land is rare, it may be a common strategy in the marine environment: many aquatic mammals (whales, dolphins and seals) also possess only a single, green-sensitive cone type. It appears that both sharks and marine mammals may have arrived at the same visual design by convergent evolution. The spectral tuning of the rod and cone pigments of sharks is also discussed in relation to their visual ecology.

Similar content being viewed by others

References
Alpern M, Lee GB, Maaseidvaag F, Miller SS (1971) Colour vision in blue-cone ‘monochromacy’. J Physiol 212(1):211–233
Anderson WG, Good JP, Pillans RD, Hazon N, Franklin CE (2005) Hepatic urea biosynthesis in the euryhaline elasmobranch Carcharhinus leucas. J Exp Zool A 303(10):917–921. doi:10.1002/jez.a.199
Bowmaker JK, Govardovskii VI, Shukolyukov SA, Zueva LV, Hunt DM, Sideleva VG, Smirnova OG (1994) Visual pigments and the photic environment: the cottoid fish of Lake Baikal. Vis Res 34(5):591–605. doi:10.1016/0042-6989(94)90015-9
Busch H, Dücker G (1987) Das visuelle leistungsvermögen der Seebären (Arctocephalus pusillus und Arctocephalus australis). Zool Anz 219(3–4):197–224
Chidlow JA (2003) The biology of wobbegong sharks (Family: Orectolobidae) from south-western Australian waters. MSc thesis, James Cook University, Townsville
Clark E (1963) The maintenance of sharks in captivity with a report on their instrumental conditioning. In: Gilbert PW (ed) Sharks and survival. DC Heath, Boston, pp 115–149
Cohen JL (1980) Functional organization of the retina of the lemon shark (Negaprion brevirostris, Poey): an anatomical and electrophysiological approach. PhD thesis, University of Miami, Florida
Cohen J (1988) Statistical power analysis for the behavioral sciences. Lawrence Erlbaum Associates, New Jersey
Cohen JL, Gruber SH (1985) Spectral input to lemon shark (Negaprion brevirostris) ganglion cells. J Comp Physiol A 156:579–586. doi:10.1007/BF00619108
Cohen JL, Hueter RE, Organisciak DT (1990) The presence of a porphyropsin-based visual pigment in the juvenile lemon shark (Negaprion brevirostris). Vis Res 30(12):1949–1953. doi:10.1016/0042-6989(90)90014-C
Compagno L, Dando M, Fowler S (2005) A field guide to the sharks of the world. Harper Collins Publishers, Ltd., London
Crognale MA, Levenson D, Pongais PJ, Deegan JF 2nd, Jacobs GH (1998) Cone spectral sensitivity in the harbor seal (Phoca vitulina) and implications for color vision. Can J Zool 76(11):2114–2118
Davies WL, Carvalho LS, Tay BH, Brenner S, Hunt DM, Venkatesh B (2009) Into the blue: gene duplication and loss underlie color vision adaptations in a deep-sea chimaera, the elephant shark Callorhinchus milii. Genome Res 19(3):415–426. doi:10.1101/gr.084509.108
Denton EJ, Shaw TI (1963) The visual pigments of some deep-sea elasmobranchs. J Mar Biol Assoc UK 43:65–70
Denton EJ, Warren FJ (1956) Visual pigments of deep sea fish. Nature 178:1059. doi:10.1038/1781059a0
Douglas RH, Partridge JC, Marshall NJ (1998) The eyes of deep-sea fish. I: Lens pigmentation, tapeta and visual pigments. Prog Ret Eye Res 17(4):597–636. doi:10.1016/S1350-9462(98)00002-0
Dowling JE, Ripps H (1990) On the duplex nature of the skate retina. J Exp Zool 256(Supplement 5):55–65. doi:10.1002/jez.1402560509
Esaias WE, Curl HC, Seliger HH (1973) Action spectrum for a low intensity, rapid photoinhibition of mechanically stimulable bioluminescence in the marine dinoflagellates Gonyaulax catenella, G. acatenella, and G. tamarensis. J Cell Physiol 82(3):363–372. doi:10.1002/jcp.1040820306
Frank TM, Widder EA (1999) Comparative study of the spectral sensitivities of mesopelagic crustaceans. J Comp Physiol A 185(3):255–265. doi:10.1007/s003590050385
Gačić Z, Damjanović I, Mićković B, Hegediš A, Nikčević M (2007) Spectral sensitivity of the dogfish shark (Scyliorhinus canicula). Fish Physiol Biochem 33(1):21–27. doi:10.1007/s10695-006-9113-6
Govardovskii VI, Lychakov LV (1977) Visual cells and visual pigments in the Black Sea elasmobranchs. Zh Evol Biokhim Fiziol 13(2):162–166
Govardovskii VI, Fyhrquist N, Reuter T, Kuzmin DG, Donner K (2000) In search of the visual pigment template. Vis Neurosci 17(4):509–528. doi:10.1017/S0952523800174036
Griebel U, Schmid A (1992) Color vision in the California sea lion (Zalophus californianus). Vision Res 32(3):477–482. doi:10.1016/0042-6989(92)90239-F
Griebel U, Schmid A (2002) Spectral sensitivity and color vision in the bottlenose dolphin (Tursiops Truncatus). Mar Freshw Behav Phy 35(3):129–137. doi:10.1080/1023624021000014716
Gruber SH (1975) Duplex vision in the elasmobranchs: histological, electrophysiological and psychophysical evidence. In: Ali MA (ed) Vision in fishes: new approaches in research. Plenum, New York and London, pp 525–540
Haddock SHD, Moline MA, Case JF (2010) Bioluminescence in the sea. Ann Rev Mar Sci 2(1):443–493. doi:10.1146/annurev-marine-120308-081028
Hart NS (2002) Vision in the peafowl (Aves: Pavo cristatus). J Exp Biol 205(24):3925–3935
Hart NS (2004) Microspectrophotometry of visual pigments and oil droplets in a marine bird, the wedge-tailed shearwater Puffinus pacificus: topographic variations in photoreceptor spectral characteristics. J Exp Biol 207(7):1229–1240. doi:10.1242/jeb.00857
Hart NS, Partridge JC, Cuthill IC (1998) Visual pigments, oil droplets and cone photoreceptor distribution in the European starling (Sturnus vulgaris). J Exp Biol 201(9):1433–1446
Hart NS, Lisney TJ, Marshall NJ, Collin SP (2004) Multiple cone visual pigments and the potential for trichromatic colour vision in two species of elasmobranch. J Exp Biol 207(26):4587–4594. doi:10.1242/jeb.01314
Hart NS, Lisney TJ, Collin SP (2006) Visual communication in elasmobranchs. In: Kapoor BG, Ladich F, Collin SP, Raschi WG (eds) Fish communication, vol. 2. Science Publishers Inc, Enfield, pp 337–392
Huveneers C, Otway NM, Gibbs SE, Harcourt RG (2007) Quantitative diet assessment of wobbegong sharks (genus Orectolobus) in New South Wales, Australia. ICES J Mar Sci 64(6):1272–1281. doi:10.1093/icesjms/fsm111
Jacobs GH, Deegan JF 2nd, Neitz J, Crognale MA, Neitz M (1993) Photopigments and color vision in the nocturnal monkey, Aotus. Vis Res 33(13):1773–1783. doi:10.1016/0042-6989(93)90168-V
Jerlov NG (1976) Marine optics. Elsevier Scientific Publishing Company, Amsterdam
Jokela M, Vartio A, Paulin L, Fyhrquist-Vanni N, Donner K (2003) Polymorphism of the rod visual pigment between allopatric populations of the sand goby (Pomatoschistus minutus): a microspectrophotometric study. J Exp Biol 206(15):2611–2617. doi:10.1242/jeb.00472
Kolb H, Famiglietti EV (1974) Rod and cone pathways in the inner plexiform layer of cat retina. Science 186(4158):47–49. doi:10.1126/science.186.4158.47
Last PR, Stevens JD (2009) Sharks and rays of Australia, 2nd edn. CSIRO, Melbourne
Levine JS, MacNichol EF Jr (1982) Color vision in fishes. Sci Am 246(2):108–117
Levine JS, MacNichol EF Jr (1985) Microspectrophotometry of primate photoreceptors: art, artefact and analysis. In: Fein A, Levine JS (eds) The visual system. Liss, New York, pp 73–87
Lisney TJ, Collin SP (2006) Brain morphology in large pelagic fishes: a comparison between sharks and teleosts. J Fish Biol 68(2):532–554. doi:10.1111/j.0022-1112.2006.00940.x
Loew ER, Lythgoe JN (1978) The ecology of cone pigments in teleost fishes. Vis Res 18(6):715–722. doi:10.1016/0042-6989(78)90150-5
Lythgoe JN, Partridge JC (1991) The modelling of optimal visual pigments of dichromatic teleosts in green coastal waters. Vis Res 31(3):361–371. doi:10.1016/0042-6989(91)90089-N
Lythgoe JN, Muntz WRA, Partridge JC, Shand J, Williams DM (1994) The ecology of the visual pigments of snappers (Lutjanidae) on the Great Barrier Reef. J Comp Physiol A 174(4):461–467. doi:10.1007/BF00191712
MacNichol EF Jr (1986) A unifying presentation of photopigment spectra. Vis Res 26(9):1543–1556. doi:10.1016/0042-6989(86)90174-4
Madsen C (1976) Tests for color discrimination and spectral sensitivity in the bottlenose dolphin, Tursiops truncatus. PhD thesis, University of Hawaii, Honolulu
McFarland WN, Munz FW (1975) Part II: the photic environment of clear tropical seas during the day. Vis Res 15:1063–1070. doi:10.1016/0042-6989(75)90002-4
Mensinger AF, Case JF (1992) Dinoflagellate luminescence increases susceptibility of zooplankton to teleost predation. Mar Biol 112(2):207–210. doi:10.1007/bf00702463
Munz FW (1957) Photosensitive pigments from retinas of deep-sea fishes. Science 125:1142–1143. doi:10.1126/science.125.3258.1142
Munz FW, McFarland WN (1973) The significance of spectral position in the rhodopsins of tropical marine fishes. Vis Res 13:1829–1874. doi:10.1016/0042-6989(73)90060-6
Newman LA, Robinson PR (2005) Cone visual pigments of aquatic mammals. Vis Neurosci 22(6):873–879. doi:10.1017/S0952523805226159
Niwa H, Tamura T (1975) Investigation of fish vision by means of S-potentials—III. Photoreceptors and spectral sensitivity in elasmobranchs’ retinae. Nippon Suisan Gakkaishi 41(4):393–401
Pang JJ, Abd-El-Barr MM, Gao F, Bramblett DE, Paul DL, Wu SM (2007) Relative contributions of rod and cone bipolar cell inputs to AII amacrine cell light responses in the mouse retina. J Physiol 580(2):397–410. doi:10.1113/jphysiol.2006.120790
Parry JW, Bowmaker JK (2000) Visual pigment reconstitution in intact goldfish retina using synthetic retinaldehyde isomers. Vis Res 40(17):2241–2247. doi:10.1016/S0042-6989(00)00101-2
Peichl L, Behrmann G, Kröger RHH (2001) For whales and seals the ocean is not blue: a visual pigment loss in marine mammals. Eur J Neurosci 13:1520–1528. doi:10.1046/j.0953-816X.2001.01533.x
Powter DM, Gladstone W, Platell M (2010) The influence of sex and maturity on the diet, mouth morphology and dentition of the Port Jackson shark, Heterodontus portusjacksoni. Mar Fresh Res 61(1):74–85. doi:10.1071/MF09021
Ripps H, Dowling JE (1990) Structural features and adaptive properties of photoreceptors in the skate retina. J Exp Zool 265(Supplement 5):46–54. doi:10.1002/jez.1402560508
Scheiber N (2007) Comparative ultrastructure of shark and ray photoreceptors. Honours thesis, The University of Queensland, Brisbane
Scholtyssek C, Kelber A, Dehnhardt G (2008) Brightness discrimination in the harbor seal (Phoca vitulina). Vis Res 48(1):96–103. doi:10.1016/j.visres.2007.10.012
Sillman AJ, Letsinger GA, Patel S, Loew ER, Klimley AP (1996) Visual pigments and photoreceptors in two species of shark, Triakis semifasciata and Mustelus henlei. J Exp Zool 276(1):1–10
Szél Á, Csorba G, Caffé AR, Szél G, Röhlich P, van Veen T (1994) Different patterns of retinal cone topography in two genera of rodents, Mus and Apodemus. Cell Tissue Res 276(1):143–150
Tamura T, Niwa H (1967) Spectral sensitivity and color vision of fish as indicated by S-potential. Comp Biochem Physiol 22:745–754
Temple S, Hart NS, Marshall NJ, Collin SP (2010) A spitting image: specializations in archerfish eyes for vision at the interface between air and water. Proc R Soc Lond B Biol Sci 277(1694):2607–2615. doi:10.1098/rspb.2010.0345
Tester AL, Kato S (1966) Visual target discrimination in blacktip sharks (Carcharhinus melanopterus) and grey sharks (C. menisorrah). Pac Sci 20:461–471
Theiss SM (2009) Sensory biology and ecology of wobbegong sharks (Orectolobidae). PhD thesis, The University of Queensland, Brisbane
Theiss SM, Lisney TJ, Collin SP, Hart NS (2007) Colour vision and visual ecology of the blue-spotted maskray, Dasyatis kuhlii Muller & Henle, 1814. J Comp Physiol A 193(1):67–79. doi:10.1007/s00359-006-0171-0
Toyoda JI, Saito T, Kondo H (1978) Three types of horizontal cells in the stingray retina: their morphology and physiology. J Comp Neurol 179(3):569–579. doi:10.1002/cne.901790308
Wartzok D, McCormick MG (1978) Color discrimination by a Bering Sea spotted seal, Phoca largha. Vis Res 18(7):781–784. doi:10.1016/0042-6989(78)90117-7
Zhao H, Rossiter SJ, Teeling EC, Li C, Cotton JA, Zhang S (2009) The evolution of color vision in nocturnal mammals. Proc Natl Acad Sci USA 106(22):8980–8985. doi:10.1073/pnas.0813201106
Acknowledgements
The authors would like to thank Scott Cutmore, Jeremy Ullmann, Clint Chapman, John Page, Alan Goldizen, Jamie Thornton, Darren Sapelli and Bob Stone for assistance in obtaining specimens, and three anonymous reviewers for their helpful comments on the manuscript. Funding was provided by the Australian Research Council and the Sea World Research and Rescue Foundation. This paper is dedicated to the late Dr. Julia Shand.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hart, N.S., Theiss, S.M., Harahush, B.K. et al. Microspectrophotometric evidence for cone monochromacy in sharks. Naturwissenschaften 98, 193–201 (2011). https://doi.org/10.1007/s00114-010-0758-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00114-010-0758-8