
Abstract
Biodiesel from microalgae seems to be the only renewable biofuel that has the potential to completely replace the petroleum-derived transport fuels. Therefore, improving lipid content of microalgal strains could be a cost-effective second generation feedstock for biodiesel production. Lipid accumulation in Scenedesmus obliquus was studied under various culture conditions. The most significant increase in lipid reached 43% of dry cell weight (dcw), which was recorded under N-deficiency (against 12.7% under control condition). Under P-deficiency and thiosulphate supplementation the lipid content also increased up to 30% (dcw). Application of response surface methodology in combination with central composite rotary design (CCRD) resulted in a lipid yield of 61.3% (against 58.3% obtained experimentally) at 0.04, 0.03, and 1.0 g l−1 of nitrate, phosphate, and sodium thiosulphate, respectively for time culture of 8 days. Scenedesmus cells pre-grown in glucose (1.5%)-supplemented N 11 medium when subjected to the above optimized condition, the lipid accumulation was boosted up to 2.16 g l−1, the value ~40-fold higher with respect to the control condition. The presence of palmitate and oleate as the major constituents makes S. obliquus biomass a suitable feedstock for biodiesel production.
Similar content being viewed by others

Introduction
In recent years, biodiesel is receiving widespread attention owing to non-toxic, biodegradable and renewable fuel. It also contributes no net carbon dioxide or sulfur to the atmosphere and emits less gaseous pollutants than the conventional diesel fuel (Vicente et al. 2004). Extensive studies have been conducted on using vegetable oils as diesel fuel. The focus has mainly been on oils like soybean, rapeseed, sunflower, and safflower (Lang et al. 2001), which are essentially edible in nature. Most recently, research effort has been aimed at identifying suitable biomass species, which can provide high energy outputs to replace the conventional fossil fuels (Miao and Wu 2004). Few attempts have however been made for producing biodiesel from non-edible sources like used frying oil, greases, tallow, lard, jatropha, and mahua oils (Mittelbach et al. 1992; Alcantara et al. 2000; Canakci and Gerpen 2001; Becker and Francis 2002; Dorado et al. 2002; Ghadge and Raheman 2005). Nevertheless, the cost of biodiesel production is still a major obstacle for large scale commercial exploitation; mainly due to the high feed cost of vegetable oils (Lang et al. 2001). Microalgae are emerging as one of the most promising sources of biodiesel because of their high photosynthetic efficiency and faster growth as compared to any energy crop (Minowa et al. 1995). They reproduce quickly and can be harvested day after day (Haag 2007). However, the lipid content in the microalga required to be high, otherwise the economic performance would be hard to achieve (Xu et al. 2006).
Each species of microalga produces different ratios of lipids, carbohydrates, and proteins. Nevertheless, these tiny organisms have the ability to manipulate their metabolism through simple manipulations of the chemical composition of the culture medium (Behrens and Kyle 1996), thus high lipid productivity can be achieved. For example, supplementation of glucose to the growth medium under nitrate limitation was found to raise the crude lipid content up to 55% of the dry cell weight (dcw, against 15% under control condition) in Chlorella protothecoides (Miao and Wu 2004). In Chlorella vulgaris lipid contents of 40% and 56.6% (dcw) were recorded, when grown respectively in low nitrogen- and iron-supplemented medium (Illman et al. 2000; Liu et al. 2008). Thus, oil content of 20–30% is easy to induce in several microalgal species (Chisti 2007). Exceptionally, an oil content of 86% (dcw) was reported in the brown resting state colonies of Botryococcus braunii (Brown et al. 1969). However, the major obstacle in focusing B. braunii as the industrial organism for biodiesel production is its poor growth rate (Dayananda et al. 2007).
In our preliminary experiments with Scenedesmus obiliquus, lipid accumulation was studied under varied pH, temperature, and spectral quality, in presence of heavy metals, heat and chilling stresses, N and P deficiencies/limitations, addition of various carbon sources, etc. with an aim to establish the actual potential of the organism for lipid accumulation. Results showed that the major factors affecting the performance of culture in the terms of lipid yield are the concentrations of nitrate, phosphate, and sodium thiosulphate. The duration of culture (time culture) was also found to affect lipid productivity significantly. In the traditional ‘one-factor-at-a-time’ technique, where one independent variable is studied while maintaining other factors at a fixed level is not only time consuming, but also it does not guarantee the determination of the real optimal condition. Moreover, this technique is unable to detect the frequent interactions between two or more factors, which may lead to inaccurate conclusions. In this report, S. obiliquus biomass was examined as a potent source for biodiesel by evaluating the interrelationships between the critical parameters that influence the lipid yield with the help of response surface methodology (RSM). The oil was characterized and fatty acid profile was also analyzed.
Materials and methods
Organism and growth conditions
Axenic cultures of the green alga S. obliquus (Trup.) Kütz. from the culture collection Gottingen (SAG 276-3a) were grown in 150-ml Erlenmeyer flasks containing 50 ml N 11 medium (Soeder and Bolze 1981) at pH 6.8. The cultures were performed in a temperature controlled incubator at 25 ± 2°C under a photoperiod of 14:10 h at light intensity of 75 µmol photon m−2 s−1 PAR without sparging with air or CO2. The cultures were hand shaken two to three times daily to avoid sticking. This was referred as control culture.
Estimation of dry weight
Dry cell weight was determined gravimetrically according to Rai et al. (1991). A known volume of algal culture was centrifuged at 5,000 rpm for 10 min and the harvested biomass was dried under vacuum at 60°C till it reach a constant weight.
Extraction of lipid from algal biomass
Extraction of lipid was done following the protocol of Bligh and Dyer (1959). A 15-ml glass vial containing algal biomass, 2 ml methanol, and 1 ml chloroform were added and kept for 24 h at 25°C. The mixture was agitated in a vortex for 2 min. One milliliter of chloroform was again added and the mixture was shaken vigorously for 1 min. After, 1.8 ml of distilled water was added and the mixture was mixed in a vortex again for 2 min. The layers were separated by centrifugation for 10 min at 2,000 rpm. The lower layer was filtered through Whatman No. 1 filter paper into a previously weighed clean vial (W1). Evaporation was carried on in a water bath and the residue was further dried at 104°C for 30 min. The weight of the vial was again recorded (W2). Lipid content was calculated by subtracting W1 from W2, and was expressed as % dcw.
Effect of N and P limitation/deficiency on lipid yield
To study the effect of nitrate and phosphate limitations on lipid accumulation, S. obliquus cells were grown at different concentrations of nitrate (0.005–0.1 g l−1) and phosphate (0.06–0.1 g l−1). N-deficiency was achieved by substituting KNO3 of the medium with equimolar concentrations of KCl. For P-deficiency, Scenedesmus cells were transferred to the mineral salt medium, where Na2HPO4.H2O and KH2PO4 were replaced by equimolar concentrations of Na2SO4 and KCl, respectively.
Effect of glucose and thiosulphate supplementation on lipid yield
Effect of mixotrophy on growth and lipid accumulation was studied by supplementing the nutrient medium with various concentrations (0.2–2.25%) of glucose. Similarly, lipid accumulation was quantified in cells grown in presence of different concentrations of sodium thiosulphate.
Optimization study for lipid yield
A five-level-four-factor central composite rotary design (CCRD) obtained by using the commercial statistical package, Design Expert-version 7.1.1 (Stat-Ease, Minneapolis, MN, USA), was employed to find out the interactive effects of four variables, viz. concentrations of nitrate (A), phosphate (B), thiosulphate (C), and time culture (D) on lipid production in 30 runs (Montgomery 2001). Stationary phase cultures of S. obliquus were transferred to N 11 medium with varying concentrations of nitrate, phosphate, and thiosulphate as given in Table 1. Duration of culture was employed as per the experimental design. The experimental data obtained from CCRD were analyzed by RSM. To optimize the level of each factor for maximum response, ‘point optimization’ technique was employed.
Analytical procedure
The saponification and acid values of the microalgal oil were determined following Vicente et al. (2004). The average molecular weight of the oil was calculated with the help of saponification and acid values as per Xu et al. (2006).
The acid catalyzed transesterification of algal oil was carried out using 60:1 molar ratio of methanol to oil and the top organic layer was taken for analysis by thin layer chromatography (TLC) and gas chromatography-mass spectrometry (GC-MS) with the help of Gas Chromatograph (Autosystem XL) equipped with Turbomass Gold Mass Spectrometer (Perkin-Elmer, Shelton, CT, USA). PE-5® phenyl, methylpolysiloxane capillary column (30 m × 0.25 mm × 0.25 µm) was used for the analysis.
Results
Accumulation of lipid in relation to growth
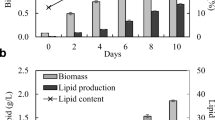
The time-course of growth and accumulation of lipid in S. obliquus under batch mode is presented in Fig. 1. Growth of the test organism increased steadily with a lag of 2 days followed by the logarithmic phase and attained the stationary phase at about 21 days. Accumulation of lipid though started at the early phase of growth, maximum accumulation (12.7% dcw) was observed at the stationary phase.

Lipid content of S. obliquus with reference to growth
Effect of N and P limitations/deficiencies on lipid accumulation
Limitations/deficiencies of nitrate and phosphate appeared to be suitable stimulants for lipid accumulation (Fig. 2a, b) in the tested microalga S. obliquus. The most significant enhancement in lipid production up to 43% (dcw) and 29.5% (dcw) was obtained when the stationary phase cultures were transferred to the media deficient in nitrate for 7 days and phosphate for 3 days, respectively. However, biomass yield under P- and N-limitations/deficiencies depicted a negative trend (Fig. 3a, b), which was more pronounced for nitrate than phosphate.

Effect of various concentrations of a nitrate (open circle) control, (close square) 0.1 g l−1 (N), (close triangle) 0.02 g l−1 (N), (open square) 0.005 g l−1 (N), and (multiplication symbol) N-deficient; b phosphate (open circle) control, (close square) 0.1 g l−1 (P), (close triangle) 0.06 g l−1 (P), (open square) 0.03 g l−1 (P), and (multiplication symbol) P-deficient; and c sodium thiosulphate (open circle) control, (close square) 1.2 g l−1 sodium thiosulphate, (close triangle) 0.9 g l−1 sodium thiosulphate, (open square) 0.6 g l−1 sodium thiosulphate and (multiplication symbol) 0.3 g l−1 sodium thiosulphate on lipid content of S. obliquus

Effect of various concentrations of a nitrate (open circle) control, (close square) 0.1 g l−1 (N), (close triangle) 0.02 g l−1(N), (open square) 0.005 g l−1 (N), and (multiplication symbol) N-deficient; b phosphate (open circle) control, (close square) 0.1 g l−1 (P), (close triangle) 0.06 g l−1 (P), (open square) 0.03 g l−1 (P), and (multiplication symbol) P-deficient; and c sodium thiosulphate (open circle) control, (close square) 1.2 g l−1 sodium thiosulphate, (close triangle) 0.9 g l−1 sodium thiosulphate, (open square) 0.6 g l−1 sodium thiosulphate, and (multiplication symbol) 0.3 g l−1 sodium thiosulphate on biomass yield of S. obliquus
Effect of glucose supplementation
When the cells were cultured in presence of exogenous glucose the biomass production showed increasing trend with increasing glucose concentration (Table 2). Most significant enhancement in biomass yield up to 5.1 g l−1 was obtained when the cells were cultured in 1.5% of glucose-supplemented medium for 10 days. Although supplementation of glucose was not found stimulatory for lipid accumulation on percent dry weight basis, the total lipid yield was increased up to 586 mg l−1 in 1.5% glucose-supplemented culture, which was ~10-fold higher against 55.5 mg l−1 under control condition (Table 2).
Effect of sodium thiosulphate supplementation
Figure 2c demonstrates the effect of sodium thiosulphate supplementation on lipid pool of the tested alga. Under supplementation of 0.6 g l−1 of sodium thiosulphate, the lipid content was raised up to 30.4% (dcw). Further increasing sodium thiosulphate concentration reduced the lipid accumulation. A negative effect of thiosulphate supplementation on biomass yield was also evident (Fig. 3c).
Optimization of lipid yield
The results of CCRD for studying the interactive effects of the four independent variables (concentrations of nitrate, phosphate, and thiosulphate, and time culture) on accumulation of lipid are presented in Table 3. The yield varied between 13.02% and 57.47% (dcw) at different combinations of the variables. The predicted values, calculated by using the model were in the range of 11.16–56.62% (dcw). Regression analysis of the experimental design demonstrated that the linear model terms (A, B, C, and D), quadratic model terms (A2, B2, C2, and D2) and interactive model terms (AB, AC, and AD) were highly significant (P < 0.05). However, the interactive model terms BC, BD, and CD did not depict significant effects on lipid yield (P > 0.05, data not shown). Applying multiple regression analysis, a second order polynomial equation was obtained, where the insignificant model terms were omitted. Thus, the mathematical regression model for lipid yield fitted in terms of coded factors is as follows:
Y (lipid yield) = +52.66 − 8.34 A − 7.69 B + 1.87 C + 1.77 D + 3.83 AB − 2.81 AC + 1.81 AD − 3.17 A2 − 5.81 B2 − 8.37 C2 − 8.73 D2
Table 4 presents the analysis of variance (ANOVA) with F- and P-values for the model. The P-value of the model was <0.0001, which indicated that the model was highly significant. A ‘lack of fit’ value of 3.73 was found to be insignificant relative to the pure error (0.0794, P > 0.05). The high values of the determination coefficient (R 2 = 0.9803) and the adjusted determination coefficient (adjusted R 2 = 0.9613) also showed that the model was highly significant.
Localization of the optimized condition
The fitted polynomial equation was expressed as three-dimensional surface plots to visualize the relationship between the response and the experimental levels of each factor used in this design, and also to demonstrate the interactions amongst the variables. The shape of the corresponding contour plot indicates whether the mutual interactions between the independent variables are significant or not. Elliptical nature of the contour in 3D-response surface graphs (Fig. 4a–c) depicted the mutual interactions between the independent variables were significant. Figure 4a explains the positive interaction between nitrate and phosphate at zero level of thiosulphate and time culture, where decrease in concentration of nitrate and phosphate contributed to increased lipid yield. Figure 4b explains the interaction of nitrate with thiosulphate, where with decreasing nitrate concentration and increasing thiosulphate level, an increase in lipid yield was evident. The interaction of nitrate and time culture at zero level of phosphate and thiosulphate was significant as visualized from Fig. 4c, where lipid yield was found to increase with decreasing level of nitrate but with time culture near the zero level. On the other hand, the interactive model terms BC (phosphate and thiosulphate), BD (phosphate and time culture), and CD (thiosulphate and time culture) were not shown graphically, since those interactions did not illustrate significant effects on lipid accumulation, which was reflected from the flat response surface and more parallel contour lines.

3D response surface: interactive effects of a varied nitrate and phosphate concentrations at 0.85 g l−1 thiosulphate and time culture of 10 days, b varied nitrate and thiosulphate concentration at 0.05 g l−1 phosphate and time culture of 10 days, and c varied nitrate concentration and time culture of 10 days at 0.05 g l−1 phosphate and 0.85 g l−1 thiosulphate
After knowing the possible direction for maximizing lipid accumulation, optimization was done using the ‘point optimization’ technique. A maximum lipid yield of 61.3% (dcw) was predicted at 0.04 g l−1 nitrate, 0.03 g l−1 phosphate, and 1.0 g l−1 thiosulphate for time culture of 8 days (Table 5).
Validation of the model
The accuracy of the model was validated with experiments done in triplicate under the aforementioned optimal condition. A lipid yield of 58.3% (dcw) was achieved, which confirmed the validity of the predicted model. The residuals from the least squares fit also play an important role in judging the model adequacy (Myers and Montgomery 2002). By constructing a normal probability plot of the residuals, a check was made for the normality assumption (Fig. 5a). The normality assumption was satisfied as the residual plot approximated along a straight line. Figure 5b presents a plot of residuals versus the predicted response. The general impression is that the residuals scatter randomly on the display, suggesting that the variance of the original observation is constant for all values of Y. Both the plots (Fig. 5a, b) were found satisfactory, thus depicting the reliability of the model. Furthermore, a lipid yield of 2.16 g l−1 was recorded in cultures pre-grown in glucose (1.5%)-supplemented N 11 medium, when subjected to the optimized condition at the second stage, the value almost 40-fold higher with respect to the control condition (Table 6).

Plot of a normal probability of internally studentized residuals and b internally studentized residuals vs. predicted response
Analysis of S. obliquus biodiesel
The saponification and acid values of the microalgal oil were 231.7 ± 9.4 and 48.1 ± 2.7 mg KOH g−1, respectively. The average molecular weight of the oil was 917 ± 21. A typical TLC profile of the fatty acid methyl ester (FAME) samples indicated that there was no discernible triglyceride or diglyceride left after 7 h of transesterification (data not shown). The gas chromatograph of biodiesel along with the mass spectrum of individual FAME is shown in Fig. 6. Peak area less than 0.1% were considered to be negligible. The FAME of the biodiesel and their relative quantities are presented in the inset of Fig. 6a. The sample consisted mainly of methyl palmitate and methyl oleate, which were almost 75% of the total FAME, while esters of other long chain fatty acids such as linoleic and linolenic acids were of 10.8% and 15%, respectively.

a Gas chromatography spectrum of FAME, mass spectra of b methyl palmitate, c methyl oleate, d methyl linolieate, and e methyl linolineate
Discussion
S. obliquus depicted a profound rise in lipid yield (43% dcw) under nitrate-deficient condition (Fig. 2a). This is well in agreement with the report of Shiflin and Chisholm (1981), where accumulation of fatty acid derivatives in 30 micro-algal species belonging to chlorophyceae and diatoms was observed under nitrogen starvation. The possible reason could be that under nitrogen deficiency/limitations the available nitrogen is utilized for synthesis of enzymes and essential cell structures. Any carbon dioxide subsequently fixed is therefore converted into carbohydrate or lipid rather than protein (Richardson et al. 1969). Limitation of phosphate also appeared to be a suitable stimulant for lipid accumulation (Fig. 2b). This confirms the earlier findings of Rhee (1978), where lipid accumulation was enhanced when growth was restricted due to unavailability of phosphate. Supplementation glucose though not found stimulatory for lipid accumulation on dry cell weight basis (Table 2), the total lipid yield was found to increase up to 585.9 mg l−1 in 1.5% glucose-supplemented vessel against 55.5 mg l−1 in the control one. A highly positive correlation (r = 0.93) between lipid content and biomass yield demonstrated that the rise in lipid yield in glucose-supplemented medium is due to the increased biomass growth. Sodium thiosulphate was also found to raise the lipid pool in Scenedesmus up to 30.4% (dcw, Fig. 2c), which agrees with the earlier observation of Feng et al. (2005), where accumulation of fatty acids in Chlorella sp. was increased by sodium thiosulphate supplementation. The possible reason could be that a reducing environment leads to increased pool size of NADH (Rao and Mutharasan 1988). Under such condition, acetyl-CoA could not enter the tricarboxylic acid cycle (TCA) cycle as the high concentrations of NADH inhibit the enzyme citrate synthase, one of the key enzymes of TCA cycle, leading to an increase in the pool of acetyl-CoA (Doi 1990). The later might be converted to malonyl-CoA, catalyzed by acetyl-CoA carboxylase (ACCase), the central carbon donor for fatty acid synthesis (Ohlrogge and Browse 1995). Thus, the significant enhancement in lipid accumulation in S. obliquus under limitations of nitrate and phosphate, and thiosulphate supplementation pointed towards the optimization of these factors to maximize the lipid yield. Among the critical factors, glucose was excluded, since glucose was not found to stimulate lipid pool of the test organism on dry cell weight basis (Table 2). Nevertheless, time culture was included in the optimization process as lipid accumulation was found to vary (Table 1).
The rotatable central composite design was used to get the optimum response (Table 3). In this study, the regression equation was obtained using the coded factors and results were fitted to a second order polynomial model. The model obtained for lipid production was adequate enough as depicted from the high F-value (53.29), insignificant lack of fit (probability > F = 0.0794) and R 2 close to unity (0.9803).
The results showed significant effects for all the model terms except BC, BD, and CD. According to the second order polynomial equation, the linear coefficient A (nitrate) and B (phosphate) showed negative effects on lipid yield, whereas C (thiosulphate) and D (time culture) exhibited positive impact. The contour plots in 3D response depicted the variation in lipid yield as a function of interaction of the variables. From Fig. 4a–c, it is concluded that decreasing nitrate and phosphate concentrations and increasing thiosulphate concentration and time culture resulted in rise of lipid content within the surface. The optimum conditions for maximum lipid yield were obtained as follows: 0.04 g l−1 nitrate, 0.03 g l−1 phosphate, and 1.0 g l−1 thiosulphate at 8 days for the predicted lipid yield of 61.3% (dcw). The experimental value (58.3% dcw) was well in agreement with the predicted one (Table 5).
In this study, lipid content of S. obliquus reached up to 58.3% of dry cell weight, which was ~5-fold higher than the control, only by manipulating the nutrient status. However, productivity of S. obliquus biomass was found to be affected under N- and P-limitations and thiosulphate supplementation. This necessitates a two-stage cultivation practice, where at the first stage S. obliquus cell must be grown under glucose-supplemented condition to reach a high cell density with regards to increased lipid accumulation (~10-fold increase in lipid yield on mg l−1 basis was observed under glucose supplementation, Table 2), followed by a second stage, as optimized in this study, for efficient lipid accumulation. Scenedesmus cells pre-grown in glucose (1.5%)-supplemented N 11 medium when subjected to the optimized condition, the lipid pool was boosted up to 2.16 g l−1, the value almost 40-fold higher with respect to the control condition (Table 6).
Polyunsaturated fatty acids with four or more double bonds are quite common in microalgal lipid. These bonds are susceptible to oxidation during storage, thus reduce the acceptability of microalgal oil for production of biodiesel (Chisti 2007). The GC-MS study (Fig. 6) demonstrates that the biodiesel from S. obliquus contains mainly saturated and mono-unsaturated fatty acids (~75% of the total fatty acyl methyl esters), which advocates its high oxidative stability. Thus, S. obliquus could be considered as a potential organism for biodiesel production.
References
Alcantara R, Amores J, Canoira L, Fidalgo E, Franco MJ, Navarro A (2000) Catalytic production of biodiesel from soybean oil, used frying oil and tallow. Biomass Bioenergy 18:515–527
Becker K, Francis G (2002) Biodiesel from Jatropha plantations on degraded land. A brief technical report. University of Hohenheim, Stuttgrart
Behrens PW, Kyle DJ (1996) Microalgae as a source of fatty acids. J Food Lipids 3:259–272
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Brown AC, Knights BA, Conway E (1969) Hydrocarbon content and its relationship to physiological state in the green alga Botryococcus braunii. Phytochemistry 8:543–547
Canakci M, Gerpen JV (2001) Biodiesel production from oil and fats with high free fatty acids. Trans ASAE 44:1429–1436
Chisti Y (2007) Biodiesel from microalgae. Biotechnol Adv 25:294–306
Dayananda C, Sarada R, Usha Rani M, Shamala TR, Ravishankar GA (2007) Autotrophic cultivation of Botryococcus braunii for the production of hydrocarbons and exopolysaccharides in various media. Biomass Bioenergy 31:87–93
Doi Y (1990) Microbial Polyesters. VCH, New York
Dorado MP, Ballesteros E, Almeida JA, Schellert C, Lohrlein HP, Krause R (2002) An alkali-catalyzed transesterification process for high free fatty acid waste oils. Trans ASAE 45:525–529
Feng F-Y, Yang W, Jiang G-Z, Xu Y-N, Kuang T-Y (2005) Enhancement of fatty acid production of Chlorella sp. (Chlorophyceae) by addition of glucose and sodium thiosulphate to culture medium. Process Biochem 40:1315–1318
Ghadge SV, Raheman H (2005) Process optimization for biodiesel production from mahua (Madhuca indica) oil using response surface methodology. Bioresour Technol 97:379–384
Haag AL (2007) Algae bloom again. Nature 447:520–521
Illman AM, Scragg AH, Shales SW (2000) Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzyme Microb Technol 27:631–635
Lang X, Dalai AK, Bakhshi NN, Reany MJ, Hertz PB (2001) Preparation and characterization of biodiesel from various bio-oils. Bioresour Technol 80:53–62
Liu Z-Y, Wang G-C, Zhou B-C (2008) Effect of iron on growth and lipid accumulation in Chlorella vulgaris. Bioresour Technol 99:4717–4722
Miao X, Wu Q (2004) High yield bio-oil production from fast pyrolysis by metabolic controlling of Chlorella protothecoides. J Biotechnol 110:85–93
Minowa T, Yokoyama S-Y, Kishimoto M, Okakurat T (1995) Oil production from algal cells of Dunaliella tertiolecta by direct thermochemical liquefaction. Fuel 74:1735–1738
Mittelbach M, Pokits B, Silberholz A (1992) Production and fuel properties of fatty acid methyl esters from used frying oil. In: Liquid Fuels from Renewable Resources: Proceedings of an Alternative Energy Conference, St. Joseph, Mich, ASAE, pp. 74–78
Montgomery DC (2001) Design and analysis of experiments, 5th edn. Wiley, New York
Myers RH, Montgomery DC (2002) Response surface methodology: process and product optimization using designed experiments, 2nd edn. Wiley, New York
Ohlrogge J, Browse J (1995) Lipid biosynthesis. Plant Cell 7:957–970
Rai LC, Mallick N, Singh JB, Kumar HD (1991) Physiological and biochemical characteristics of a copper tolerant and a wild type strain of Anabaena doliolum under copper stress. J Plant Physiol 138:68–74
Rao G, Mutharasan R (1988) Altered electron flow in a reducing environment in Clostridium acetobutilicum. Biotech Lett 10:129–132
Rhee GY (1978) Effect of N:P atomic ratio and nitrate limitation on algal growth, cell composition and nitrate uptake. Limnol Oceanogr 23:10–25
Richardson B, Orcutt DM, Schwertner HA, Martinez Cara L, Wickline Hazel E (1969) Effects of nitrogen limitation on the growth and composition of unicellular algae in continuous culture. Appl Micobiol 18:245–250
Shiflin NS, Chisholm SW (1981) Phytoplankton lipids: interspecific differences and effects of nitrate, silicate and light dark cycles. J Phycol 17:374–384
Soeder CJ, Bolze A (1981) Sulphate deficiency stimulates the release of dissolved organic matter in synchronus culture of Scenedesmus obliquus. Physiol Pl 52:233–238
Vicente G, Martınez M, Aracil J (2004) Integrated biodiesel production: a comparison of different homogeneous catalysts systems. Bioresour Technol 92:297–305
Xu H, Miao X, Wu Q (2006) High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J Biotechnol 126:499–507
Acknowledgements
The authors would like to thank Mr. Subhash Chand for his assistance and Mr. Shovon Mandal is thankful to the Indian Institute of Technology Kharagpur, India, for providing fellowship.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mandal, S., Mallick, N. Microalga Scenedesmus obliquus as a potential source for biodiesel production. Appl Microbiol Biotechnol 84, 281–291 (2009). https://doi.org/10.1007/s00253-009-1935-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-009-1935-6