
Abstract
This study analyses the rhodolith-bearing deposits in the largest and most rhodolith-rich outcrop of the Polish Outer Carpathian flysch, located in the Silesian Nappe, at the village of Melsztyn. The rhodoliths and sparse associated biota occur as resedimented components in a deep-marine succession of siliciclastic conglomerates and coarse-grained sandstones, deposited by high-density turbidity currents and debris flows. The sediment was derived from a fan-delta system located at the southern margin of the Silesian flysch basin. Stratigraphic data indicate that the succession represents the Upper Istebna Sandstone deposited during the Late Paleocene. The rhodoliths are composed mostly of coralline red algae with seven genera and eight species representing the family Sporolithaceae and the subfamilies Mastophoroideae and Melobesioideae. Rhodoliths show sub-spheroidal and sub-ellipsoidal shapes with encrusting, warty and lumpy growth forms. Lumpy growth forms show massive inner arrangements, whereas the encrusting growth forms are usually made of thin thalli and show more loosely packed inner arrangements. The rhodoliths grew on a moderately mobile siliciclastic substrate in a shallow-marine environment with a low net sedimentation rate. It is inferred that the growth of rhodoliths was favored during a relative sea-level rise. During the subsequent sea-level fall, the rhodoliths and associated siliciclastic deposits were resedimented by gravity flows into the deep-sea setting. The analyzed deposits, like other Paleocene–Eocene deposits of the Polish Outer Carpathians, provide no evidence of coeval widespread shallow-marine carbonate sedimentation along the margins of the Outer Carpathian flysch basins.
Similar content being viewed by others

Introduction
The Polish Outer Carpathians are the northernmost segment of Europe’s Alpine orogenic belt. They comprise several tectono-stratigraphic units stacked upon one another as rootless nappes and thrust over the Neogene foredeep and the underlying European Platform (see Książkiewicz 1977). The nappes consist of latest Jurassic–Early Miocene deep-marine deposits, mainly siliciclastic, referred to as ‘flysch’ (Książkiewicz 1975; Geroch and Nowak 1984; Olszewska 1997). The flysch successions have been extensively studied, but much less known are the conditions in coastal and shallow-marine environments of the basin margins. Some authors have suggested periodic development of carbonate platforms along the margins (Rajchel and Myszkowska 1998b; Cieszkowski et al. 2003; Golonka et al. 2003, 2005), but no evidence of widespread carbonate sedimentation has thus far been found. A valuable insight in basin-margin conditions is provided by contemporaneous resedimented components, such as shallow-marine biota and carbonate clasts found in the flysch, particularly in its sandstones and paraconglomerates.
Poorly diversified, shallow-water carbonate debris, frequently coralline algae, occurs commonly in the Paleogene flysch of the Polish Carpathians. The debris occurs as a resedimented component in gravity-flow deposits (Golonka 1974; Leszczyński 1978; Rajchel and Myszkowska 1998a, b; Golonka et al. 2005), except for slivers of sedimentary cover within tectonic klippes near Andrychów (Książkiewicz 1951). Much less common are rhodoliths as well as pebbles and cobbles of coralline algal limestones (Leszczyński 1978; Bassi et al. 2000; Leszczyński et al. 2005). Only in the Inner Carpathians to the south do coralline algal limestones occur in situ in an Eocene transgressive succession (Małecki 1956). Detailed studies of Paleogene rhodoliths have generally been limited to occurrences in Upper Eocene to Oligocene strata, mainly in the Mediterranean region. Resedimented rhodoliths in flysch and the taxonomy of coralline algae from such deposits have seldom been investigated (Leszczyński 1978; Stockar 2000; Puga-Bernabéu et al. 2009).
The present study focuses on an abundant occurrence of rhodoliths in the Paleocene flysch of the central segment of the Silesian Nappe, cropping out at the village of Melsztyn (Fig. 1). The resedimented rhodoliths are the main carbonate component of the generally siliciclastic mass-flow deposits. The outcrop section is the largest exposure of a rhodolith-bearing flysch in the Polish Outer Carpathians, and also the richest in rhodoliths, providing a unique material for sedimentological and taxonomic analysis.

The aims of the study are: (a) to assess the depositional setting of the deep-marine sedimentary succession from its stratigraphic development; (b) interpret the depositional conditions of the mixed siliciclastic-carbonate sediment in its parental shallow-marine environment; (c) to improve the sparse existing knowledge on the range of basin-margin paleoenvironments in the Polish Outer Carpathians; and (d) to review the existing data on coralline algae in the Polish Carpathian flysch and discuss factors controlling their formation. Previous data (Leszczyński and Kołodziej 2004; Leszczyński et al. 2005) are supplemented with new field observations, biostratigraphic evidence and a detailed study of the rhodoliths.
Geological setting
The Silesian Nappe is one of the main thrust-sheets of the Polish Outer Carpathians (Fig. 1a). It is bordered to the south by the thrust-folded Fore-Magura Group, the Dukla Nappe and locally by the Magura Nappe, whereas to the north it is thrust over the Sub-Silesian Nappe. The Late Cretaceous–Oligocene sedimentary succession of the Silesian Nappe, including the studied outcrop section (Fig. 2), was deposited in a deep-water sub-basin of the Carpathian Flysch Sea, referred to as the Silesian Basin (Książkiewicz 1975).

The Silesian Basin was bordered to the south by an intrabasinal ridge, originally called the Silesian Cordillera (Książkiewicz 1956) and more recently referred to as the Silesian Ridge. To the north, the basin was bordered by a ridge known as the Sub-Silesian Cordillera or Sub-Silesian Ridge (Koszarski and Żytko 1965; Fig. 3). The Silesian Ridge was uplifted in the Late Cretaceous to Paleocene time, during the Laramian phase of the Alpine orogeny. The uplift is reflected in the greater thickness and sediment coarseness of the Coniacian–Lower Eocene part of the flysch succession, particularly in the Silesian Nappe (Bieda et al. 1963; Unrug 1968). Paleocurrent directions in this part of the succession indicate sediment derivation mainly from the Silesian Ridge on the southern side of the basin (Książkiewicz 1962; Unrug 1963; Leszczyński 1981).

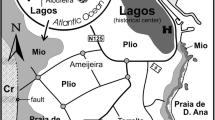
Paleogeographic location of the study area. a Paleocene paleogeography of the European segment of the Tethys and adjoining areas (compiled from Ricou et al. 1986; Dercourt et al. 2000; Vlahović et al. 2005). The red rectangle indicates area shown in b. b Sedimentary basins of the Polish Outer Carpathians in the Paleocene
Sedimentary facies and their distribution suggest deposition in a system of coalescing basin-floor fans (Dziadzio et al. 2005). The increased supply of coarse-clastic sediment from the south indicates an elevated, tectonically active basin margin. In Campanian to Early Eocene time, the Istebna Beds and the overlying Ciężkowice Sandstone were deposited in nearly the whole Silesian Basin. The studied outcrop section is in the north-central part of the Silesian Nappe, where the Istebna Beds and the overlying Ciężkowice Sandstone show many lithofacies similarities and hence are often difficult to distinguish from each other in small, isolated outcrops. The deposits described here were included by Koszarski and Kuciński (1966) in the Lower Istebna Sandstone (i.e., the lower part of the Istebna Beds of the Silesian Nappe) and were ascribed a Senonian age (Bieda et al. 1963; Koszarski 1985), but without any supporting stratigraphic evidence. The data from the present study indicate that these deposits belong to the Upper Istebna Sandstone and represent the Upper Paleocene.
Materials and methods
The field study was carried out in an outcrop section on the eastern slope of the castle hill in Melsztyn (Fig. 1b). The outcrop section is 170 m wide and up to 20 m high, showing a 16-m succession of sandstone and pebbly conglomerate beds (Fig. 4a). In order to understand better the sedimentary succession, it was mapped in strike-parallel direction in a 1.5-km zone over a distance of 4 km towards the northwest. The rhodolith-bearing deposits, up to 11 m thick, form an indistinctly bedded unit (composite bed sensu Wood and Smith 1959) extending over the whole width of the outcrop. The most detailed investigation was done with the use of a climbing rope in the outcrop part directly below the southern margin of the castle ruins, where the deposits are richest in rhodoliths. The texture, mineral composition, and primary structure of deposits were documented in the whole outcrop section. The size, shape, and internal structure of rhodoliths were studied in selected fresh rock samples and also in loose blocks fallen from the outcrop wall. Samples of the best preserved and most representative rhodolith varieties were taken for laboratory analysis.

Lithofacies and stratigraphic position of the rhodolith-bearing flysch succession (the Late Paleocene Upper Istebna Sandstone, Silesian Nappe, Polish Outer Carpathians). a Generalized lithofacies log and stratigraphic position of the flysch succession in Melsztyn. The grey arrow points to the bed segment shown in b and the black arrow indicates the location of samples collected from mudstone intraclasts for foraminiferid biostratigraphy. b Close-up view of the distribution of rhodoliths (marked as white spots and nodules) in a siliciclastic matrix. The white rectangle indicates outcrop detail shown in c. c Indistinct cluster of rhodoliths. d Rhodoliths of different sizes and spheroidal to sub-ellipsoidal shapes occur embedded in a siliciclastic matrix. e Rhodoliths showing lumpy and warty growth forms. f Rhodoliths showing growth-form development from encrusting to lumpy; the inner void spaces in rhodoliths are filled with siliciclastic sediment. Scale bar 1 cm
Foraminifera were analyzed in five samples of rip-up mudstone clasts from the rhodolith-bearing deposits, nine samples of mudshales from a stratigraphically adjacent part of the outcrop section (ca. 10–15 m below the rhodolith-bearing deposits), and seven thin sections of three limestone clasts from the rhodolith-bearing deposits (Fig. 1b). The foraminiferal analyses of fine-grained deposits were performed on samples disaggregated with Glaubert’s salt and subsequently washed through a 63-μm screen sieve. The laboratory study included analysis of the size, shape and internal structure of rhodoliths in six polished slabs and in 30 thin-sections of isolated rhodoliths. Thin-section analysis included also the content and distribution of fossils, microfacies, the mineral and textural composition of rhodolith-bearing sediment, and the characteristics of mudstone intraclasts.
The descriptive terminology for coralline algae growth-forms follows Woelkerling et al. (1993). The family and subfamily circumscriptions are according to Woelkerling (1988), Braga et al. (1993, 2010), Aguirre and Braga (1998), and Braga (2003). The study has taken into account all basic taxonomic uncertainties concerning fossil coralline algae, as discussed by Braga et al. (1993), Braga and Aguirre (1995), Rasser and Piller (1999), Bassi and Nebelsick (2000) and Iryu et al. (2009).
The field study with sampling was carried out by the first two authors (S.L. and B.K.), the analysis of coralline algae was done by the third author (D.B.), whereas the two other authors (E.M. and M.A.G.) analyzed the associated foraminifera assemblages.
Results
The rhodolith-bearing flysch exposed in Melsztyn
The flysch succession in the outcrop section contains a composite bed (sensu Wood and Smith 1959) nearly 4 m thick composed of siliciclastic, granule-bearing to pebbly sandstone that grades upwards into a coarse-grained sandstone with calcitic cement patches (Fig. 4a). The patchy cementation makes sandstone pseudo-nodules stand out from the weathered rock surface. The nodules tend to be aligned parallel to bedding but locally show stock-like vertical grouping. Pebbles are up to 1 cm in size and, like the sand and silt fraction, consist mainly of quartz. Feldspar and mica are subordinate components of the sand and silt fraction, and the sandstone has a low mud content. The base of the bed is hidden by talus, whereas its top is uneven, erosional.
The overlying deposits are a package of indistinct and laterally discontinuous layers of pebble conglomerate and coarse-grained sandstone, rich in rhodoliths (Fig. 4). This rhodolith-bearing unit is 11 m thick and its component layers, up to 2 m thick, are massive (non-stratified) and seem to show normal or inverse grading or virtually lack grain-size grading. The normal-graded layers, are commonly underlain by non-graded ones and contain few rhodoliths, whereas the inversely graded layers, up to 40 cm thick, are rich in rhodoliths, whose abundance also increases upwards. The layer richest in rhodoliths occurs in the middle of the package, is non-graded, and nearly 2 m thick.
The abundance of rhodoliths thus varies from layer to layer and also laterally within the layers, and the rhodolith-rich deposits generally contain carbonate cement. The rhodolith-poor layers tend to be richer in mud and show little or no carbonate cement. The groundmass is dominated by light-grey quartz and the quartz pebbles are up to 3.5 cm in size. Intraformational rip-up clasts of brown to blackish-brown or dark-grey mudstone are up to 1 m in length, some with sandy interlaminae or thin laminated interbeds. The rip-up clasts tend to dip towards the southwest, which may suggest imbrications and transport direction towards the northeast. Pebble- to cobble-sized ?sideritic clasts occur mainly in the lower part of the rhodolith-bearing package.
The local mapping has shown that the rhodolith-bearing deposits are cropping out at several other localities up to 3.5 km away from the outcrop in Melsztyn (Fig. 1b). These other outcrops are in a stream bed and banks, up to 2 m high and 50 m long, and show deposits similar to those exposed in Melsztyn.
Underlying and overlying deposits
Deposits underlying the rhodolith-bearing coarse-grained succession are poorly exposed in Melsztyn, covered with talus, and accessible nearby in only two small outcrops in the valley below. The closest outcrop is ~50 m away to the southeast and stratigraphically ~10 m below the outcrop section studied in Melsztyn. It shows greenish-grey mudshales interbedded with thin sandstone beds, but the exposed stratigraphic thickness is only ~1 m. The other outcrop, located ~50 m to the southeast from the former, shows faulted and variably tilted grey to dark-grey mud- and clayshales with rare thin sandstone interbeds, and a total exposed stratigraphic thickness of ~4 m. This outcrop section is estimated to be located ca. 5 m below that of the former outcrop.
Other outcrops of the deposits underlying the rhodolith-bearing succession are as far away as 0.7 km to the west and 2.3–2.5 km to the northwest (coordinates N: 49°52′54.66″ to 49°53′3.6″, E: 20°44′26.86″ to 20°44′31.41″). Except for the outcrops at the passage to the coarse-grained, rhodolith-bearing deposits and one 6-m-thick package where thick-bedded sandstones interbedded with mudshales occur, they show dark greenish-grey clayey to silty shales with thin sandstone and siderite interbeds. The exposed stratigraphic thicknesses range from 50 cm to several tens of meters. Exact thickness in the widest outcrop (almost 360 m) is unclear as the deposits show tectonic tilting and brecciation indicative of thrust faulting. The deposits in the widest outcrop form the core of an anticline.
As to the deposits overlying the rhodolith-bearing succession in Melsztyn, their nearest outcrop is ~2.9 km to the northwest (coordinates N: 49°53′3.6″, E: 20°44′31.41″) and stratigraphically just above this succession. Their exposed thickness is several tens of meters. In the lower, 4-m-thick part of the section, these are dark-grey mudshales and clayshales. The higher part of the section is built of variegated (mainly red) mudshales and clayshales.
Rhodoliths and coralline algal debris
Rhodoliths and coralline algal debris are the most abundant carbonate components (up to 90 %) of the analyzed rhodolith-bearing sandstones and conglomerates exposed in Melsztyn.
Two types of rhodoliths were distinguished (Figs. 4d–f, 5): (1) abundant sub-ellipsoidal laminar rhodoliths with a looser internal packing, but few constructional voids; and (2) common sub-spheroidal rhodoliths with a massive, well-packed interior. They are mainly 2.5–3.5 cm in size, with a maximum diameter of 6 cm, and their growth-form ranges from encrusting to warty or lumpy.

Polished surfaces of the studied Upper Paleocene rhodoliths. a–d Laminar, sub-ellipsoidal rhodoliths with loosely packed inner structure and encrusting to lumpy growth-forms. e–f Massive sub-spheroidal rhodoliths with encrusting growth forms. g Fragment of a rhodolith showing a thick laminar succession of superimposed encrusting thalli. Scale bar 1 cm
The laminar rhodoliths show common encrusting, warty and lumpy growth-forms, with the outer surface commonly made of lumpy thalli. The massive rhodoliths are characterized by common encrusting and rare warty coralline growth forms; the last growth stage of these rhodoliths typically consists of encrusting thalli. However, in both rhodolith shapes growth-form successions from encrusting to lumpy or vice versa are quite common.
Asymmetrical inner arrangements are dominant, but symmetrical ones are also present (Figs. 4d–f, 5). The asymmetry is usually greatest in the inner part of the rhodolith and decreases outwards. The rhodolith nuclei are relatively small and consist of an infilling of siliciclastic material or rarely of massive coralline thallus, coral, or mollusc shell. Siliciclastic matrix also fills the inner voids of rhodoliths.
The assemblage of coralline red algae consists of the family Sporolithaceae and the subfamilies Mastophoroideae and Melobesioideae (Table 1; Figs. 6, 7). The predominant coralline taxa are Sporolithon Heydrich (S. sp. A), Lithothamnion Heydrich (L. sp. A, L. sp. B) and Mesophyllum Lemoine (M. sp. A). Subordinate taxa are Karpathia Maslov (Bassi et al. 2005), Spongites Kützing (S. sp. A), Neogoniolithon Setchell and Mason (N. cf. contii (Mastrorilli) Quaranta, Vannucci & Basso), and Lithoporella Foslie (L. sp. A) and the peyssonneliacean Polystrata alba (Pfender) Denizot, 1968. Rhodoliths can be multi-generic (no more than two genera) and multispecific (no more than two species). Monogeneric rhodoliths consist of Sporolithon sp. A and Lithothamnion sp. B. Accessory components of the rhodoliths include bryozoans, serpulids (relatively common and large in size), and encrusting foraminifera (acervulinids, planorbulinids).

Thin-section photomicrographs of coralline algae from the studied rhodoliths. a Sporolithon sp. A, lumpy protuberance with sporangial compartments on a row of elongated cell layers (arrow), grouped in sori. b Lithoporella sp. A, single layers of palisade cells with cell fusion (arrows). c Karpathia sp., superimposed undulating encrusting thalli showing cell fusion between adjacent cells (arrow). d Spongites sp. A, section through superimposed encrusting thalli showing a uniporate conceptacle with long cylindrical pore canal (p) and the non-coaxial core filaments (arrowhead). Scale bar 0.2 mm in a, c–d, and 0.1 mm in b

Thin-section photomicrographs of coralline algae from the studied rhodoliths. Mesophyllum sp. A; a superimposed encrusting thalli with several multiporate conceptacles (arrow) and the coaxial ventral core (vc); b longitudinal section of a multiporate conceptacle. Lithothamnion sp. A; longitudinal sections of a protuberance (c) and encrusting superimposed thalli (d) showing the multiporate conceptacles and the non-coaxial ventral core (arrow). e Lithothamnion sp. B, superimposed encrusting thalli with non-coaxial ventral core (vc) and a longitudinal section of multiporate conceptacle (arrow). f Polystrata alba (Pfender) Denizot, longitudinal oblique section of an encrusting thallus. Scale bar 0.5 mm in a, and 0.2 mm in b–f
The rhodoliths commonly show borings, with smooth rounded outlines in cross-sections, in four diameter ranges: 0.07, 0.12–0.1, 0.4–0.6, 2.0 mm. Some borings show irregularly shaped, single or multiple, smooth-walled, oval chambers corresponding to the ichnogenus Entobia (Fig. 8a). Networks of branching micro-galleries with irregular changes in diameter between 0.10 and 0.20 mm can be ascribed to the activities of fungi, algae, bacteria, and/or sponges (Fig. 8b). Two galleries occasionally converge in a Y-shape pattern, and the borings in some cases seem to extend from the inner to the outer part of the rhodolith. Some borings enclose foraminifera, which may belong to the sedimentary infill or represent the boring forms or cavity dwellers (coelobites; see Schlagintweit 2005).

Thin-section photomicrographs of studied rhodoliths showing the morphologies of common bioerosion traces. a Rounded multi-chambers of sponge-related borings referable to the ichnogenus Entobia. b Microgalleries with rounded transversal sections produced by fungi, algae, bacteria and/or sponges. Scale bar 0.5 mm
Bioclasts and algal limestone pebbles
Bioclasts consist mainly of coralline algal debris which is usually smaller than 1 cm, representing parts of fruticose and lumpy plants as well as superimposed encrusting thalli associated with other encrusting organisms. The coralline algal debris is invariably abraded, composed mainly of Mesophyllum, Lithothamnion, and Sporolithon.
Molluscs are rare in the samples, represented mainly by ostreids scattered in siliciclastic sediment and present inside some rhodoliths. Sporadic are poorly preserved fragments of originally aragonitic bivalve shells (“ghost structures”). Bryozoans, echinoderm plates, and small fragments of corals are very rare in the sediment. Foraminifera are represented by miliolids and rotaliids (Fig. 9).

Benthic foraminiferids from a carbonate clast (sample SL 3) in the Late Paleocene Upper Istebna Sandstone at Melsztyn. a Miliola ?andrusovi Samuel, Köhler and Borza. b Miliola ?andrusovi Samuel, Köhler and Borza. c Mississippina binkhorsti (Reuss). d Cocoarota orali İnan. e Planorbulina cretae (Marsson). f Miniacina sp. Scale bar 1 mm
Three pebble-sized clasts of algal limestones have been found, one composed of fruticose coralline algal fragments (Mesophyllum, Lithothamnion) in a micritic matrix, and the other two representing algal-bioclastic grainstone/packstone. The limestones contain also bivalves, echinoderms, foraminifera, serpulids, and bryozoans as bioclasts, but no larger foraminifera nor green calcareous algae have been found. Quartz and glauconite grains are minor, and also rhodoliths in these pebbles are rare and very small.
Foraminifera in the rhodolith-bearing and underlying deposits
Foraminiferal content has been examined in: (1) nine samples (M2, 3, 6A–8, 10–13) collected from the grey to greenish-grey mudstones stratigraphically located from ~10 to 20 m below the rhodolith-bearing succession; (2) five samples (M4, 6, 14–16) of mudstone rip-up clasts found in the lower part of the rhodolith-bearing succession; and (3) thin-sections of rhodoliths and the three pebbles of algal limestone found in a stream-bank outcrop of the rhodolith-bearing succession ~1.5 km northwest of the outcrop in Melsztyn (coordinates N: 49°52′29.14″, E: 20°44′44.15″).
Foraminifera are rare in samples M2, 3, 8, and M10–13, which show the low-diversity, impoverished flysch-type assemblages of deep-water agglutinated forams (the DWAF association of Kuhnt and Kaminski 1989). Large tubular forms of Nothia, Ammosphaeroidina pseudopauciloculata (Mjatliuk) and Gerochammina lenis (Grzybowski), with solitary specimens of the planktonic genera Heterohelix, ?Rugoglobigerina, and Globigerinelloides, have been found in sample M10.
Samples M6A and M7 yielded a typical DWAF assemblage with Placentammina placenta (Grzybowski), Glomospira diffundens Cushman and Renz, Rzehakina epigona (Rzehak), Rzehakina fissistomata (Grzybowski), Caudammina excelsa (Dylążanka), Hormosina velascoensis (Cushman), Ammosphaeroidina pseudopauciloculata (Mjatliuk), Thalmannammina subturbinata (Grzybowski), and Trochamminopsis cf. altiformis (Cushman and Renz), accompanied by tubular forms of Nothia and Rhabdammina as well as by Trochamminoides div. sp. and Paratrochamminoides div. sp. Sample M6A contains also Kalamopsis grzybowskii (Dylążanka), Ammobaculites cf. agglutinans (Orbigny) emend. Bartenstein, 1952 and relatively abundant C. excelsa, whereas Annectina grzybowskii (Jurkiewicz) has been found in sample M7.
In sample M10, apart from large tubular forms of Nothia, Ammosphaeroidina pseudopauciloculata (Mjatliuk) and Gerochammina lenis (Grzybowski), single specimens of the planktonic genera Heterohelix, ?Rugoglobigerina and Globigerinelloides have been found.
The mudstone-clast samples M4, M6 and M14–16 yielded mostly poor assemblages of agglutinated foraminifera. Sample M4 was barren. The relatively rich assemblage in sample M6 included a few pyritized specimens of ?Praebulimina, Nonionella, and fragments of Chilostomella and Globigerina s.l.
Foraminifera in one of the algal limestone clasts (SL3), examined in three thin-sections, consist of small benthic taxa, both agglutinated and relatively more numerous, calcareous. Agglutinated foraminifera include Dorothia sp. A, Dorothia sp. B, and Clavulinoides sp. Calcareous benthic taxa are represented by Miliola ?andrusovi Samuel, Köhler and Borza (Fig. 9a, b), Mississippina binkhorsti (Reuss) (Fig. 9c), Cocoarota orali İnan (Fig. 9d) and encrusting Planorbulina cretae (Marsson) (Fig. 9e) as well as Miniacina sp. (Fig. 9f). Some other forms belonging to miliolids and rotaliids have also been found, but their exact taxonomic identification was hindered by sparse and non-diagnostic sections.
Stratigraphic re-evaluation of the succession
Lithostratigraphic comparisons
In terms of their composition and sedimentary facies, the rhodolith-bearing deposits resemble the Ciężkowice Sandstone of latest Paleocene–Early Eocene age. The similarity includes especially the occurrence of rhodoliths, which are known from many outcrops of the Ciężkowice Sandstone (Leszczyński 1978), but not from the Senonian Lower Istebna Sandstone to which these deposits were ascribed by Koszarski and Kuciński (1966). However, underlying and overlying dark-grey shales render the rhodolith-bearing deposits similar to the Upper Istebna Sandstone of the Late Paleocene (Unrug 1963; Bieda et al. 1963; Geroch and Nowak 1984; Koszarski 1985), rather than to the Ciężkowice Sandstone, as the latter formation is generally sandwiched between variegated, reddish-greenish shales. The Ciężkowice Sandstone seems to be absent in the study area, as noted earlier by Birkenmajer et al. (1962) and Koszarski and Kuciński (1966).
The underlying shale-rich succession with its facies and stratigraphic position correspond to the Lower Istebna Shale of latest Senonian–Early Paleocene (Bieda et al. 1963; Geroch and Nowak 1984; Koszarski 1985; Fig. 2), whereas the dark-grey shales overlying the rhodolith-bearing deposits appear to corresponds to the Upper Istebna Shale of Late Paleocene age (Bieda et al. 1963; Geroch and Nowak 1984; Koszarski 1985). Although these overlying shales seem here to be considerably thinner than the regional unit of Upper Istebna Shale, the difference can be attributed a tectonic structural thinning.
Biostratigraphic inferences
In this section, the tentative lithostratigraphic correlations will be assessed on the basis of biostratigraphic data from the shales underlying the rhodolith-bearing flysch and from the mudshale and limestone clasts found in the coarse-grained siliciclastic flysch.
The underlying shale succession, correlated with the Lower Istebna Shale, contains relatively rare and long-ranging taxa of agglutinated foraminifera (samples M2, 3, 8, 10–13). Most of the taxa found in samples M6A and M7 are common in the Campanian–Paleocene assemblages of agglutinated forams in the Polish Outer Carpathians (Olszewska 1997; Kaminski and Gradstein 2005). However, some more exact inferences can be made. Rzehakina fissistomata has its first appearance in the Late Maastrichtian but if relatively abundant, it can serve as an index species of the Paleocene Rzehakina fissistomata zone of Geroch and Nowak (1984).
The total stratigraphic range of Annectina grzybowskii is Campanian–Paleocene (Kaminski and Gradstein (2005), but the taxon is thought to be characteristic of Paleocene foraminifera assemblages (Malata 2002). Accordingly, the shale represented by sample M7 is most likely of a Paleocene age. Sample M6A shows a considerable amount of C. excelsa, which is a characteristic component of Campanian–Maastrichtian assemblages in the Polish Outer Carpathians (Olszewska 1997; Malata 2002), but does not preclude a Paleocene age. The solitary specimens of the planktonic genera Heterohelix, ?Rugoglobigerina, and Globigerinelloides found in sample M10 are generally indicative of a Late Cretaceous age. The foraminifera assemblages thus support regional correlation with the Late Senonian–Early Paleocene Lower Istebna Shale.
The mudshale intraclasts in the flysch show mainly long-ranging foraminiferal taxa, but at least these found in sample M6 suggest an age not older than Paleocene. More indicative are foraminiferal taxa in the limestone clasts. The species Miliola ?andrusovi, found in one of the three clasts, is known from the Paleocene of the West Carpathians (Samuel et al. 1977), though it has recently been reported also from the Lower to Middle Illerdian (Lower Eocene) of Sardinia (Dieni et al. 2008). Cocoarota orali has not previously been reported from the Carpathian flysch, but was described from the Upper Maastrichtian–Lower Lutetian of Turkey (İnan 2003). Mississippina binkhorsti is common in Paleocene to Eocene shallow-water carbonate deposits (Samuel et al. 1972; İnan and İnan 2008; Olszewska 2009). Planorbulina cretae has been reported mainly from the Upper Cretaceous and Paleocene (Bignot and Larsonneur 1969; Samuel et al. 1972; Köhler and Buček 2005; Dieni et al. 2008), though found also in the Upper Paleocene–Lower Oligocene of Bulgaria (Dimitrova 2003). The co-occurrence of the above-mentioned species of small foraminifera suggests a Paleocene–Early Eocene (Illerdian) age of the limestones from which the clasts were derived. This evidence generally supports regional correlation of the coarse-grained flysch succession with the Upper Istebna Sandstone.
Discussion
Origin of the rhodolith-bearing deposits
The coarse-grained sedimentary succession shows features indicative of deposition by submarine sediment-gravity flows, mainly high-density turbidity currents and debris flows (sensu Lowe 1982). The lowest composite bed in the outcrop section shows a repeated normal grading, which suggests two consecutive surges of a turbulent high-density flow. Multiple normal grading in the overlying rhodolith-bearing succession suggests deposition by several consecutive surges of one or more similar flows. The amalgamated mass-flow deposits and their facies resemble closely those of the Istebna Beds and the Ciężkowice Sandstone (Unrug 1963; Leszczyński 1981), formed at bathyal depths (Koszarski and Żytko 1965; Książkiewicz 1975; Leszczyński 1981), and their origin can be attributed to a deep-sea channel-fill or channel–lobe transition zone (Palanques et al. 1995; Wynn et al. 2002). Either depositional setting might explain the occurrence of such deposits (Janbu et al. 2007; Wynn et al. 2002) and the emplacement of debris flows far away from the sediment source (Janbu et al. 2007; Ito 2008). Imbricated rip-up clasts indicate paleotransport towards the northeast. Paleogeographic reconstructions (e.g., Książkiewicz 1962), suggest that the probable shallow-marine environment for the growth of coralline algae and formation of rhodoliths was on the Silesian Ridge at the southern margin of the Silesian Flysch Basin. The narrow ridge is unlikely to have hosted extensive river systems and large deltas, and the coarse-grained siliciclastic deposits thus suggest resedimentation from a fan-delta (coastal alluvial fan) system.
Paleoecology of rhodoliths
The examined rhodoliths and associated calcareous benthic biota represent shallow-marine biogenic components thriving mainly in the photic zone, above the storm wave base. Rhodoliths are free-living nodules composed predominantly of non-geniculate coralline red algae with an admixture of other encrusting organisms (peyssonneliacean algae, encrusting foraminifera, serpulids, bryozoans; e.g., Bosence 1983a; Marrack 1999; Bassi et al. 2009, 2011). Their development is controlled by an array of environmental factors, and hence rhodoliths are a valuable indicator of paleoenvironmental conditions (e.g., Bosence 1983b; Nebelsick and Bassi 2000; Nalin et al. 2008; Bassi et al. 2009, 2010).
The dominance of Sporolithon and melobesioids in the analyzed rhodoliths suggests a low light-intensity setting. Despite the cosmopolitan distribution of the group, Sporolithon is restricted mainly to tropical latitudes (Johnson 1963; Adey and Macintyre 1973; Johansen 1981), where it primarily occurs in relatively deep-water and cryptic sites (Adey 1979, 1986). However, the species, S. ptychoides Heydrich seems to be the only one confidently reported as currently inhabiting the Mediterranean Sea, where it is common on submarine cliffs and can occur down to infra- and circalittoral settings (e.g., Alongi et al. 1996; Bressan and Babbini 2003; Braga and Bassi 2007; Braga et al. 2009). Modern melobesioids, together with Sporolithon, increase in abundance with water depth in tropical and non-tropical seas (Adey 1979; Adey et al. 1982; Lund et al. 2000), but at higher latitudes where water turbidity is greater they are also common in shallower water rhodoliths and pebble coatings (Bosence 1983b; Foster 2001; Braga et al. 2009).
A shallow-marine origin of the studied rhodoliths is indicated also by the occurrence of subordinate mastophoroids (with three genera), which tend to predominate in shallow-water settings of modern tropical (e.g., Borowitzka and Larkum 1986; Braga and Aguirre 2004; Flamand et al. 2008) and temperate seas (e.g., Aguirre et al. 2000; Lund et al. 2000; Braga and Aguirre 2001). Fossil examples of shallow-water rhodolith deposits characterized by sporolithaceans, melobesioids, and mastophoroids have been described from the Upper Eocene and Oligocene of the western Tethys (e.g., Bassi 1998, 2005; Nebelsick et al. 2000, 2005; Rasser 2000; Rasser and Nebelsick 2003; Rasser and Piller 2004; Brandano et al. 2009; Bassi and Nebelsick 2010).
Rhodolith inner structure in several modern environments has been related to depth, sedimentation rate, and hydrodynamic regime (e.g., Minnery 1990; Bosence 1983a, b, 1991; Marrack 1999; Lund et al. 2000; Bassi et al. 2009). The rhodoliths of deeper water or low hydrodynamic-energy environments show large constructional void spaces, may lack an identifiable nucleus, and are composed of thin, irregular, encrusting thalli (e.g., Braga and Martín 1988; Barattolo et al. 2007; Bassi 1998, 2005; Nalin et al. 2008; Bassi et al. 2006, 2009, 2010). The predominantly asymmetrical and loosely packed inner structure of the studied rhodoliths in the present case indicates occasional overturning in an environment with moderate water turbulence.
The occurrence of siliciclastic material as an infill of voids in the rhodoliths indicates their growth on a substrate rich in such material. The abundance of borings in the rhodoliths supports the notion of an environment with a low net sedimentation rate, allowing a relatively long residence time on the substrate. The boring activity took place during the active growth of the rhodoliths. The rounded shape of the borings, their size, curved course and scattered occurrence suggests the activity of sponges, fungi, algae, and polychaete worms (e.g., Rice and Macintyre 1982; Bromley 1992, 1994; Checconi et al. 2010). Microborings recognized in a coral in one of the rhodolith nuclei are similar to those produced or inhabited by foraminifera, as described from Paleocene peri-reefal deposits in Austria (Schlagintweit 2005).
The low abundance of biota other than coralline algae and ostreids may suggests a non-hospitable environment, although this impression may partly be due to a taphonomic bias resulting from the low fossilization potential of aragonitic skeletons, mostly molluscs (Wright et al. 2003). The proportion of aragonitic shells may have originally been higher, as suggested by the patchy calcitic cementation of both the rhodolith-bearing deposits and the underlying ones. The same is suggested also to some extent by the “ghost structures” of molluscs.
Depositional model
The integrated analysis of regional stratigraphic correlation, flysch lithofacies, their biogenic components and rhodolith characteristics led us to the following depositional model of the rhodolith-bearing flysch succession in Melsztyn. Two main depositional stages are distinguished: (I) accumulation of mixed siliciclastic-carbonate sediment with rhodolith growth, in a shallow-marine environment of the southern margin of the basin (Fig. 10a) and (II) subsequent resedimentation into the deep-water settings (Fig. 10b). The two stages are thought to have been related to relative sea-level change. This interpretation is suggested by the spatially restricted and relatively short-lived, episodic supply of sediment enriched in contemporaneous neritic biota and carbonate rock debris. Regional evidence from the Outer Carpathian Flysch indicates that the shelf zone of the basin margin was dominated by siliciclastic sedimentation. The production of carbonate material was apparently restricted to one or more favorable areas of the shelf, with suitable bathymetry, hydraulic energy level, substrate composition, and sediment accumulation rate (Nalin et al. 2010). Shelf erosion is known to be greatly enhanced during a pronounced sea-level fall and lowstand (Catuneanu 2006), when also the local niches of carbonate production would likely become subject to resedimentation (e.g., Aybulatov and Drushchits 1986; Vecsei and Sanders 1997). In a tectonically active basin, the shedding of inner-shelf sediment may indicate a relative sea-level fall due to uplift of the basin margin.

Depositional model for the shallow-water origin and ultimate deep-water emplacement of Late Paleocene mixed carbonate-siliciclastic deposits in the Outer Carpathian flysch basin. a Stage I A relative sea-level rise up to highstand creates local conditions for bioclastic carbonate production on a narrow, fan delta-hosting siliciclastic shelf at the basin margin. b Stage II A subsequent sea-level fall and lowstand cause erosion and resedimentation of the rhodolith-bearing deposits into deep water. Not to scale
In summary, we infer that the depositional stage I commenced with the onset of relative sea-level rise (Fig. 10a), when the accommodation space of the shelf zone increased, erosion declined, and sediment accumulation prevailed. Significant production of biogenic carbonates would occur in shallow-water areas with a suitable hydraulic energy and limited sediment supply (e.g., Monstad 2000; Philip 2003; Nalin et al. 2008). This stage would probably persist until at least the early highstand (Sarg 1988; Tucker and Wright 1990; Leinfelder 1993; Brachert et al. 2003), whereas erosion of the inner shelf might have commenced with the fall of relative sea level and persisted until the lowstand phase (Fig. 10b).
As discussed in the previous chapter, the rhodoliths were apparently formed on a moderately mobile sandy siliciclastic substrate, in restricted shoal-water, which suggests coarse-grained coastal shoals provided commonly by fan deltas (e.g., Dutton 1982). Arguably, fan deltas are the only restricted shelf areas where considerable volumes of submature to mature gravel and sand can accumulate and where biogenic carbonate sedimentation may occur in abandoned or inactive shoal-water zones (e.g., Dabrio and Polo 1988). The subordinate admixture of carbonate components (mainly rhodoliths and ostreids) in the resedimented siliciclastic material suggests a volumetrically limited carbonate production and the limited thickness and spatial extent of the rhodolith-bearing flysch suggest further that the carbonate production was relatively short-lived and areally restricted. The common occurrence of boring activity developed during the rhodolith growth testifies to periods with a very low sedimentation rate.
The occurrence of bioclast-free deposits below the rhodolith-bearing deposits in the outcrop section in Melsztyn implies that, in terms of an erosional shelf stripping, before resedimentation in deep water, the rhodolith production on the shelf had ceased at some point and the rhodolith-rich sediment was buried by bioclast-free sediment. The burial might most likely have occurred during normal regression (sea-level highstand), when seafloor aggradation and shoreline progradation prevailed along the basin margin. The immediate burial of bioclast-rich sediment by barren sediment could first of all have happened in areas overwhelmed by sediment supply from river mouths and subject to delta progradation.
Stage II involved erosion of inner shelf and resedimentation of rhodoliths and associated siliciclastic sediments in deep-water. It might have commenced with a pronounced fall of relative sea level, and persisted until the lowstand phase (Fig. 10b). Such a sedimentary response to sea level change is characteristic of siliciclastic shelves (Catuneanu 2006) like mostly those of the Carpathian flysch basins.
Comparison with other rhodolith-bearing deposits and algal limestones in the Paleogene flysch of the Polish Outer Carpathians
Coralline algal detritus is a significant local component in some parts of the Paleogene flysch of the Polish Outer Carpathians. This carbonate material is scattered in sandstones and granule to pebble conglomerates in almost all of the lithostratigraphic units, particularly those spanning the Paleocene–Eocene and the Eocene–Oligocene boundary (e.g., Cieszkowski et al. 2003). In situ coralline algal limestones in the Polish Outer Carpathians have been reported only from the relics of Paleocene shallow-marine cover of tectonic klippes near Andrychów (Książkiewicz 1951). The existing regional literature and a cursory study of the outcrop in Targanice by one of us (SL), suggest that these limestones occur there in coralline algal debris facies. Some lithostratigraphic flysch units, such as the Paleocene Szydłowiec Sandstone, Czerwin Sandstone, and Bircza Limestone, or the Early Oligocene Łużna Limestone and Skalnik Limestone, are characterized by the common occurrence of coralline algal debris (Książkiewicz 1951; Rajchel and Myszkowska 1998a; Cieszkowski et al. 2005; Leśniak et al. 2005). Other units, such as the latest Paleocene–Early Eocene Ciężkowice Sandstone, contain coralline algal debris only in some beds (Bieda et al. 1963; Leszczyński 1978; Koszarski 1985; Rajchel and Myszkowska 1998a). It should be noted that the units traditionally referred to in the literature as limestones (e.g., the Bircza Limestone, the Łużna Limestone, the Skalnik Limestone) are actually carbonate-rich sandstones to sandy limestones (see Bieda 1962, 1968).
In the above-mentioned Paleocene–Eocene lithostratigraphic units, the rhodoliths occur mainly in the Ciężkowice Sandstone (Leszczyński 1978). The best-known outcrop of this unit on the eastern side of Lake Rożnów (~20 km south of Melsztyn), is presently poorly exposed. It shows paraconglomerates of the lowermost part of the unit (latest Paleocene) containing pebbles and cobbles of rhodolith limestone, algal debris limestone, and Discocyclina limestone mixed with “exotic” pebbles and cobbles of crystalline and other sedimentary rocks. The algal limestones are packstones/grainstones and rhodolith floatstones. Coralline algae are represented by the genera Mesophyllum, Lithothamnion, Lithoporella, Spongites, Karpathia sphaerocellulosa, and Sporolithon, accompanied by large foraminifera such as Discocylina spp. (Leszczyński 1978). The rhodoliths scattered in the sandstones and pebbly conglomerates are similar to those in the Melsztyn outcrop section.
In the village of Skawinki near Lanckorona, the Oligocene Krosno Beds of the Silesian Nappe contain an olistostrome, with pebbles and cobbles of limestones similar to those in the Ciężkowice Sandstone. The limestones represent rhodolith facies, coralline algal packstone/grainstone facies, and crustose coralline algal facies. Coralline algae are represented by the genera Mesophyllum, Lithothamnion, Lithoporella, Spongites, Neogoniolithon, and Sporolithon. Coral fragments are rare, representing four species. The associated large foraminifera suggest a Middle–Late Eocene age of the limestones, which are apparently free of siliciclastic material (Bassi et al. 2000).
Rajchel and Myszkowska (1998b), Cieszkowski et al. (2003) and Golonka et al. (2003) suggested a carbonate platform and algal reef provenance of the algal debris and the clasts of the bioclastic algal limestones found in the Polish Outer Carpathian flysch, but no convincing proof of such a provenance was provided. In reality, the rare occurrences of pebbles and cobbles of algal limestones, and particularly crustose coralline algal limestone suggest that there were probably no extensive carbonate platforms or shelves dominated by carbonate sedimentation along the margins of the Carpathian flysch basins. The benthic communities, particularly coralline algae, were only locally and occasionally able to thrive on the basin-margin shelf and contribute abundant carbonate components to the siliciclastic sediment.
Relatively diverse resedimented debris of colonial and solitary corals (some 15 species of 7 genera), albeit rare and small, occur in the Late Paleocene Babica Clays of the Skole Nappe. However, they represent level-bottom assemblages and not reefal ones, as indicated also by the lack of a biogenic or abiogenic framework (Kołodziej and Stolarski 2000). The Paleocene reefal limestones known from the Inner Carpathians in Slovakia are irrelevant, as they represent a different geotectonic domain (Scheibner 1968; Köhler et al. 1993, Krobicki et al. 2004; Köhler and Buček 2005).
Remarks on Paleogene carbonate shelf sedimentation in the Polish Outer Carpathians
The Paleogene neritic carbonate debris found in the deep-water flysch of the Polish Carpathians has an irregular distribution limited to certain areas and relatively narrow stratigraphic intervals. The distribution may reflect derivation from isolated sources and over short periods. In this section, we discuss briefly the paleo-oceanographic and geodynamic factors that may have controlled carbonate sedimentation in the shelf zones of the flysch basin.
The occurrences of resedimented Paleocene–Eocene coralline algae, mainly as debris and rarely as rhodoliths or pebbles and cobbles of algal limestone, indicates some local shallow-marine environments suitable for algal growth at the basin margin. However, evidence of pure carbonate algal sedimentation is sparse, since algal limestone pebbles and cobbles are rare.
The Paleocene Carpathian flysch was deposited in a mid-latitude arm of the northern Tethys (see Scheibner and Speijer 2008), and the occurrence of coralline algal facies there may reflect the general increase in the diversification of coralline algae since the Late Paleocene (Aguirre et al. 2000, 2007). For comparison, well-developed algal facies, including crustose, are known from Eocene basins at similar paleolatitudes, such as the Eocene Alpine Foreland (Rasser and Piller 2004).
As suggested by the present study, the shelf niches for carbonate sedimentation may have been created episodically by a major sea-level rise and early highstand and have subsequently been removed by erosion and resedimentation during the relative sea-level fall and lowstand (major forced regression). The Carpathian flysch basins were tectonically active and their shelf zones were likely narrow, fringing a structural ridge, whereby the sea level might have occasionally fallen below the shelf edge.
According to Nalin et al. (2008), the deposition of Cenozoic rhodolith-bearing sediments occurred commonly during marine transgressions, particularly (though not uniquely) in non-tropical areas. At the advanced stage of transgression, these rhodolith-bearing deposits would be covered by fine-grained subneritic facies (Nalin et al. 2008) or buried by shoreline progradation during normal regression (Catuneanu 2006). For example, most of the Eocene shallow-water carbonates, including rhodolith facies, in the Alpine–Pannonian region were deposited with a major basal unconformity over Mesozoic rocks and overlain by deeper-water siliciclastic sediments (Sahy et al. 2008).
Little is known about the Oligocene shallow-water carbonate sedimentation in the Polish Outer Carpathian flysch basins. Sparse resedimented carbonate debris occurs scattered chiefly in the Lower Oligocene. The lack of such contemporaneous debris in the higher part of the Oligocene may possibly reflect isolation of the northern part of Tethys, including the Outer Carpathian basins, from the main part of the ocean (Báldi 1984; Rögl 1999).
Conclusions
-
1.
The studied section in Melsztyn, Polish Outer Carpathians, is the largest and most rhodolith-rich outcrop of the siliciclastic flysch of the Silesian Nappe.
-
2.
The flysch lithofacies and regional stratigraphic correlations suggest that these rhodolith-bearing deposits represent the Upper Istebna Sandstone deposited in the Late Paleocene, and not the Senonian Lower Istebna Beds as earlier suggested in the literature.
-
3.
The analyzed rhodoliths and sparse associated biota (small benthic foraminifera, ostreids and fragments of echinoderms, bryozoans, and corals) occur as resedimented components in deep-marine siliciclastic conglomerates to coarse-grained sandstones deposited by high-density turbidity currents and debris flows. The sediment is inferred to have been derived from a fan-delta system developed on the shelf of the Silesian Ridge bordering the Silesian Basin from the south.
-
4.
The rhodoliths are sub-spheroidal to sub-ellipsoidal in shape, with encrusting, warty, and lumpy growth forms. Lumpy forms have massive inner structure, whereas the encrusting forms are usually made of thin thalli and have a more loosely packed inner structure. The inner structure indicates rhodolith formation in an environment of moderate water turbulence. Common borings in the rhodoliths (ichnogenus Entobia and microborings) indicate a low net sedimentation rate.
-
5.
The rhodoliths are composed mostly of coralline red algae with seven genera and eight species representing the family Sporolithaceae and the subfamilies Mastophoroideae and Melobesioideae. Sporolithon, Lithothamnion, and Mesophyllum are the dominant taxa. Karpathia, Lithoporella, Neogoniolithon and Spongites are subordinate.
-
6.
The origin of the rhodoliths and associated biogenic carbonates and their emplacement in deep water as an admixture of siliciclastic flysch are inferred to have been related to major relative sea-level changes. Local niches for biogenic carbonate development on the shelf are thought to have formed during a major sea-level rise and early highstand, with rhodoliths growing on moderately mobile siliciclastic substrates under a low net sedimentation rates. The rhodoliths with the host siliciclastic sediment were then eroded and resedimented into deep water during a subsequent fall and lowstand of the relative sea level (major forced regression). The lack of, or poor early cementation allowed mobilization of skeletal particles and their resedimentation by mass flows.
-
7.
There is no evidence of widespread carbonate sedimentation at the shallow-water margins of the Outer Carpathian flysch basins during the Paleogene. Significant algal sedimentation dominated by maerl facies was episodic and occurred only locally.
References
Adey WH (1979) Crustose coralline algae as microenvironmental indicators in the Tertiary. In: Gray J, Boucot AJ (eds) Historical biogeography, plate tectonics and the changing environment. Oregon State University Press, Corvallis, pp 459–464
Adey WH (1986) Coralline algae as indicators of sea-level. In: van de Plassche O (ed) Sea-level research: a manual for the collection and evaluation of data. Free University of Amsterdam, Amsterdam, pp 229–279
Adey WH, MacIntyre IG (1973) Crustose coralline algae: a re-evaluation in the geological sciences. Geol Soc Am Bull 84:883–904
Adey WH, Townsend RA, Boykins WT (1982) The crustose coralline algae (Rhodophyta: Corallinaceae) of the Hawaiian Islands. Smithson Contrib Mar Sci 15:1–74
Aguirre J, Braga JC (1998) Redescription of Lemoine’s (1939) types of coralline algal species from Algeria. Palaeontology 41:489–507
Aguirre J, Riding R, Braga JC (2000) Diversity of coralline red algae: origination and extinction patterns from the Early Cretaceous to the Pleistocene. Paleobiology 26:651–667
Aguirre J, Baceta JI, Braga JC (2007) Recovery of marine primary producers after the Cretaceous–Tertiary mass extinction: Paleocene calcareous red algae from the Iberian Peninsula. Palaeogeogr Palaeoclimatol Palaeoecol 249:393–411
Alongi G, Cormaci M, Furnari G (1996) On the occurrence of Sporolithon ptychoides Heydrich (Corallinales, Sporolithaceae, Rhodophyta) in the Mediterranean Sea. Cryptogam Algol 17:131–137
Aybulatov NA, Drushchits VA (1986) Bottom fauna as a dynamic factor in the redeposition of shelf sediments. Int Geol Rev 28:524–535
Báldi T (1984) The terminal Eocene and Early Oligocene events in Hungary and the separation of an anoxic, cold Paratethys. Eclogae Geol Helv 77:1–27
Barattolo F, Bassi D, Romano R (2007) Upper Eocene larger foraminiferal-coralline algal facies from the Klokova Mountain (Southern continental Greece). Facies 53:361–375
Bassi D (1998) Coralline algal facies and their palaeoenvironments in the Late Eocene of Northern Italy (Calcare di Nago). Facies 39:179–202
Bassi D (2005) Larger foraminiferal and coralline algal facies in an Upper Eocene storm-influenced, shallow water carbonate platform (Colli Berici, north-eastern Italy). Palaeogeogr Palaeoclimatol Palaeoecol 226:17–35
Bassi D, Nebelsick JH (2000) Calcareous algae from the Lower Oligocene Gornji Grad limestones of Northern Slovenia. Riv Ital Paleont Stratigr 106:99–122
Bassi D, Nebelsick JH (2010) Components, facies and ramps: redefining Upper Oligocene shallow water carbonates using coralline red algae and larger foraminifera (Venetian area, northeast Italy). Palaeogeogr Palaeoclimatol Palaeoecol 295:258–280
Bassi D, Kołodziej B, Machaniec E, Polak A (2000) Coralline algal limestones and rhodoliths from olisthostromes of the Krosno Beds (Oligocene, Polish Flysch Carpathians)—preliminary results. Slo Geol Mag 6:307
Bassi D, Braga JC, Zakrevskaya E, Radionova EP (2005) Re-assessment of the type collections of corallinalean genera (Corallinales, Rhodophyta) described by Maslov (1935–1962). Palaeontology 48:1–17
Bassi D, Carannante G, Murru M, Simone L, Toscano F (2006) Rhodalgal/bryomol assemblages in temperate type carbonate, channelised depositional systems: the Early Miocene of the Sarcidano area (Sardinia, Italy). In: Pedley HM, Carannante G (eds) Cool-water carbonates: depositional systems and palaeoenvironmental control. Geol Soc Lond Spec Publ 255:35–52
Bassi D, Nebelsick JH, Checconi A, Hohenegger J, Iryu Y (2009) Present-day and fossil rhodolith pavements compared: their potential for analysing shallow-water carbonate deposits. Sediment Geol 214:74–84
Bassi D, Carannante G, Checconi A, Simone L, Vigorito M (2010) Sedimentological and palaeoecological integrated analysis of a Miocene canalized coralline red algal carbonate margin (Matese Mountains, Central-Southern Apennines, Italy). Sediment Geol 230:105–122
Bassi D, Humblet M, Iryu Y (2011) Recent ichonocoenosis in deep-water macroids, Ryukyu Islands, Japan. Palaios 26:232–238
Bieda F (1962) Sur un faciès calcaire dans l’Éocène supérieur du Flysch des Karpates Polonaises. Ann Soc Geol Pol 32:401–410 (in Polish with French summary)
Bieda F (1968) Formation nummulitique dans le Flysch des Karpates Occidentales. Ann Soc Geol Pol 38:233–262 (in Polish with French summary)
Bieda F, Geroch S, Koszarski L, Książkiewicz M, Żytko K (1963) Stratigraphie des Karpates externes polonaises. Biul Inst Geol 181:1–174
Bignot G, Larsonneur C (1969) Etude du Cretace superieur au large du Cotentin et remarques sur les Planorbulina du Cretace superieur et du Paleocene. Rev Micropaléont 12:25–39
Birkenmajer K, Gucik S, Jasionowicz J, Koszarski L, Książkiewicz M, Nowak W, Sikora W, Szymakowska F, Ślączka A, Żgiet J, Żytko K (1962) Facies map of the lower Eocene in the Polish External Carpathians. In: Książkiewicz M (ed) Geological Atlas of Poland. Stratigraphic and facial problems, Fascicle 13—Cretaceous and Early Tertiary in the Polish External Carpathians, Instytut Geologiczny, Warszawa
Borowitzka MA, Larkum AWD (1986) Reef algae. Oceanus 29:49–54
Bosence DWJ (1983a) Description and classification of rhodoliths (rhodoids, rhodolites). In: Peryt TM (ed) Coated grains. Springer, Berlin, pp 217–224
Bosence DWJ (1983b) The occurrence and ecology of recent rhodoliths—a review. In: Peryt TM (ed) Coated grains. Springer, Berlin, pp 225–242
Bosence DWJ (1991) Coralline algae: mineralization, taxonomy, and palaeoecology. In: Riding R (ed) Calcareous algae and stromatolites. Springer, Berlin, pp 98–113
Brachert T, Forst MH, Pais JJ, Legoinha P, Reijmer JJG (2003) Lowstand carbonates, highstand sandstones? Sediment Geol 155:1–12
Braga JC (2003) Application of botanical taxonomy to fossil coralline algae (Corallinales, Rhodophyta). Acta Micropalaeontol Sin 20:47–56
Braga JC, Aguirre J (1995) Taxonomy of fossil coralline algal species: Neogene Lithophylloideae (Rhodophyta, Corallinaceae) from Southern Spain. Rev Palaeobot Palynol 86:265–285
Braga JC, Aguirre J (2001) Coralline algal assemblages in upper Neogene reef and temperate carbonates in Southern Spain. Palaeogeogr Palaeoclimatol Palaeoecol 175:27–41
Braga JC, Aguirre J (2004) Coralline algae indicate Pleistocene evolution from deep, open platform to outer barrier reef environments in the northern Great Barrier Reef margin. Coral Reefs 23:547–558
Braga JC, Bassi D (2007) Neogene history of Sporolithon Heydrich (Corallinales, Rhodophyta) in the Mediterranean region. Palaeogeogr Palaeoclimatol Palaeoecol 243:189–203
Braga JC, Martín JM (1988) Neogene coralline-algal growth-forms and their palaeoenvironments in the Almanzora River Valley (Almería, SE Spain). Palaeogeogr Palaeoclimatol Palaeoecol 67:285–303
Braga JC, Bosence DW, Steneck RS (1993) New anatomical characters in fossil coralline algae and their taxonomic implications. Palaeontology 36:535–547
Braga JC, Aguirre J, Esteban J (2009) Calcareous algae of Cabo de Gata–Níjar Nature 1009 Park. Field guide, ACUAMED-Consejeria Medio Ambiente. Junta de Andalucia, 1010 Madrid (Spanish text 143 pp, English text 63 pp)
Braga JC, Bassi D, Piller W (2010) Palaeoenviromental significance of Oligocene–Miocene coralline red algae—a review. In: Mutti M, Piller WE, Betzler C (eds) Carbonate systems during the Oligocene–Miocene climatic transition: Int Assoc Sediment Spec Publ 42:165–182
Brandano M, Frezza V, Tomassetti L, Pedley M, Matteucci R (2009) Facies analysis and palaeoenvironmental interpretation of the Late Oligocene Attard Member (Lower Coralline Limestone Formation), Malta. Sedimentology 56:1138–1158
Bressan G, Babbini L (2003) Corallinales del Mar Mediterraneo: guida alla determinazione. Biol Mar Mediterr 10:1–237
Bromley RG (1992) Bioerosion: eating rocks for fun and profit. In: Maples CG, West RR (eds) Trace fossils: short courses in paleontology 5:121–129
Bromley RG (1994) The palaeoecology of bioerosion. In: Donovan SK (ed) The palaeobiology of trace fossils. Wiley, Chichester, pp 134–154
Catuneanu O (2006) Principles of sequence stratigraphy. Elsevier, Amsterdam 375 p
Checconi A, Bassi D, Monaco P, Carannante G (2010) Re-deposited rhodoliths in the Middle Miocene hemipelagic deposits of Vitulano (Southern Apennines, Italy): coralline assemblage characterization and related trace fossils. Sediment Geol 225:50–66
Cieszkowski M, Golonka J, Rajchel J, Ślączka A (2003) Paleogene organogenic and organodetrital limestones as an indication of the tectonic evolution of the Western Outer Carpathians. 22nd IAS meeting of sedimentology, Opatija 2003. Abstracts Book, p 31
Cieszkowski M, Golonka J, Malata E, Olszewska B, Rajchel J, Ślączka A (2005) Wystąpienia wapieni organogenicznych, organodetrytycznych oraz innych skał węglanowych w Karpatach zewnętrznych. In: Cieszkowski M, Golonka J (eds) Wapienie organogenizne i organodetrytyczne w Karpatach zewnętrznych i ich znaczenie dla rekonstrukcji paleogeograficznych Tetydy. Seminarium naukowe, 21 kwietnia 2005, Kraków, pp 5–8
Dabrio CJ, Polo MD (1988) Late Neogene fan deltas and associated coral reefs in the Almanzora Basin, Almeria Province, southeastern Spain. In: Nemec W, Steel RJ (eds) Fan deltas—sedimentology and tectonic settings. Blackie, London, pp 354–367
Dercourt J, Gaetani M, Vrielynck B, Barrier E, Biju-Duval B, Brunet MF, Cadet JP, Crasquin S, Sandulescu M (2000) Atlas Peri-Tethys, Palaeogeographical Maps. CCGM/CGMW, Paris, p 269
Dieni I, Massari F, Medus J (2008) Age, depositional environment and stratigraphic value of the Cuccuru’e Flores Conglomerate: insight into the Palaeogene to Early Miocene geodynamic evolution of Sardinia. Bull Soc Géol Fr 179:51–72
Dimitrova E (2003) First data about the presence of genera Planorbulina and Sulcoperculina in the Upper Cretaceous and Paleogene in Bulgaria. Rev Bulg Geol Soc 64:55–57
Dutton SP (1982) Pennsylvanian fan-delta and carbonate deposition, Mobeetie Field, Texas Panhandle. AAPG Bull 66:389–407
Dziadzio PS, Enfield MA, Watkinson MP, Porębski SJ (2005) The Ciezkowice sandstone: examples of basin-floor fan-stacking patterns from the main (upper Paleocene to Eocene) reservoir in the Polish Carpathians. In: Golonka J, Picha FJ (eds) The Carpathians and their foreland: geology and hydrocarbon resources. Am Assoc Petrol Geol Mem 84:477–496
Flamand B, Cabioch G, Payri C, Pelletier B (2008) Nature and biological composition of the New Caledonian outer barrier reef slopes. Mar Geol 250:157–179
Foster MS (2001) Rhodoliths: between rocks and soft places. J Phycol 87:659–667
Geroch S, Nowak W (1984) Proposal of zonation for the late Tithonian-late Eocene, based upon arenaceous foraminifera from the outer Carpathians, Poland. In: Oertli HJ (ed) Benthos ‘83, 2nd international symposium on Benthic Foraminifera, (Pau 1983). Elf Aquitaine, Esso REP, Total CFP, Pau and Bordeaux, pp 225–239
Golonka J (1974) Glony wapienne paleogenu Krościenka (jednostka magurska) i Złatnego (pieniński pas skałkowy). Geol Quart 18:391–405
Golonka J, Cieszkowski M, Rajchel J, Ślączka A (2003) Paleogeography of the Paleogene carbonate platforms within the Northern Outer Carpathian realm. 22nd IAS meeting of sedimentology, Opatija 2003. Abstracts Book, p 66
Golonka J, Cieszkowski M, Krobicki M, Matyszkiewicz J, Olszewska B, Oszczypko N, Rajchel J (2005) Paleogeography of the Outer Carpathian carbonate platforms in Poland. Geolines 19:41–42
Haq BU, Hardenbol J, Vail P (1988) Mesozoic and Cenozoic chronostratigraphy and cycles of sea-level change. In: Wilgus CK, Hastings BS, Posamentier H, van Wagoner J, Ross CA, St Kendall CG (eds) Sea-level changes: an integrated approach. SEPM Spec Publ 42:71–108
İnan N (2003) Cocoarota orali n. sp. (Foraminifera) from Upper Maastrichtian-Lower Lutetian deposits of Turkey. Micropaleontology 49:201–204
İnan N, İnan S (2008) Selandian (Upper Paleocene) benthic foraminiferal assemblages and their stratigraphic ranges in the northeastern part of Turkey. Yerbilimleri 29:147–158
Iryu Y, Bassi D, Woelkerling W (2009) Re-assessment of the type collections of fourteen corallinalean species (Corallinales, Rhodophyta) described by W. Ishijima (1942–1960). Palaeontology 52:401–427
Ito M (2008) Downfan transformations from turbidity currents to debris flows at a channel-to-lobe transitional zone: the lower Plaeistocene Otadai Formation, Boso Peninsula, Japan. J Sediment Res 78:668–682
Janbu NE, Nemec W, Kırman E, Özaksoy V (2007) Facies anatomy of a channelized sand-rich turbiditic system: the Eocene Kusuri formation in the Sinop Basin, north-central Turkey. In: Nichols G, Paola C, Williams EA (eds) Sedimentary environments, processes and basins—a tribute to Peter Friend. Int Assoc Sediment Spec Publ 38:457–517
Johansen HW (1981) Coralline algae, a first synthesis. CRC Press, Boca Raton (239 pp)
Johnson JH (1963) The genus Archaeolithothamnium and its fossil representatives. J Paleontol 37:175–211
Kaminski MA, Gradstein FM (2005) Atlas of Paleogene cosmopolitan deep-water Foraminifera. Grzybowski Found Spec Publ 10:1–547
Köhler E, Buček S (2005) Paleocene reef limestones near Veľký Lipník (Pieniny Mts, NE Slovakia): facial environment and biogenic components. Slo Geol Mag 11:249–267
Köhler E, Salaj J, Buček S (1993) Paleogeographical development of the Myjava sedimentary area (western Slovakia) during the existence of the Paleocene reef complex. Geol Carpathica 44:373–380
Kołodziej B, Stolarski J (2000) Paleoceńskie koralowce z “iłów babickich”. In: XVII Konferencja Paleontologów, Kraków, 21–23 Sept 2000, pp 46–47
Koszarski L (1985) General outline of geology of Central part of Carpathian Foredeep and some essential data on the nappes of Polish Outer Carpathians. In: Koszarski L (ed) Geology of the middle Carpathians and the Carpathian Foredeep. Guide to excursion 3, Carpatho-Balkan geological association 13th congress, Cracow, Poland, 1985, Wydawnictwa Geologiczne, Warszawa, pp 11–39
Koszarski L, Kuciński T (1966) Szczegółowa Mapa Geologiczna Polski (bez utworów czwartorzędowych). Arkusz M34–78B Wojnicz—wydanie tymczasowe. Instytut Geologiczny, Warszawa
Koszarski L, Żytko K (1965) Le problème de la profondeur de la mer du géosynclinal karpatique de flysch. In: Reports, Carptho-Balkan Geol Assoc 7° Congress, part 2, vol 2, Sofia, pp 81–86
Krobicki M, Golonka J, Kołodziej B, Olszewska B, Oszczypko N, Słomka T, Tragelehn H, Wieczorek J (2004) Paleoceńskie olistolity wapieni koralowo-glonowych rejonu Haligowiec (pieniński pas skałkowy, Słowacja). Seminarium Egzotyki Karpackie—znaczenie w rekonstrukcjach paleogeograficzno-geotektonicznych, Kraków, 13 Dec 2004, pp 53–55
Książkiewicz M (1951) Objaśnienia do arkusza Wadowice. Wydawnictwa Państwowego Instytutu Geologicznego, pp 3–283
Książkiewicz M (1956) Geology of the northern Carpathians. Geol Rundsch 45:369–411
Książkiewicz M (ed) (1962) Geological Atlas of Poland. Stratigraphic and facial problems. Issue 13, Cretaceous and Early Tertiary in the Polish External Carpathians. Instytut Geologiczny, Warszawa
Książkiewicz M (1972) Budowa geologiczna Polski, tom IV, Tektonika, część 3, Karpaty. Wydawnictwa Geologiczne, Warszawa, pp 1–228
Książkiewicz M (1975) Bathymetry of the Carpathian Flysch Basin. Acta Geol Pol 25:309–367
Książkiewicz M (1977) The tectonics of the Carpathians. In: Pożaryski W (ed) Geology of Poland, IV: tectonics. Wydawnictwa Geologiczne, Warsaw, pp 476–620
Kuhnt W, Kaminski M (1989) Upper Cretaceous deep-water agglutinated benthic foraminiferal assemblages from the Western Mediterranean and adjacent areas. In: Wiedmann J (ed) Cretaceous of the Western Tethys. Proceedings of the 3rd international Cretaceous symposium, Tübingen 1987, pp 91–120
Leinfelder RR (1993) A sequence stratigraphic approach to the Upper Jurassic mixed carbonate-siliciclastic succession of the central Lusitanian Basin, Portugal. Profil 5:119–140
Leśniak T, Cieszkowski M, Golonka J, Waśkowska-Oliwa A (2005) Pozycja geologiczna piaskowców z Czerwina w jednostce podśląskiej polskich Karpat fliszowych. In: Cieszkowski M, Golonka J (eds) Wapienie organogeniczne i organodetrytyczne w Karpatach zewnętrznych i ich znaczenie dla rekonstrukcji paleogeograficznych Tetydy. Seminarium naukowe, 21 kwietnia 2005, Kraków, pp 13–18
Leszczyński S (1978) Algal limestones and rhodolites from Ciężkowice sandstone of the Silesian Unit, Polish Carpathians. Ann Soc Geol Pol 25:150–158 (in Polish with English summary)
Leszczyński S (1981) Ciężkowice sandstone of the Silesian Unit in Polish Carpathians: a study of coarse-clastic sedimentation in deep water. Ann Soc Geol Pol 51:435–502 (in Polish with English summary)
Leszczyński S, Kołodziej B (2004) Silikoklastyczno–węglanowe osady we fliszu Karpatach zewnętrznych: przykład z warstw istebniańskich w Melsztynie (płaszczowina śląska, paleocen?). Polska Konferencja sedymentologiczna, 8° Krajowe Spotkanie Sedymentologów Zakopane, 21–24 June 2004. Materiały konferencyjne: Przewodnik sesji terenowych, streszczenia referatów i posterów, p 104
Leszczyński S, Kołodziej B, Bassi D (2005) Siliciclastic-carbonate deposits in the flysch of the Outer Carpathians: a case study from the Istebna Beds in Melsztyn (Silesian Nappe, Paleocene), vol 1. Ann Univ Ferrara, Mus Sci Nat, 5th Reg Symp Int Fossil Algae Ass, 30–31 Aug 2005, Ferrara, p 30
Lowe DR (1982) Sediment gravity flows: II. Depositional models with special reference to the deposits of high-density turbidity currents. J Sediment Petrol 52:279–297
Lund M, Davies PJ, Braga JC (2000) Coralline algal nodules off Fraser Island, eastern Australia. Facies 42:25–34
Malata E (2002) Albian-Early Miocene foraminiferal assemblages of the Magura Nappe (Polish Outer Carpathians). Geol Carpathica 53(special issue):77–79
Małecki J (1956) The calcareous algae of the Tatra Eocene. Ann Soc Geol Pol 25:135–158 (in Polish with English summary)
Marrack EC (1999) The relationship between water motion and living rhodolith beds in the southwestern Gulf of California, Mexico. Palaios 14:159–171
Minnery GA (1990) Crustose coralline algae from the Flower Garden Banks, northwestern Gulf of Mexico: controls on distribution and growth morphology. J Sediment Petrol 60:992–1007
Monstad S (2000) Carbonate sedimentation on inactive fan-delta lobes: response to sea-level changes, Sant Llorenç del Munt fan-delta complex, NE Spain. Sediment Geol 138:99–125
Nalin R, Nelson CS, Basso D, Massari F (2008) Rhodolith-bearing limestones as transgressive marker beds: fossil and modern examples from North Island, New Zealand. Sedimentology 55:249–274
Nalin R, Ghinassi M, Basso D (2010) Onset of temperate carbonate sedimentation during transgression in a low-energy siliciclastic embayment (Pliocene of the Val d’Orcia Basin, Tuscany, Italy). Facies 56:353–368
Nebelsick J, Bassi D (2000) Diversity, growth-forms and taphonomy: key factors controlling the fabric of coralline algal dominated shelf carbonates. In: Insalaco E, Skelton P, Palmer T (eds) Carbonate platform systems: components and interactions. Geol Soc Lond Spec Publ 178:89–107
Nebelsick JH, Bassi D, Drobne K (2000) Microfacies analysis and palaeoenvironmental interpretation of Lower Oligocene, shallow-water carbonates (Gornji Grad Beds, Slovenia). Facies 43:157–176
Nebelsick JH, Rasser M, Bassi D (2005) Facies dynamics in Eocene to Oligocene circumalpine carbonates. Facies 51:197–216
Olszewska B (1997) Foraminiferal biostratigraphy of the Polish Outer Carpathians: a record of basin geohistory. Ann Soc Geol Pol 67:325–337
Olszewska B (2009) Small foraminifera of “Nummulitic Eocene” of the Tatra Mts.—stratigraphical aspect and paleoenvironment. Prz Geol 57:703–713
Palanques A, Kenyon NH, Alonso B, Limonov A (1995) Erosional and depositional patterns in the Valencia Channel mouth: an example of a modern channel-lobe transition zone. Mar Geophys Res 17:503–517
Philip J (2003) Peri-Tethyan neritic carbonate areas: distribution through time and driving factors. Palaeogeogr Palaeoclimatol Palaeoecol 196:19–37
Puga-Bernabéu Á, Vonk AJ, Nelson CS, Kamp PJJ (2009) Mangarara Formation: exhumed remnants of a middle Miocene, temperate carbonate, submarine channel-fan system on the eastern margin of Taranaki Basin, New Zealand. N Z J Geol Geophys 52:73–93
Rajchel J, Myszkowska J (1998a) Litologia wapieni z warstwy wapienia litotamniowego z Birczy (wt)—jednostka skolska, zewnętrzne Karpaty fliszowe. Prz Geol 46:1247–1253
Rajchel J, Myszkowska J (1998b) Exotic clasts of organodetritic algal limestones from lithosomes of the Babica Clay, Skole Unit (Outer Flysch Carpathians, Poland). Ann Soc Geol Pol 68:225–235
Rasser MW (2000) Coralline red algal limestones of the Late Eocene Alpine Foreland Basin in Upper Austria: component analysis, facies and paleoecology. Facies 42:59–92
Rasser MW, Nebelsick JH (2003) Provenance analysis of Oligocene autochthonous and allochthonous coralline algae: a quantitative approach towards reconstructing transported assemblages. Palaeogeogr Palaeoclimatol Palaeoecol 201:89–111
Rasser MW, Piller WE (1999) Application of neontological taxonomic concepts to Late Eocene coralline algae (Rhodophyta) of the Austrian Molasse Zone. J Micropalaeontol 18:67–80
Rasser MW, Piller WE (2004) Crustose algal frameworks from the Eocene Alpine Foreland. Palaeogeogr Palaeoclimatol Palaeoecol 206:21–39
Rice ME, Macintyre IG (1982) Distribution of sipuncula in the coral reef community, Carrie Bow Cay, Belize. In: Rützler K, Macintyre IG (eds) The Atlantic barrier reef ecosystem at Carrie Bow Cay, Belize I: structure and communities. Smithson Contr Mar Sci 12:1–539
Ricou LE, Mercier de Lepinay B, Marcoux J (1986) Evolution of the Tethyan seaways and implications for the oceanic circulation around the Eocene–Oligocene boundary. In: Pomerol C, Premoli Silva I (eds) Terminal Eocene events. Dev Palaeont Stratigr 9:387–394
Rögl F (1999) Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geol Carpathica 50:339–349
Sahy D, Săsăran E, Tămaş T (2008) Microfacies analysis of Upper Eocene shallow-water carbonates from the Rodnei Mountains (N Romania). Studia Univ Babeş-Bolyai Geol 53:13–24
Samuel O, Borza K, Köhler E (1972) Microfauna and lithostratigraphy of the Paleogene and adjacent Cretaceous of the Middle Vah Valley (West Carpathians). GUDS, p 246
Samuel O, Köhler E, Borza K (1977) Haddonia praeheissigi and Miliola? andrusovi, two new species from Upper Senonian and Paleocene bioherm limestones of West Carpathians (Slovakia). Zapadne Karpaty, ser paleont 2–3, GUDS, pp 87–95
Sarg JF (1988) Carbonate sequence stratigraphy. In: Wilgus CK, Hastings BS, Kendall CG, Posamentier HW, Ross CA, van Wagoner JC (eds) Sea-level changes: an integrated approach. SEPM Spec Publ 42:155–181
Scheibner E (1968) Contribution to the knowledge of the Paleogene reef-complexes of the Myjava-Hričov-Haligovka zone (West Carpathians). Mitt Bayer Staatssamml Paläont Hist Geol 8:67–97
Scheibner C, Speijer RP (2008) Late Paleocene–Early Eocene Tethyan carbonate platform evolution—A response to long- and short-term paleoclimatic change. Earth Sci Rev 90:71–102
Schlagintweit F (2005) Cryptobiotic foraminifera from the Paleocene Kambühel Formation (Northern Calcareous Alps, Austria). Studia Univ Babes-Bolyai Geol 50:13–17
Stockar R (2000) Fossil coralline algae from the Paleocene Montorfano Member type-section (Tabiago Formation, northern Italy). Eclogae Geol Helv 93:409–427
Tucker ME, Wright VP (1990) Carbonate sedimentology. Blackwell, Oxford, p 482
Unrug R (1963) Istebna Beds—a fluxoturbidity formation in the Carpathian flysch. Ann Soc Geol Pol 33:49–92
Unrug R (1968) The Silesian Cordillera as the source of clastic material of the Flysch sandstone of the Beskid Slaski and Beskid Wysoki ranges (Polish Western Carpathians). Ann Soc Geol Pol 38:81–164 (in Polish with English summary)
Vecsei A, Sanders DGK (1997) Sea-level highstand and lowstand shedding related to shelf margin aggradation and emersion, Upper Eocene–Oligocene of Maiella carbonate platform, Italy. Sediment Geol 112:219–234
Vlahović I, Tišljar J, Velić I, Matičec D (2005) Evolution of the Adriatic Carbonate Platform: paleogeography, main events and depositional dynamics. Palaeogeogr Palaeoclimatol Palaeoecol 220:333–360
Woelkerling WJ (1988) The coralline red algae: An analysis of the genera and subfamilies of nongeniculate Corallinaceae. British Museum (Natural History)/Oxford University Press, London/Oxford, p 268
Woelkerling WJ, Irvine LM, Harvey AS (1993) Growth-forms in non-geniculate coralline red algae (Corallinales, Rhodophyta). Aust Syst Bot 6:277–293
Wood A, Smith AJ (1959) The sedimentation and sedimentary history of the Aberystwyth Grits (Upper Landdoverian). Quart J Geol Soc Lond 114:163–195
Wright P, Cherns L, Hodges P (2003) Missing molluscs: field testing taphonomic loss in the Mesozoic through early large-scale aragonite dissolution. Geology 31:211–214
Wynn RB, Kenyon NH, Masson DG, Stow DAV, Weaver PPE (2002) Characterization and recognition of deep-water channel–lobe transition zones. AAPG Bull 86:1441–1462
Acknowledgments
The research by the Polish authors was funded by the Jagiellonian University grants DS. The authors thank Barbara Olszewska (Kraków) for discussing the taxonomy of foraminifera and Pauline Harris (Norwich) for correcting the first version of the English text. The present version of the manuscript strongly benefited from thoughtful and constructive suggestions by Wojciech J. Nemec (Bergen). The reviewers Werner E. Piller (Graz) and Michael W. Rasser (Stuttgart) as well as the editor Franz Fürsich are acknowledged for critical remarks and useful comments on a previous version of this paper.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Leszczyński, S., Kołodziej, B., Bassi, D. et al. Origin and resedimentation of rhodoliths in the Late Paleocene flysch of the Polish Outer Carpathians. Facies 58, 367–387 (2012). https://doi.org/10.1007/s10347-012-0302-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-012-0302-8