
Abstract
This paper presents archaeobotanical and invertebrate evidence from the flotation and analyses of 46 archaeological contexts from six middens from the Samaca and Ullujaya basins, lower Ica Valley, south coast of Peru. This is part of one of the world’s driest deserts and organic remains can enjoy extraordinary preservation in its hyperarid climate. Each of these contexts represents snapshots with which to piece together a picture of changing human ecology in the lower Ica Valley over nearly two millennia, from Ocucaje Phases 3/4 of the Early Horizon (c. 750 b.c.); through to Early Nasca Phases 2/3 (c. a.d. 100–450); Late Nasca Phases 6/7 (c. a.d. 450–600) and Middle Horizon Epoch 2 (c. a.d. 900). They also offer proxy evidence of wider ecological changes in these basins. Read together with geoarchaeological and pollen data, the archaeobotanical data we present show a gradual intensification of agriculture from small-scale Early Ocucaje societies subsisting mainly on gathered marine and terrestrial resources, through to sophisticated irrigation agriculture by Nasca times, but culminating in a collapse of agricultural production and a return to the gathering of wild marine and plant resources much later, during the Middle Horizon. This trajectory of human ecology is consistent with the model presented elsewhere of a gradual removal of Prosopis dominated riparian woodland for the purpose of increasing agricultural production, which in time exposed the landscape of the lower Ica Valley to high-energy, episodic flood events and of one of the world’s strongest and most persistent wind regimes.
Similar content being viewed by others

Introduction
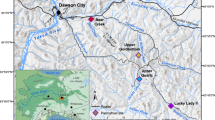
Typically, the intermittent rivers of the Peruvian desert coast emerge from the western slopes of the Andes to flow across a narrow coastal plain to the Pacific Ocean. The Río Ica on the south coast, however, has a very different geomorphological configuration. For its access to the sea is blocked by the uplifted sedimentary fore-arc basin of the Tablazo de Ica and consequently it is diverted southward to flow parallel to the coast for some 150 km through a series of wide basins which it has incised into that sedimentary desert tableland. These basins alternate with narrows and canyons where the river has encountered remnants of the hard and ancient igneous rock of the Andean basement.
Today settlement and agriculture is concentrated in the middle Ica Valley, lying between the Andean foothills and the Ocucaje Basin. Even in Ica itself the river is popularly imagined to end after Ocucaje, but this is far from the case. Downstream from Ocucaje, the Río Ica still has around 85 km to run until the Pacific through a series of basins: Callango, Ullujaya and Samaca, followed by a long canyon cut through the coastal cordillera (Fig. 1). Finally it enters a last wide basin, Montegrande, and thence the ocean.

The South Coast and the Lower Ica Valley (Landsat 7 etm + 2000)
These great basins of the lower valley present a striking contrast to the urban bustle and extensive agriculture about the modern town of Ica, for they are mostly desertified and almost empty of people and cultivation. Although there is some agriculture in Callango, and indeed recent efforts to rejuvenate Ullujaya and Samaca, these are exceptions to prove a rule; today, human settlement is concentrated in the middle Ica Valley (Fig. 1a). So it was during the 16th century when the Spanish founded the city of Ica there. Yet the archaeological record suggests that this was not always the case.
As Massey (1991, p. 317) observes, the middle Ica Valley was only intensively settled “during the Late Intermediate with the expansion of the Chirana irrigation canal from the upper valley to the zone of Tacaraca-Ica Viejo, the Ica-Inca capital in the upper middle valley” (Fig. 2). Prior to this, archaeological remains attest to the lower Ica Valley as being the site of most settlement and home to substantial prehistoric populations. Strong, for instance, made an early excursion down the valley in 1941, observing that:
“It is already apparent that in earlier times this part of the Ica valley was thickly populated. Today, owing to the almost complete diversion of the water of the Ica river in the upper part of its drainage area, the lower portions of the valley below Ocucaje are desolate and largely without habitants. Even the most cursory examination indicates that this was not always the case.” [Strong et al. 1943 p. 23]

It scarcely seems credible from their presently austere condition that, during the first millennium b.c., these basins of the lower Valley were likely the centre of human settlement in the Ica Valley (Fig. 1b). Yet, Callango is the location of, among others, two large habitation sites, known as Animas Bajas and Animas Altas, covering 50 and 100 ha, respectively (Massey 1991; Cook 1999; DeLeonardis 2005). And the Callango Basin is very large, with traces of once extant vegetation extending over some 3,600 ha. The Ullujaya and Samaca Basins are smaller but, if anything, show even greater ecological degradation. The lower Ica Valley thereby presents a prima facie case for dramatically changed ecological and landscape conditions. Our investigations seek to answer the questions of when, how and indeed why, such changes took place. These basins have a remarkable archaeological record, preserved precisely because of their subsequent abandonment (Fig. 3). As Rossel Castro (1977) p. 59, our translation) observes, “in Samaka and Ullujaya one may find all the strata of the civilisations of Ica. This little known region is of great importance to the field of archaeology”.

The Area of Investigation: the Samaca and Ullujaya Basins, lower Ica Valley
This paper describes the methods and reports the results of the archaeobotanical and invertebrate analyses of our investigations. Read together with geoarchaeological and other lines of data, the data we present here help to elaborate a picture of gradual landscape change in the lower Ica Valley—a case-study in anthropogenic geomorphology—from the Ocucaje Phases 3/4 of the Early Horizon (c. 750 b.c.) through to the Epoch 2 of the Middle Horizon (c. a.d. 900).
Materials and methods
Archaeological investigations in the Samaca and Ullujaya basins
The rubric for our investigations in the lower Ica Valley was that long since advocated by Butzer (1982) of ‘archaeology as human ecology’. Moreover, we aspire to tracking changes in human ecology over the passage of time—the perspective of historical ecology (Balée 1998). Since fields and canal systems are environments modified by humans, they are ‘anthropogenic’ in the just same sense as, say, ceramic fragments. Indeed, in our context here, entire degraded landscapes may well be artefacts. Our methodology, therefore, combines geoarchaeological and archaeobotanical approaches. Understanding geomorphological change is a requisite for interpreting the extant archaeological record and hence for understanding why the landscape itself looks like it does today. Archaeobotany, meanwhile, offers direct and indirect insights into human diet and proxy evidence of ecological change.
Our investigations focussed on two large areas of archaeological remains defined as H-13 and G-8/9 in the survey by Cook (1994) in the Samaca and Ullujaya basins, respectively (Fig. 3). Here we carried out high-resolution archaeological surveys by means of differential GPS and made excavations following the British convention of determining stratigraphic units and recording archaeological sediments, natural layers, lenses, finds and other elements according to those units (Molas 1994). Both G-8/9 and H-13 occupy the surfaces of relict river terraces, which define much of the landscape of these basins. Although there is some patchy vegetation on Ullujaya G-8/9 as a consequence of a canal cut during the 1950s in a failed enterprise to grow cotton (Fig. 3), otherwise these relict terraces are today entirely barren. Yet there is plenty of evidence that this was not always so, including, most conspicuously, almost four hundred relict huarango (Prosopis sp.) tree trunks recorded across their surfaces (Fig. 4; Cook 1999 p. 65; Beresford-Jones et al. 2009b, pp. 307–311). Geomorphological analyses reveal that within this desolate landscape are buried fragments of ancient, once organic soils (Beresford-Jones et al. 2009a). From these and other lines of data we determine that these terraces were abandoned as productive landscapes sometime during the Middle Horizon, since when they have suffered a long process of surface aeolian deflation.

Map of relict huarango tree trunks of the Samaca and Ullujaya Basins
This is because the region has an extraordinarily strong and unimodal wind regime (McCauley et al. 1977; Parker Gay 1999), with a continuous average wind speed measured over 9 months in the Samaca basin of 28.8 kph, and frequent gusts over 115 kph (Beresford-Jones et al. 2009a). The landscape itself here stands in testimony to the great strength and stability of this wind regime: wind deflation evident in a classic ‘inversion of topography’, whereby features such as canal courses, once cut into the land surface, are preserved today by calcite enrichment standing above their surrounding deflated relict terrace surfaces. Meanwhile, artefacts spanning some two millennia to the Middle Horizon, have been left as mixed assemblages scattered across the abandoned terrace surfaces (Beresford-Jones et al. 2009a, b).
So, unsurprisingly, our high resolution archaeological survey reveals Ullujaya G-8/9 and Samaca H-13 to be, not so much individual archaeological sites, but rather the now-faint traces of multiple occupations over considerable time depths. These traces are conglomerates of the remains of human settlement, made up of mounds; fragments of enclosures, platforms and buildings of adobe brick or stone; rubbish middens; ancient floors with post holes, hearths and ovens; cemeteries; and remnants of the ancient agricultural landscape in the form of relict canal courses (see Beresford-Jones et al. 2004; Beresford-Jones 2005; Beresford-Jones and Arce 2010; Beresford-Jones in press for further details).
Within this archaeological palimpsest, our excavations focussed on habitation sites preserved now only as low mounds of domestic rubbish with occasional adobe architecture middens—the rubbish of ancient settlement—for the record they offer of human ecology, not least of changing diet. They may also turn out to be proxy indicators of changing ecological conditions in the basins of the lower Ica Valley. We present here data from excavations of 46 archaeological contexts from six middens, dated by ceramic fragments and other material culture associated with them (Fig. 3; Table 1). Each of these contexts represent snapshots, of varying exposures, with which to piece together a picture of changing subsistence in the lower Ica Valley over nearly two millennia from c. 750 b.c. to a.d. 900 (Fig. 2).
Sampling, extraction and identification of remains
In the lower Ica Valley organic remains can enjoy extraordinary preservation if they are deposited above a fluctuating and alkaline water table. For here it virtually never rains so that desiccated plant materials can endure for centuries. Naturally however, any line of archaeological data—even one enjoying the remarkable preservation conditions of the lower Ica valley—offers only a fragmented and partial picture of the past. Moreover, middens are of course artefacts of human activity: anthropogenic assemblages that can offer only imperfect reflections of past environments and incomplete records of human diet. Such weaknesses may be mitigated by careful selection of excavation contexts and sampling strategies. More importantly, however, interpretations based several ‘multi-proxy’ lines of data will always be more secure than those offered by any one single dataset. Consequently, the interpretations of the archaeobotanical and invertebrate data presented here will be drawn against other lines of inference, including geoarchaeological, pollen and other archaeological data and, not least, a thorough understanding of today’s riparian ecology (Whaley et al. 2010, 2011), which we shall set out as we go along.
Flotation, using a simplified version of the standard Cambridge Flotation Tank in the field, proved an extremely effective way of extracting desiccated organic remains from the dry and friable sediments of the lower Ica Valley. Samples of uniform, eight-litre volumes from selected archaeological deposits were floated and their contents separated into 0.5 and 2.0 mm light fractions using a sieve deck, and a residual 2.0 mm heavy fraction. Most desiccated or charred plant remains and occasional smaller terrestrial molluscs were sorted from light fractions, while most marine and terrestrial molluscs, crustaceans and echinoderm remains and a few charcoals with mineral impregnations were found in heavy fractions. These were subsequently sorted and identified in the Museo de Historia Natural de la Universidad Ricardo Palma, Lima and in the Pitt-Rivers Bioarchaeology Laboratory, Cambridge using modern reference collections from the area, including one housed in the Royal Botanical Gardens, Kew (Whaley et al. 2010, 2011) and available literature (Martin and Barkley 1961; Sagástegui Alva 1973; Brako and Zarucchi 1993; Brack A 1999). Archaeobotanical identifications were made by Alarcón, Beresford-Jones and Whaley with contributions by Cadwallader and Oscar Portocarrero. Invertebrate identifications were made by Cadwallader and Manuel Gorriti. Sediments for pollen analysis were prepared in the laboratory following Berglund and Ralska-Jasiewiczowa (1986) and identifications made by Alex Chepstow-Lusty.
Results
Table 2 (see also ESM) summarises the archaeobotanical data from these contexts in the lower Ica Valley.
The table shows 46 archaeological contexts analysed, totalling almost 400 l of samples. The remains analysed from these contexts have been grouped according to their ecological associations (Galan de Mera 1999; Whaley et al. 2010, 2011):
-
a.
Trees of the riparian woodland, dominated by huarango
-
b.
Shrubs and herbs of riparian woodland and woodland edges
-
c.
Agrestal weeds associated with areas of agriculture and other human disturbance
-
d.
Plants of water courses and marshy areas
-
e.
Grasses of open areas
-
f.
Cultivated plants of the agricultural landscape
-
g.
Wood charcoals, charred parenchyma remains and other flowering plant fragments that could not be further identified.
Table 3, meanwhile, shows the marine and terrestrial mollusc and other marine foods recovered from the heavy fractions of the products of flotation of selected lower Ica Valley archaeological contexts. We shall refer to all of these here as ‘invertebrate’ data: most are molluscs, but crustaceans and sea urchins of course are not.
All this data comes from flotation samples of a uniform size and are therefore in a broad sense comparable. Yet we will resist what Hubbard and Clapham (1992) p. 117) call the “Pavlovian urge to quantify”, for as they rightly observe, quantification beyond that justified by context or taphonomy “degenerates into nonsense”. Plant materials in Table 2 and invertebrate remains in Table 3 are therefore classified according to their occurrence in the assemblages of the different archaeological contexts as: ‘some’ (less than 10 items); ‘moderate’ (between 10 and 100); and, ‘abundant’ (more than 100): broad categories that smooth out some distortions which may arise from sampling contexts with different taphonomic histories. Heavy fraction materials are also shown since these provide a measure of the relative density of these various midden contexts, which are useful when comparing contexts across time.
Discussion
Arranged by time along one axis and by major ecological group down the other, the archaeobotanical data in Table 2 show patterns, which, considered together with other lines of evidence, including invertebrate data (Table 3), geoarchaeological data (Beresford-Jones et al. 2009a), pollen evidence and, not least, the archaeological record in general (Beresford-Jones et al. 2009b; Beresford-Jones in press), suggest a gradual story of change in human ecology over nearly two millennia as follows.
Early Ocucaje—gathered wild resources
The contexts analysed dating to Ocucaje phases 3/4 of the Early Horizon (c. 750 b.c.) are probably among the oldest midden contexts yet scientifically analysed in the Ica Valley (see also DeLeonardis 1997, pp. 588–589, Table 8.4, although these data pertain to later Ocucaje phases). They contain no obvious plant domesticates (Group F in Table 2), other than seeds and fibre of cotton (Gossypium sp.). These remains are inferred to be of Gossypium barbadense based upon biogeography (Westengen et al. 2005), for only rarely are sufficient morphological characters preserved in archaeological specimens to allow for identification between species (Stephens 1970).
Cotton is one of the first plants recorded in the earliest sedentary human occupations on the coast of Peru, used for the lines and nets to harvest its bountiful marine resources (Moseley 1975; Dillehay et al. 2007). Over the long unfolding of the period that some call the ‘Cotton Pre-ceramic’ the fabric technologies of fishing evolved into weaving. Much later, by the time of the Early Horizon, all of the structural innovations that characterise the extraordinary Andean textile tradition had been developed (Conklin 2008). And cotton textiles are some of the most famous examples of that Chavín material culture, to which the mound Feature 21 contexts so clearly relate (Beresford-Jones et al. 2011). Although few Chavín textiles have been scientifically excavated, some are reportedly from Samaca, Ullujaya and Callango in the lower Ica Valley (Burger 1992, p. 198; Roe 2008). And the cotton plant itself is often prominent in the iconography of some of these textiles (Conklin 2008, p. 265).
Cotton being the only identifiable domesticated plant in these Early Horizon contexts begs the question of what these people were eating, to which the answer, it seems, was predominately foods gathered from the sea, lomas and riparian woodland. The Ullujaya basin perfectly fits Kroeber’s (1944, p. 24) characterisation of the region as “coastal but not maritime”, because it is separated from the Pacific Ocean by the Lomas de Ullujaya (Fig. 1). Part of the Coastal Cordillera, these rise to almost a thousand metres blocking ocean mists which are thereby deposited on their flanks supporting lomas vegetation and a rare coastal population of wild guanaco (Lama guanacoe). The ocean does not feel close. Indeed, it is a long walk of at least 2 days to reach the mouth of the Ica River by following its course south through the hot and contorted canyon of its lower reaches. Yet, walking west across the aptly named Pampa Correviento over a 600 m pass in the lomas, one may reach the vast Playa Lomitas in only 8 h. And these Chavín contexts contain abundant vestiges of such journeys.
Since marine foods seem to make up a huge proportion of the total Early Ocucaje midden these contexts are almost one order of magnitude richer in heavy marine and terrestrial remains than those same volume samples recorded from other, later archaeological contexts (Table 3). These dense middens are dominated by sea urchin (Tetrapygus niger) remains, and by the shells of three types of small marine mussels (Table 3; Fig. 5), although some of the very smallest mussel shells in these contexts may have been collected and thereby deposited inadvertently along with larger molluscs.

Selected marine foods recovered from the Early Horizon midden context (feature 21/4, S.U. 7027) of mound 21, Ullujaya G-8/9; a Sea urchin spines (Tetrapygus niger), b small mussel shells (Brachidontes sp.), c charred plant parenchyma fragments
They also contain large numbers of land snail shells (Bostryx sp., Bulimulidae). Several species of Bostryx have a number of morphological and physiological adaptations that allow them to pass long periods in stasis and thereby survive in the ephemeral fog meadow environment of the lomas. Today they are found living exclusively in such seasonal lomas ecologies (Ramírez et al. 2003) and never in the riparian ecology of the valley basins. In some locations on the south coast of Peru, today barren of vegetation, the presence of these molluscs has been taken as evidence of past climatic change (e.g. Mächtle et al. 2010). In the specific contexts of the basins of the lower Ica Valley, however, the pattern of these terrestrial mollusc deposits—ubiquitous in midden contexts of all archaeological time periods (Table 3), but confined to them and never found in off-site contexts—do not suggest to us good evidence that lomas conditions and ecology once extended down into these basins. Nor is there any archaeobotanical, geoarchaeological or other evidence here for such a radical change in past atmospheric humidity and climate. Rather, there are powerful geomorphological and other indications that, beyond the reach of the river, the region has been unchangingly dry for time periods far deeper than the archaeological ones of interest to us here (Beresford-Jones et al. 2009a), not least the very preservation of the desiccated organic remains discussed in this paper. Consequently we interpret the land snail remains in the middens of the lower Ica Valley as food refuse: anthropogenic assemblages gathered in the lomas some 4 h walk away. Moreover, evidence in huge land shell mounds in the Lomas de Ullujaya attest to precisely that activity (Fig. 6).

Land snail mounds in the Lomas de Ullujaya; a huge mounds of land snails (Bostryx sp., Bulimulidae), b live Bostryx sp., c Obsidian point from land snail shell mound, of precisely the dimensions to extract flesh
The range of invertebrate species in these Early Ocucaje contexts is more limited than those of later periods. In particular, the Early Horizon occupants seemed to have had a preference for smaller, lighter kinds of shellfish. There may be many reasons for that choice, but ease of foraging and transport—perhaps without the aid of llama caravans—would seem likely to be among them. For while remains of camelids in the form of bones and fibres in textiles become common in the middens of the lower Ica Valley in Late Ocucaje (phase 10) and thereafter, they are absent from these Early Ocucaje contexts.
Turning to the other remains in these Early Ocucaje contexts shown in Table 2, there is a dearth of identifiable domesticated plants compared with other, later archaeological contexts. There are also no grasses which might indicate open areas (Group E), very few weeds associated with human disturbance or agriculture (Group C), and few shrubs and larger herbs that grow in clearances and along the edges of the south coast riparian woodland (Group B).
There are copious charcoal fragments and some charred remains of parenchyma which indicate that some plant foods were almost certainly being consumed. In principle such charred parenchymous materials can be identified using the guidelines set forth by Hather (1993). No published reference materials exist yet for the south coast of Peru, however, and, as Hather (2000) puts it, “the certain identification of some fragments as parenchyma and, in almost all cases, the precise identification of this category of material, requires the use of SEM [Scanning Electron Microscopy]”. We have not attempted here to identify plant parenchyma. There are however many desiccated huarango leaf litter remains (poña), charred leaflets and indeed the stony endocarp of its seeds, which in these contexts might suggest that the soft, sweet mesocarp of Prosopis pods was part of human diet and is conceivably the source of some of this parenchymous material. Other possible sources could include the starchy rhizomes of various genera of Cyperaceae which are recorded as providing food in New World Deserts (Holden 1991; Rea 1997; Hodgson 2001). Cyperaceae remains are also recorded in these Early Ocucaje contexts as well as in contexts from later time periods (Table 2).
Nasca—agricultural intensification
Table 2 also shows that the middens of the subsequent Early Nasca period were seemingly empty of obvious plant domesticates, bar cotton. This is, however, likely to be an artefact of sampling, because the relative density of the heavy fraction component of these Early Nasca middens (Table 2) show them to be extremely sparse relative to those of other time periods. And other lines of data strongly suggest a fully developed agricultural landscape in these basins by the Early Intermediate, the most obvious of which are the many fragments of relict canals across the surface of H-13 and G-8/9. The precise dating of features such as canal systems, in use over extended time-periods, is problematic. Nonetheless, the evidence from excavation of several of these relict canals, including some with Late Nasca platforms constructed across their courses, is that they are all fragments of the same, contemporary system and further, that they date terminus ante quem to the end of the Early Nasca period (Beresford-Jones et al. 2009b; Beresford-Jones in press).
Compared to preceding Early Ocucaje contexts, these Early Nasca middens, despite their relative scarcity, nonetheless included significantly more remnants of grasses (Group E) and woody shrubs like Galvezia sp. and Tessaria/Pluchea, typical of open areas of mixed riparian vegetation (Group B). They also had considerably more remains of weedy herbaceous plants (Group C), than those earlier remains.
This latter group included chenopods and amaranths (sometimes classed together here where they are difficult to distinguish) two genera that include many halophytes (plants adapted to estuarine or otherwise saline environments) and are the classic indicators of the clearances and ecological disturbances that come with human activities such as agriculture. These and the other weedy species of Group C are found growing abundantly today amid and along the edges of the modern fields of Huerto Huamaní in Samaca.
Like the Early Ocucaje middens, these Early Nasca contexts contained seeds of the shrub, Indigofera aff. truxillensis called ‘tintillo’ locally (Whaley et al. 2010). This is a plant from which an intense blue dye may be obtained from the leaves, through a particularly obscure, unpredictable and complex process (Stone-Miller 1992, pp. 17, 20). Early Nasca textiles are remarkable among the vast corpus of later, pre-Hispanic Andean textiles for their extravagant use of the colour blue (Stone-Miller 1992, pp. 165–167): often for the entirety of a large plain weave textile then fringed with the most intricate polychrome needle-knit borders depicting flowering plants and hummingbirds (Stone-Miller 1992, pp. 72–91). These ancient seeds hint also at ecological change, for, not only does blue become ever rarer in later period textiles, but also, in our later archaeological contexts, there are no Indigofera remains. Indigofera thrives in the shade, such as that found in riparian woodland, or along flood channels. And, while it is still present, albeit uncommon, elsewhere on the south coast (Whaley et al. 2010), we have yet to encounter a single plant growing in the lower Ica Valley today.
Again, these deposits also included moderate numbers of huarango leaflets and leaf rachis fragments. Indeed, macrofossil remains of huarango were the single common denominator of all the archaeological contexts analysed in G-8/9 and H-13. Morphological features, including leaflet shape and size, an absence of thorns, pod size and shape, and number of interfoliar glands per rachis, allow these to be identified as P. pallida forma pallida (following Harris et al. 2003), or P. limensis (as preferred here, following Mom et al. 2002; Burghardt et al. 2010).
Table 2 shows that Late Nasca midden contexts contain the full suite of south coast plant domesticates (Group F), including cotton and gourd (Lagenaria siceraria), together with the domesticated food plants of maize (Fig. 7), beans (Phaseolus lunatus and Phaseolus vulgaris), pumpkin (Cucurbita sp.), peanut (Arachis hypogaea), achira (Canna edulis), the fruit trees pacay (Inga feuillei) and guava (Psidium guajava) and chilli seeds (Capsicum sp.).

Selected examples of the most common plant finds from excavations in the Samaca and Ullujaya Basins; maize cobs (Zea mays) from Late Nasca midden contexts in mound feature 1, Ullujaya G-8/9: a S.U. 3010, b S.U. 3012; from middle horizon midden contexts S.U. 32 in mound feature 23, Samaca H-13: c huarango pods (Prosopis limensis), d cotton seeds (Gossypium sp.)
The presence of achira, pacay and guava in this assemblage, just like that of Indigofera, are each noteworthy for their possible implications for ecological change. The wild ancestor of achira comes from the subtropical forests on the eastern side of the Andes (Gade 1966; Ugent et al. 1984) and, on the south coast, it can thrive only in the most sheltered, shady and humid conditions, most often alongside irrigation canals, because its large delicate leaves will not tolerate wind, nor exposure to full sun. Pacay too appears have been domesticated from an Amazonian ancestor (Inga striata and/or Inga sapindoides, Pennington 1997; Pennington and Fernandes 1998). Today, both pacay and guava are grown in the middle Ica Valley as part of a traditional huerta (mixed orchard and kitchen garden) agricultural system (Whaley et al. 2010, 2011). Our archaeobotanical evidence is consistent with the botanical understanding that these are cultigens here, introduced by humans and not occurring naturally in the riparian woodland of the valley. For they are absent from the Early Ocucaje contexts we have analysed from the lower Ica Valley. Pacay leaves and pods are visible in looted contexts dating to the Late Ocucaje (phase 10)/Nasca 1 transition (Feature 1004 in Fig. 3), which suggests that the tree was introduced here sometime in the half a millennium or so that elapsed between phase 4 and the end of the Ocucaje period. In any case, the profuse remains of both pacay and guava in these Late Nasca contexts also speak of altered ecological conditions because today neither of these large, simple leafed trees can be grown easily anywhere in the lower Ica Valley because of its wind regime, a point we shall return to shortly.
Chilli peppers (Capsicum spp.), meanwhile, are of course members of the Solanaceae family, which counts among its members many of those New World plant domesticates whose worldwide impact has been most profound, including the potato (Solanum tuberosum), tomatoes (Solanum lycopersicum) and tobacco (Nicotiana spp.). Some of these show vestiges of a general feature of the Solanaceae—toxicity. Moreover, a number of other important New World domesticates, including, for instance, the sweet potato and manioc, also derive from families of plants that include many toxic and psychoactive relatives. As Jones (2009) notes, this suggests a particularly refined ecological intelligence in the long process of human-plant co-evolution that underlies domestication in the Americas. Many wild Solanaceae plants grow as weeds today in the fields of the south coast, including those of the genera Solanum, Nolana, Datura, Nicotiana, Physalis and Lycium. Indeed, Solanaceae is the fourth most common family in our modern plant collections on the south coast (Whaley et al. 2011).
Two of these, very common in the fields of Huerto Huamaní, Samaca, produce huge quantities of tiny delicious tomato fruits (Solanum pimpinellifolium and S. peruvianum—known as tomatillo, Fig. 8d; Sagástegui Alva 1973, pp. 377–382). These prostrate herbs grow as dense mats over extensive open parts of the lower Ica Valley and although their fruits are individually small, in summer (January) they produce great quantities of wild tomatoes. The history of tomato domestication is currently uncertain with Peru and Mexico being variously cited as centres of origin (Paran and van der Knaap 2007). But the quantities of S. pimpinellifolium seeds identified in our Late Nasca and other contexts, many with coprolite encrustations, strongly suggest that wild tomatoes were part of the human diet in the lower Ica Valley in pre-Hispanic times.

Selected examples of agrestal weed macrofossils recovered from Middle Horizon midden contexts in mound 23, S.U. 48, Samaca H-13; a desiccated Amaranthus sp., seeds with coprolite encrustations (S.U. 48), b Sesuvium (Portulaca sp.), c Chenopodium sp., d Solanum sp. (cf. pimpinellifolium or ‘tomatillo’), e Lippia sp. (Syn. Phyla)
Aside from chilli and tomatíllo, the occurrence of other Solanaceae seeds in these ancient middens might hint at other extramundane uses, conferred perhaps by their toxicity. The fruit of S. nigrum for instance contains powerful alkaloids and is poisonous to livestock, although it is used as an anaesthetic (Sagástegui Alva 1973, pp. 377–382). Datura, meanwhile, is powerfully psychoactive (Schultes 1972, pp. 46–51). Unfortunately, however, many seeds of the different genera of Solanaceae can be “confusingly similar” as Martin and Barkley (1961, p. 196) put it, so that we have felt it prudent to lump some of these under Solanum in Group C—weedy plants associated with agriculture.
Also Late Nasca contexts in the lower Ica Valley are part of a seemingly clear trend in Table 2 of increasing intensity of those agrestal weeds (Group C), throughout the various archaeological periods analysed. At the same time, those other indicators of open-area disturbance to riparian woodland, grasses (Group E) and shrubs (Group B), also appear to increase steadily. Together with domesticated plant remains, what we seem to be witnessing in our archaeobotanical record is a gradual increase in the amount of agriculture being carried out over time in these basins. Moreover, along with the aforementioned chenopods and amaranths, Sesuvium portulacastrum (Syn. Portulaca) is another classic halophyte, particularly well adapted to saline conditions (Fig. 8b). So the increasing amounts of Sesuvium in this record may reflect a gradual accumulation of salts, often a consequence of long-term irrigation of arid, low-lying areas (Redman 1999).
The Middle Horizon—collapse
There are no settlement remains on the relict river terraces of Samaca and Ullujaya after the Middle Horizon, which, in itself, suggests that this period from around a.d. 800 to 1000 was one of critical change, (Beresford-Jones et al. 2009b). The archaeobotanical data from mound 23, Samaca H-13 helps elaborate a model for how and why that change occurred.
While its upper contexts were those of a midden dating to Epoch 2 of the Middle Horizon, the contexts beneath were dominated by up to 40% by volume of desiccated poña in situ—the thick layer of tree litter that carpets the desert under huarango tree. These identify mound 23 as a relict nabkha, or phytogenic mound, which accumulate about vegetation in many desert landscapes subject to wind erosion (Cooke et al. 1993, p. 356; Beresford-Jones et al. 2009b, p. 314). This identification tells us, on the one hand, that these relict terraces of the lower Ica Valley were not suddenly denuded of all vegetation following some catastrophic El Niño event, but, on the other, that by Epoch 2 of the Middle Horizon, this was a landscape already partially degraded and being moulded by the region’s extraordinarily strong wind regime. Almost 50 m long, the dimensions of mound 23 speak of a nabkha that built up under the canopy of a very large tree over a considerable period of time. Indeed, the lifespan of a large huarango could easily have exceeded the duration of the entire Middle Horizon (Beresford-Jones in press).
Its upper deposits were full of the rubbish of human settlement: copious marine and terrestrial mollusc shells, lithic flakes, partly spun cotton and even desiccated human coprolites (Fig. 9). They contained abundant fibres of cotton, some of which was partly spun into yarn, a few pumpkin (Cucurbita sp.) seeds, and remains of those same fruit trees which we encountered earlier in Late Nasca deposits: leaf fragments of pacay (Inga feuillei) and guava (Psidium guajava) seeds. They also included moderate numbers of coca (Erythroxylum sp.) seeds (Fig. 10). This latter conjunction of plant remains, noted also in the Late Nasca contexts of midden mound 1, Ullujaya G-8/9 (Table 2), is perhaps rather remarkable.

Selected examples of midden materials recovered from the Middle Horizon midden contexts of mound 23, Samaca H-13; a obsidian flake from heavy fraction, S.U. 42, b land snail shell (Bulimulidae), S.U. 48, c various marine mollusc shells, S.U. 48, d partly spun cotton fibre (Gossypium sp.), S.U. 32, e 0.5 mm flotation sample, S.U. 51, prior to sorting showing dominance of huarango poña, f broken desiccated human coprolite, S.U. 41, containing masticated huarango pod fragments and seeds (marked ‘i.’)

Coca cultivation? A plant assemblage from a Middle Horizon midden context (S.U. 32) of mound 23, Samaca H-13; a selected Erythroxylum sp. (coca) seeds, b selected Psidium guajava (guava) seeds, c detail of Inga feuillei (pacay) leaf fragment
Coca leaf, used as a mild stimulant, is found in south coast archaeological contexts but only becomes relatively common in the Middle Horizon. Prior to this, its remains are very few and restricted Late Nasca contexts (Piacenza 2005; Table 2). The plant therefore seems to have been first used on the south coast rather late, around about the time that the Wari Empire was beginning its expansion into the region. Indeed, Schreiber (1999, 2001) has argued that the expansion of the Wari Empire out of the Ayacucho highlands was driven, at least in part, by seeking new locations for coca cultivation.
Today coca—predominately Erythroxylum coca var. coca, or ‘Huanaco’ coca—is grown at high altitudes on the eastern slopes of the Andes above 1,000 m. But at the time of the Spanish conquest coca consumption was much more widespread; including all over Caribbean South America and into the Isthmus of Panama (Warwick Bray, personal communication). In fact, another species of coca, Erythroxylum novogranatense, is far more flexible in its adaptation and can be grown as low as sea level. In 1875 it was propagated successfully in the Far East and for this reason is sometimes known as ‘Java coca’. Since the work of Plowman (1984), a variety of this species (var. truxillense, or ‘Trujillo’ coca) has been recognised growing on the Peruvian coast. The range of this drought-resistant coca is put historically as ‘chaupi yunga’, an ecozone which Rostworowski de Diez Canesco (1989), pp. 245, 246) defines between 200 and 1,200 m in areas free of cold sea mists. Coca, like coffee, is cultivated in the shade of larger trees (Plowman 1984). Rostworowski de Diez Canesco (1989) cites various 16th century sources which describe Pacific coastal plantations of E. novogranatense growing under the protective shade of two species of fruit tree in particular: pacay and guava. A 1559 document, for instance, describes Quivi in the Chillón Valley: “they harvest nothing from these lands save coca but here they have trees of guabas and guayabas and they don’t sow or harvest maize because this land is only for coca” (Rostworowski de Diez Canesco 1989, p. 252—our translation).
Clearly, it would be precipitous to draw definitive conclusions based upon these data from the anthropogenic assemblages of the Late Nasca and Middle Horizon midden contexts in the lower Ica Valley. Coca may well have arrived here through trade. Nonetheless, the conjunction of coca seeds (rather than leaves, which would more likely be traded) with guava and pacay remains in the same stratigraphic unit of this Middle Horizon midden, is striking at least in light of Rostworowski’s historical documentation. The lower Ica Valley lies at the lower end of the altitude range specified by Rostworowski and is considerably lower than Pataraya, the site which Schreiber speculates may represent a Wari coca plantation in Nazca. Yet, its climate of year-round sunshine is rather peculiar for the Peruvian coast and is, indeed, more typical of the chaupi yunga. Conceivably then, this assemblage might be the remains of a relatively local source of coca cultivation. Moreover, several of the relaciónes cited by Rostworowski de Diez Canesco (1989) describe coca being produced together with cotton, another plant for which we have abundant evidence of production in the lower Ica Valley. This Middle Horizon midden was also full of cotton fibre, seeds, capsule fragments and leaves, and looted Middle Horizon tombs in the lower Ica Valley contain vast quantities of raw cotton fibre.
Be that as it may, these archaeobotanical remains certainly point to changed ecological conditions. For neither coca, nor indeed pacay and guava, whose remains are, as we have already seen, so profuse in both Late Nasca and Middle Horizon middens, can be grown easily today anywhere in the lower Ica Valley, because of its wind regime. If the occurrence of any one of this intriguing conjunction of plants represents local cultivation, then local ecological conditions, specifically the wind protection and sub-canopy microclimatic alteration effected by a much more extensive woodland canopy, must have changed dramatically in the Samaca and Ullujaya basins.
Yet what is otherwise most noticeable about these Middle Horizon middens is their relative dearth of cultivated foods. While they contained large quantities of marine and terrestrial molluscs and indeed, a significantly wider range of gathered marine foods than preceding time periods (Table 3), these contexts contained no obvious food cultivars, aside from pacay, guava and a few zapallo (Cucurbita sp.) seeds (a plant which grows today uncultivated as a ‘volunteer’ plant in the Samaca Basin). Archaeobotanical remains from the Middle Horizon site of Casa Vieja upstream in Callango offer an interesting contrast, for its contexts included the full range of typical south coast crops including maize, beans, potato, sweet potato and manioc (Roque et al. 2003; Cook and Parrish 2005).
Desiccated human coprolite fragments were also extracted by flotation from the deposits of midden mound 23 offering direct evidence of what people were eating, including masticated huarango pods. Many of the other plants identified in these contexts, including Solanum, Sesuvium, Amaranthus, Chenopodium, Crotalaria and Lippia, also showed evidence of coprolite encrustation.
This evokes an analogy with another arid American environment, the Sonoran Desert of the southwestern United States, in which the importance of all these genera as gathered wild plant foods is well documented in a rich ethnobotany, and where such agrestal weeds were encouraged within agricultural fields, precisely because they were an all-important, additional food source (Nabhan 1979, 1985). Indeed, the most abundant plant genera in these Middle Horizon contexts virtually mirrors the list of those wild plant food sources most valued by the peoples of the Sonoran, omitting only certain important cactus species (Castetter and Bell 1942; Nabhan 1979; Hodgson 2001). And by far the most important of all the gathered wild foods that Sonoran peoples depended upon was the pods of Prosopis. Even for maize agriculturalists such as the Akimel O’odham, Prosopis pods remained the single most important foodstuff (Castetter and Bell 1942, p. 56). The pre-eminence of Prosopis in human subsistence evoked by this ethnobotanical analogy and which we note in the archaeobotanical data of the lower Ica Valley, is to be found reflected in almost all the other available archaeobotanical data from the Peruvian south coast (Donaire Vizarreta 1998 [1959], p. 199; Roque et al. 2003, p. 36; Silverman 1993, p. 293; Piacenza 2005).
There is of course wide evidence that the Wari Middle Horizon was, in large part, based upon the intensification of food production in the intermontane valleys of the highlands: the ‘quichua ecozone’. The evidence includes large-scale shifts in settlement and in subsistence regimes from tubers to maize cereals, construction of terracing on a huge scale (Schreiber 1992, 2001) and the introduction of new maize varieties (Grobman et al. 1961; Bird et al. 1984). All appear to have been instigated by some form of ‘state’ in order to supply distant urban populations. Isbell (1988, p. 182) credits the Wari Middle Horizon with the development of a uniquely Andean form of ‘state finance’—what Godelier (1977, p. 188) had first called the “Inca mode of production”—whereby long-standing local traditions of reciprocal exchanges of labour were elaborated into a system of labour-taxation. Indeed, some see the innovations wrought by Wari in these respects as so fundamental and enduring that they defined the course of subsequent Andean civilization (Isbell 1988, p. 182), and settlement patterns even to this day (Schreiber 1992, p. 260; Williams and Nash 2002, p. 255).
Elsewhere on the south coast, interpretations have been advanced that Wari imperialism was engaged in precisely this sort of tribute extraction in the form of coca (Schreiber 1999) or cotton (Kellner and Schoeninger 2008) production. As we have seen there are hints of both in the archaeobotanical record of the Middle Horizon in the lower Ica Valley. Yet that record also suggests that, at least by Epoch 2 of the Middle Horizon, human subsistence seems to have turned full circle, back to the diet of the Early Ocucaje period some two millennia earlier, to consist of gathered, wild marine and plant foods sources. Here, in an ecology very different to that of its highland heartland, the imposition of the Wari ‘mode of production’ seems to have floundered, with consequences which have indeed endured until today.
Coherence with a pollen record
Pollen assemblages from a sequence of contexts in Samaca H-13 (Fig. 12 in Beresford-Jones et al. 2009b, p. 320) seem to mirror the trends we have noted here in the plant macro fossil data, especially once their respective taphonomic differences are taken into account. Those contexts lie under a climbing dune anchored on the western flanks of the Samaca Basin by the unidirectional wind regime. Grain size and other geomorphological analyses reveal these to be a series of aggrading aeolian deposits; surfaces deposited over time by a wind of progressively greater energy, which, in time were buried and sealed under the climbing dune itself (Beresford-Jones et al. 2009a).
The pollen assemblage recorded for the bottom of these sequences is dominated by heavy, entomophilous pollen of woodland species such as Prosopis (70%). Indeed, it is very similar to a modern analogue pollen assemblage recorded from the floor of Usaca, one of the last remaining fragments of ‘old-growth’ woodland left on the south coast of Peru. Woodland pollen declines steadily up the sequence, as pollen from cultivated plants appear and increase. The earliest of these are mostly broken pollen grains from the Malvaceae family, which, by size and morphology, are almost certainly cotton, the only cultivar identified in our earliest midden contexts dating to the Early Horizon. Further up the sequence, however, maize (Zea mays) pollen appears along with that pollen of the Papilionaceae Family, which may well be that of Phaseolus sp. since the two are traditionally inter-cropped. Self-pollinating maize produces a very low pollen rain, which is never dispersed far from the plant (Weir and Eling 1986), such that its occurrence in these contexts is evidence of maize growing in significant quantities in the immediate vicinity. Maize pollen reaches a maximum in contexts also containing Early Nasca pottery sherds—the same period during which a canal system was in operation across the relict terraces of Samaca and Ullujaya (Beresford-Jones et al. 2009b).
As Prosopis pollen declines steadily up the sequence, pollen of chenopods/amaranths, those pioneers both of open ground and classic markers of anthropogenic disturbance, steadily increase. In Middle Horizon contexts, they have come to dominate the pollen assemblage entirely, while Prosopis, other tree species, and maize and cotton have all but disappeared. In sum, just like the plant macrofossil record, the pollen record also reflects a gradual replacement of huarango woodland with cultivation and anthropogenic disturbance, culminating in a landscape entirely dominated by halophytic weeds by the Middle Horizon.
The only real exception to this excellent correlation between plant macro- and microfossil data is from those of trees, predominantly huarango. For the remains of huarango are fairly constant across time in the macrofossil remains of Table 2 (Group A); while in the pollen record they fall from levels almost the same as those of a modern analogue, the forest floor of the Usaca old-growth woodland, to almost nothing. Yet this serves only to emphasise the importance of huarango resources, and shade, to human ecology, for the plant remains in Table 2 are, of course, from anthropogenic contexts. And the huge amounts of huarango poña in the Middle Horizon midden of mound 23, Samaca H-13, are, as we have seen, part of its identification as a relict nabkha—a place where people gathered under the canopy of a single, isolated tree amid a landscape already undergoing gradual change under the hand of the wind (Beresford-Jones et al. 2009a).
Conclusions
The archaeobotanical and other data presented here are consistent with a model of gradual anthropogenic landscape change in the lower Ica Valley which we have presented in detail elsewhere (Beresford-Jones et al. 2009a, b). The model presents a story of a thousand-year transition from relatively undisturbed, old-growth huarango woodland to a controlled, irrigated agricultural landscape during the Early Nasca Period.
Remains of Ocucaje period settlement in the Ullujaya and Samaca basins are few. The earliest occupations we have yet recorded date back to the Chavín Early Horizon (Ocucaje phase 3) at around 750 b.c. Cotton is the only domesticated plant evident in these ancient rubbish remains which otherwise contain no domesticated food plants, but only sea urchin and mussel debris with other marine resources gathered from the Pacific Ocean some 25 km distant and land snails from the lomas, 10 km away. They do however contain some wild plant remains, including gathered wild foods such as huarango beans, and other plants typical of riparian woodland. This evidence suggests that, at this early date, the occupants of the Ullujaya basin were not subsisting on any form of crop agriculture. They carried out some floodplain agriculture growing cotton for textiles and fishing materials but seem otherwise to have made little impact upon the natural dry forest ecology.
By the end of the Ocucaje Period (phase 10/Nasca 1 transition) there is rather more evidence for settlement in Samaca and Ullujaya, including the large site of 1004 (Fig. 3), which we have not excavated or systematically floated, but whose looted mortuary contexts also contain abundant evidence of habitation, as if bodies were interred within a settlement site. Such evidence includes heavy grinding stones, large quantities of camelid bones, fragments of cotton and camelid fibre textiles, cotton fishing nets, marine and terrestrial molluscs, desiccated pacay leaves and pods, peanut (Arachis hypogaea) shells, pumpkin (Cucurbita sp.) seeds, tubers of yuca (Manihot esculenta) and small popcorn maize cobs. This suggests an intensification of both agriculture and interactions between the basins of the lower Ica Valley and the highlands at this time (Fig. 11). Moreover, this site lies high up the edge of the Samaca Basin well over a hundred metres above the current vegetated river floodplain and the traces of later settlement. This, in itself, suggests some radical change in the geomorphology of the basin and diminution of the extent of the vegetation within it since that ancient time.

Selected midden materials recovered from Late Ocucaje (phase 10)/Nasca 1 transition site 1004, Samaca Basin; a peanut (Arachis hypogaea) shell, b pumpkin (Cucurbita sp.), c marine and terrestrial molluscs, d maize (Zea mays) cobs, e pacay (Inga feuillei) pod exocarp and seed, f yuca (Manihot esculenta) tubers
By the subsequent Early Intermediate Periods, midden and pollen data tell a story of a changed, farmed landscape. Rubbish middens of the Late Nasca Period contained a complete suite of south coast domesticates including maize, beans, pumpkin, peanuts, achira, pacay, chilli peppers surrounded by groves of native huarango and the cultivar fruit trees, guava and pacay. The occurrence of plants such as pacay, guava, achira, Indigofera and yuca (manioc) in this archaeobotanical record implies a once much more extensive and mature woodland ecology in these basins, for today none of these can persist here in the region’s extraordinary wind regime. Meanwhile, just like the pollen sequence, the plant macrofossil record showed a clear trend of increasing intensity of agrestal weeds such as chenopods/amaranths, up through the various archaeological periods analysed. At the same time those other indicators of open-area disturbance to riparian woodland, grasses and shrubs, also increased steadily. Sesuvium, another classic halophyte, and particularly well-adapted to saline conditions, increased markedly in the plant macrofossil record, which may well reflect a gradual increase in salinity, often a consequence both of long-term irrigation and of deforestation.
While our archaeobotanical record bears witness to the gradual intensification of agriculture being carried out over time in these basins, nonetheless, our model still envisages a landscape largely dominated by trees during the Nasca Period. For it is quite simply impossible to carry out successful agriculture in the environment of the lower Ica Valley without it being underpinned by huarango. We have surveyed almost 400 relict huarango stumps across wind-deflated surfaces of the H-13 and G-8/9 river terraces (Fig. 4). On various lines of evidence these relict trunks can be interpreted as the remains of trees that met their demise at some time prior to the Late Intermediate Period (Beresford-Jones et al. 2009b). This leguminous tree was an important source of food, forage, timber and fuel for people. Furthermore, as we have discussed elsewhere, it was the ecological ‘keystone’ species in this desert region, fixing nitrogen and enhancing soil fertility and moisture, ameliorating desert extremes in the microclimate beneath its canopy and underpinning the floodplain with one of the deepest root systems of any tree known (Beresford-Jones in press; Beresford-Jones et al. 2009b; Whaley et al. 2010, 2011).
However by the Middle Horizon human subsistence in the lower Ica Valley would seem to have turned a full circle, back to the subsistence regime of the Early Horizon almost two millennia earlier. Rubbish middens from this time contained few obvious food plant domesticates. Rather, they were again full of marine and land snail molluscs from distant locations, in conjunction with a plant assemblage dominated by those wild plant foods whose importance to human diet are recorded in other, analogous arid American environments. The consumption of the most important of these, the fruit of the huarango, is evident in human coprolites from those Middle Horizon middens.
Thus, the model presents a story, commencing in the Late Nasca Period and culminating in the Middle Horizon, of a gradual removal of huarango dominated riparian woodland for the purpose of increasing agricultural production. In time that process undermined the multiple ecological influences of the genus Prosopis in this environment, and in particular the protection it affords in an environment of high-energy, episodic flood events and of one of the world’s strongest and most persistent wind regimes. The loss of riparian vegetation from the floodplain permitted high-energy, lateral instability in river channel form and down-cutting. So, while chaotic climatic fluctuations like El Niño and La Niña were likely tipping points, causing geomorphological and ecological thresholds to be breached, the seeds of that sudden instability had been sown long before through this process of gradual, human-induced change. Once ecological thresholds were crossed, however, change became rapid. Irrigation systems were both damaged in the short-term by the resultant flood events, and rendered inoperative over the long-term by the river’s entrenchment. Hence, agriculture was largely abandoned across the relict terraces of the Samaca and Ullujaya basins.
Indeed, by the end of the Middle Horizon when human occupants of the valley used mound 23 as a sheltered spot to consume a diet dominated by gathered, wild foodstuffs, the surrounding area had already undergone and was still undergoing gradual change from the stable, productive agricultural landscape that had prevailed during the preceding Early Intermediate Period. Even the identification of mound 23 as a relict nabkha is itself indicative of gradual environmental change, for only land surfaces that are already subject to considerable wind erosion give rise to nabkha formation. For by now the strong unidirectional wind regime of the region was entraining particles from a dry surface, some of which were being deposited in a few isolated nabkha formations like that of mound 23. The aeolian deflation of the H-13 and G-8/9 terrace surfaces had begun. Perhaps the clearest evidence of the irreversible ecological change wrought in the lower Ica Valley is that, by the Middle Horizon, these now relict river terraces had become the location of cemeteries. In all time periods these are inevitably sited outside agricultural lands and beyond the corruption of moisture and fertility. By the Middle Horizon, a landscape of the dead was being set amidst the traces of a lost Nasca landscape of the living (Beresford-Jones in press).
At some point prior to the Late Intermediate even the large individual huarangos, about which nabkha mound 23 had formed, had died, or been dug out for firewood or construction materials. By then gradual ecological change on the relict terraces of the lower Ica Valley had ceased, while geomorphological change was accelerating. With no vegetation to impede the wind, the process became irreversible. Wind deflation of land surfaces increased. No significant cultural materials from this, or any later cultural phases, are noted across the great expanses of desolate terraces in the lower Ica Valley. Today, some 1,000 years after the Middle Horizon, the surfaces of these relict terraces with their canal systems thrown into inverted relief, their mixed scatters of sherds from various time epochs and their relict tree trunks and other fossil evidence sealed beneath climbing dunes, stand in testimony to this story of what Doolittle (2000) p. 6) calls “anthropogenic geomorphology”.
References
Balée W (1998) Historical ecology: premises and postulates. In: Balée W (ed) Advances in historical ecology. Columbia University Press, New York, pp 13–29
Beresford-Jones DG (2005) Pre-hispanic prosopis-human relationships on the south coast of Peru: Riparian Forests in the Context of Environmental and Cultural Trajectories of the Lower Ica Valley. PhD thesis, University of Cambridge, http://www.arch.cam.ac.uk/dgb27/
Beresford-Jones DG (in press) The Lost Woodlands of Ancient Nasca. A Case-study in Ecological and Cultural Collapse. British Academy Postdoctoral Monograph Series, Oxford University Press
Beresford-Jones DG, Arce S (2010) Informe de los Trabajos Realizados Durante la Temporada 2007. Presentada ante la Comisión Técnica de Arqueología. Instituto Nacional de Cultura del Perú. Lima, Peru, http://www.arch.cam.ac.uk/dgb27/
Beresford-Jones DG, Arce S, Grimaldo C (2004) Informe de los Trabajos Realizados Durante la Temporada 2002. Presentada ante la Comisión Técnica de Arqueología. Instituto Nacional de Cultura del Perú. Lima, Peru, http://www.arch.cam.ac.uk/dgb27/
Beresford-Jones DG, Arce Torres S, Whaley OQ, Chepstow-Lusty AJ (2009a) The Role of Prosopis in ecological and landscape change in the Samaca Basin, lower Ica Valley, south coast Peru from the early horizon to the late intermediate period. Latin Am Antiq 20:303–332
Beresford-Jones DG, Lewis HA, Boreham S (2009b) Linking cultural and environmental change in Peruvian prehistory: geomorphological survey of the Samaca Basin, south coast Peru. Catena 78:234–249
Beresford-Jones DG, Alarcón C, Arce S, Chepstow-Lusty A, Whaley O, Sturt F, Gorriti M, Portocarrero O, Cadwallader L (2011) Ocupación y subsistencia durante el horizonte temprano en el contexto de cambios ecológicos a largo plazo en las cuencas de Samaca y Ullujaya, valle baja de Ica. In: Kaulicke P, Onuki Y (eds) El Periodo Formativo en el Perú. Boletín de Arqueología pucp 13, Fondo Editorial de la Pontificia Universidad Católica del Perú
Berglund BE, Ralska-Jasiewiczowa M (1986) Pollen analysis and pollen diagrams. In: Berglund B (ed) Handbook of holocene palaeoecology and palaeohydrology. Wiley, Chichester, pp 455–484
Bird RM, Browman L, Durbin ME (1984) Quechua and maize: mirrors of Central Andean culture history. Journal of the Steward Anthropological Society 15:187–240
Brack AE (1999) Diccionario Enciclopedico de Plantas Utiles del Perú. Cusco: Centro Regionales Andinos ‘Bartolomé de las Casas’
Brako L, Zarucchi J (1993) Catalogo de las Angiospermas y Gimnospermas del Perú. Missouri Botanical Garden, St. Louis, Missouri
Burger RL (1992) Chavin and the origins of Andean civilisation. Thames and Hudson, London
Burghardt AD, Brizuela MM, Mom MP, Albán L, Palacios RA (2010) Análisis numérico de las especies de Prosopis L. (Fabaceae) de las costas de Perú y Ecuador. Revista Peruana de Biología 17:317–323
Butzer KW (1982) Archaeology as human ecology. Cambridge University Press, Cambridge
Castetter EF, Bell WH (1942) Pima and Papago Indian Agriculture. University of New Mexico Press, Albuquerque, New Mexico
Conklin WJ (2008) The culture of Chavín textiles. In: Conklin WJ, Quilter J (eds) Chavín: art, architecture and culture. Cotsen Institute of Archaeology Monograph 61. Cotsen Institute of Archaeology, Los Angeles, pp 261–278
Cook AG (1994) Invesitigaciones de Reconocimiento Arqueologico en la Parte Baja del Valle de Ica. Informe Final (1988–1990) al Instituto Nacional de Cultura
Cook AG (1999) Asentamientos Paracas en el Valle Bajo de Ica, Perú. Gaceta Arqueologica Andina 25. INDEA, Lima, Peru
Cook AG, Parrish N (2005) Gardens in the desert: archaeobotanical analysis from the lower Ica Valley, Peru. Andean Past 7:135–156
Cooke RU, Warren A, Goudie AS (1993) Desert geomorphology. University College London Press, London
DeLeonardis L (1997) Paracas settlement in Callango, lower Ica Valley, 1st Millennium BC, Perú. Unpubl. PhD thesis, Catholic University of America, Washington, DC. University Microfilms Inc., Ann Arbor, Michigan
DeLeonardis L (2005) Early Paracas cultural contexts: new evidence from Callango. Andean Past 7:27–55
Dillehay TD, Rossen J, Andres T, Williams DE (2007) Preceramic adoption of peanut, squash, and cotton in Northern Peru. Science 316:1,890-1,893
Donaire Vizarreta J (1998 [1959]) Campiña Iqueña. Aspectos Folklóricos (1941), 4th edn. Ital Perú, Lima
Doolittle WE (2000) Cultivated landscapes of Native North America. Oxford University Press, Oxford
Gade DW (1966) Achira, the Edible Canna, its cultivation and use in the Peruvian Andes. Econ Bot 20:407–415
Galan de Mera A (1999) La Clases Fitosociologicas De la Vegetación del Perú. Boletín de Lima 117:84–94
Godelier M (1977) The concept of ‘social and economic formation’: the Inca example. In: Perspectives in Marxist Anthropology. Cambridge University Press, Cambridge
Grobman A, Salhuana W, Sevilla R, Mangelsdorf P (1961) Races of Maize in Peru. National Research Council Publication 915. National Academy of Sciences, Washington
Harris PJC, Pasiecznik NM, Smith SJ, Billington JM, Ramírez L (2003) Differentiation of Prosopis juliflora (Sw.) DC and P. pallida (H. & B. ex. Willd.) H.B.K. using foliar characters and ploidy. For Ecol Manag 180:153–164
Hather JG (1993) An archaeobotanical guide to root and tuber identification. Volume 1 Europe and South West Asia. Oxbow Monograph 28, Oxbow Books, Oxford
Hather JG (2000) Archaeological parenchyma. Archetype Publications, London
Hecht N (2009) Of layers and shreds: a context-based relative chronology of the Nasca Style Pottery from Palpa. In: Reindel M, Wagner GA (eds) New technologies for archaeology: multidisciplinary investigations in Palpa and Nasca, Peru. Springer, Berlin, pp 207–230
Hodgson WC (2001) Food plants of the Sonoran Desert. University of Arizona Press, Tucson
Holden TG (1991) Evidence of prehistoric diet from northern Chile: coprolites, gut contents and flotation samples from the Tulán Quebrada. World Archaeol 22:320–331
Hubbard RNLB, Clapham A (1992) Quantifying macroscopic plant remains. Rev Palaeobot Palynol 73:117–132
Isbell WH (1988) City and state in Middle Horizon Huari. In: Keatinge RW (ed) Peruvian prehistory. Cambridge University Press, Cambridge, pp 164–189
Jones MK (2009) Moving North: archaeobotanical evidence for plant diet in middle and upper Palaeolithic Europe. In: Richards MP, Hublin J-J (eds) The evolution of hominid diets: integrating approaches to the study of palaeolithic subsistence. Springer, London, pp 171–180
Kellner CM, Schoeninger MJ (2008) Wari’s imperial influence on local Nasca diet: the stable isotope evidence. J Anthropol Archaeol 27:226–243
Kroeber AL (1944) Peruvian Archaeology in 1942. Viking fund publications in anthropology 4, New York
Mächtle B, Unkel I, Eitel B, Kromer B, Schiegl S (2010) Molluscs as evidence for a late Pleistocene and early Holocene humid period in the southern coastal desert of Peru (14.5 S). Quat Res 73:39–47
Martin AC, Barkley WD (1961) Seed identification manual. University of California Press, Berkeley
Massey SA (1991) Social and political leadership in the lower Ica Valley: Ocucaje Phases 8 and 9. In: Paul A (ed) Paracas art and architecture. University of Iowa Press, Iowa City, pp 349–416
McCauley JF, Breed CS, Grolier MJ (1977) Yardangs. In: Doehring DO (ed) Geomorphology in arid regions. Allen and Unwin, Boston, pp 233–272
Molas (1994) Archaeological site manual, III edn. Museum of London Archaeological Service, Museum of London
Mom MP, Burghardt AD, Palacios RM, Albán L (2002) Los algarrobos Peruanos: Prosopis pallida y su delimitación. Arnaldoa 9:39–48
Moseley ME (1975) The maritime foundations of Andean Civilization. Cummings Archaeological Series, Merlo Perk, Cummings
Nabhan GP (1979) The ecology of floodwater farming in arid south-western North America. Agro-Ecosystems 5:245–255
Nabhan GP (1985) Gathering the desert. University of Arizona Press, Tucson
Paran I, van der Knaap E (2007) Genetic and molecular regulation of fruit and pepper domestication traits in tomato and pepper. J Exp Bot 59:3,841–3,852
Parker Gay S Jr (1999) Observations regarding the movement of Barchan sand dunes in the Nazca to Tacna Area of southern Peru. Geomorphology 27:279–293
Pennington TD (1997) The genus Inga: botany. Royal Botanic Gardens, Kew
Pennington TD, Fernandes ECM (eds) (1998) The genus Inga: utilization. Royal Botanic Gardens, Kew
Piacenza L (2005) Evidencias botánicas en asentamientos Nasca. Boletín del Museo de Arqueología y Antropolgía (UNMSM) 5:3–13
Plowman T (1984) The origin, evolution and diffusion of coca, Erythroxylum spp., in South and Central America. In: Stone D (ed) Pre-columbian plant migration. Papers of the peabody museum of archaeology and ethnology 76:125–163
Ramírez R, Córdova S, Caro K, Duárez J (2003) Response of a land snail species (Bostryx conspersus) in the Peruvian central coast lomas ecosystem to 1982–1983 and 1997–1998 El Niño Events. In: Haas J, Dillon MO (eds) El Niño in Peru: biology and culture over 10,000 years. Fieldiana Botany 43. Field Museum of Natural History, Chicago, pp 10–23
Rea AM (1997) At the desert’s edge: an ethnobotany of the Gila River Pima. University of Arizona Press, Tucson
Redman CL (1999) Human impact on ancient environments. University of Arizona Press, Tuscon
Roe PG (2008) How to build a raptor: why the Dumbarton Oaks “Scaled Cayman” Callango textile is really a Chavín jaguaroid Harpy Eagle. In: Conklin WJ, Quilter J (eds) Chavín: art, architecture and culture. Cotsen Institute of Archaeology Monograph 61. Cotsen Institute of Archaeology, Los Angeles, pp 181–216
Roque J, Cano A, Cook A (2003) Restos vegetales del sitio arqueológico Casa Vieja, Callango (Ica). Revista Peruano de Biología 10:33–43
Rossel Castro A (1977) Arqueología Sur del Perú. Editorial Universo S.A, Lima
Rostworowski de Diez Canesco M (1989) Costa Peruana Prehispanica. Instituto de Estudios Peruanos, Lima
Sagástegui Alva A (1973) Manual de las Malezas de la Costa Norperuana. Universidad Nacional de Trujillo, Trujillo
Schreiber KJ (1992) Wari Imperialism in Middle Horizon Peru. Anthropological Papers 87. University of Michigan, Museum of Anthropology, Ann Arbor
Schreiber KJ (1999) Regional approaches to the study of prehistoric empires: examples from Ayacucho and Nasca, Peru. In: Billman BR, Feinman GM (eds) Settlement pattern studies in the Americas: fifty years since Virú. Smithsonian Institution Press, Washington, DC, pp 160–171
Schreiber KJ (2001) Los wari en su contexto local: Nasca y Sondondo. In: Kaulicke P, Isbell WH (eds) Huari y Tiwanaku: Modelos vs. Evidencias. Boletín de Arqueología PUCP 4, Lima, pp 425–448
Schultes RE (1972) Hallucinogens in the Western Hemisphere. In: Furst PT (ed) Flesh of the Gods: the ritual use of Hallucinogens. Waveland Press, Prospect Heights, IL, pp 3–54
Silverman H (1993) Cahuachi in the ancient Nasca World. University of Iowa Press, Iowa City
Stephens SG (1970) The botanical identification of archaeological cotton. Am Antiq 35:367–373
Stone-Miller R (1992) To weave for the sun: ancient andean textiles in the museum of fine arts. Boston, Thames and Hudson, London
Strong WD, Willey GR, Corbett JM (1943) Archaeological studies in Peru. Columbia University Press, New York
Ugent D, Pozorski S, Pozorski T (1984) New evidence for ancient cultivation of Canna edulis in Peru. Econ Bot 38:417–432
Unkel I, Kromer B (2009) The clock in the corn cob. In: Reindel M, Wagner GA (eds) New technologies for archaeology: multidisciplinary investigations in Palpa and Nasca, Peru. Springer, Berlin, pp 231–244
Weir GH, Eling HH Jr (1986) Pollen evidence for economic plant utilization in pre-hispanic agricultural fields of Jequetepegue Valley, Northern Peru. In: Matos R, Turpin SA, Eling HH Jr (eds) Andean archaeology, Papers in memory of Clifford Evans. Monograph XXVII, Institute of Archaeology, University of California, Los Angeles
Westengen OT, Huamán Z, Heun M (2005) Genetic diversity and geographic pattern in early South American cotton domestication. Theor Appl Gen 110:392–402
Whaley OQ, Orellana A, Pérez E, Tenorio M, Quinteros F, Mendoza M, Pecho O (2010) Plantas y Vegetación de Ica, Perú–Un recurso para su restauración y conservación. Royal Botanic Gardens, Kew
Whaley OQ, Beresford-Jones DG, Milliken W, Orellana A, Smyk A, Leguia J (2011) An ecosystem approach to restoration and sustainable management of dry forest in Southern Peru. Kew Bull 65:1–29
Williams PR, Nash DJ (2002) Imperial interaction in the Andes: Huari and Tiwanaku at Cerro Baúl. In: Isbell WH, Silverman H (eds) Andean archaeology I: variations in sociopolitical organization. Plenum, New York, pp 243–266
Acknowledgments
We thank all the members of the Proyecto de Investigación Arqueológica Samaca for their fieldwork and many other invaluable contributions, especially Mario Advíncula, Claudia Grimaldo, Kevin Lane, Sandy Pullen, Fraser Sturt and, in particular, its co-director Susana Arce Torres of the Museo Regional de Ica. For the wider interpretations drawn on in this paper we acknowledge with thanks the contributions of Alex Chepstow-Lusty (pollen identifications), Manuel Gorriti (mollusc identifications), Helen Lewis (micromorphological interpretations) and Oscar Portocarrero (plant macrofossil identification). We also thank two anonymous reviewers for their useful suggestion and comments on this paper and Steve Boreham, Warwick Bray, Elizabeth DeMarrais, Charly French, Martin Jones and all members, past and present, of the George Pitt-Rivers Bioarchaeology Laboratory, University of Cambridge, for their help and advice. We thank the Instituto Nacional de Cultura (INC), Perú for granting us research permits. And, above all, we thank the people of the Escuela Libre de Puerto Huamaní and Don Alberto Benavides G. This research was funded by the Natural Environmental Research Council (NERC), the British Academy and the McDonald Institute for Archaeological Research, Cambridge.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by L.A. Newsom.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Beresford-Jones, D.G., Whaley, O., Ledesma, C.A. et al. Two millennia of changes in human ecology: archaeobotanical and invertebrate records from the lower Ica valley, south coast Peru. Veget Hist Archaeobot 20, 273–292 (2011). https://doi.org/10.1007/s00334-011-0292-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-011-0292-4