
Abstract
Aims/hypothesis
We describe a novel model of insulin-deficient diabetes with a single copy of the gene encoding insulin 1 (Ins1) and no gene encoding insulin 2 (Ins2).
Materials and methods
We constructed five lines of mice: mice with two copies of Ins1 (NODIns1+/+,Ins2−/−), mice with a single copy of Ins1 (NODIns1+/−,Ins2−/−), mice with two copies of Ins2 (NODIns1−/−,Ins2+/+), mice with a single copy of Ins2 (NODIns1−/−,Ins2+/−) and NODIns1+/−,Ins2−/− mice with a transgene encoding B16:Ala proinsulin.
Results
By 10 weeks of age, all male NODIns1+/−,Ins2−/− mice were diabetic, whereas all female NODIns1+/−,Ins2−/− were not diabetic (p<0.0001). In contrast, neither male nor female NODIns1−/−,Ins2+/− with a single copy of Ins2 (rather than single copy of Ins1) developed early diabetes and no mice with two copies of either gene developed early diabetes. Islets of the diabetic male NODIns1+/−,Ins2−/− at this early age had no lymphocyte infiltration. Instead there was heterogeneous (between islet cells) weak staining for insulin. Although only male NODIns1+/−,Ins2−/− mice developed diabetes, both male and female NODIns1+/−,Ins2−/− mice had markedly decreased insulin content. In NODIns1+/+,Ins2−/−, there was also a significant decrease in insulin content, whereas NODIns1−/−,Ins2+/+ mice, and even NODIns1−/−,Ins2+/− mice, were normal. Male NODIns1+/−,Ins2−/− mice were completely rescued from diabetes by introduction of a transgene encoding proinsulin. On i.p. insulin tolerance testing, male mice had insulin resistance compared with female mice.
Conclusions/interpretation
These results suggest that Ins1 is a ‘defective gene’ relative to Ins2, and that the mouse lines created provide a novel model of sex-dimorphic insulin-deficient diabetes.
Similar content being viewed by others


Introduction
Insulin is synthesised by pancreatic beta cells and plays a predominant role in glucose homeostasis. Mice have two genes encoding insulin, insulin 1 (Ins1) on chromosome 19, and insulin 2 (Ins2) on chromosome 7. Ins1 differs from Ins2 by two amino acids at positions B9 and B29 and there are additional differences in the leader sequence and the connecting peptide. In addition, Ins1 lacks an intron present in Ins2. These structural features suggest that Ins1 was generated by an RNA-mediated duplication–transposition event involving a transcript of Ins2, which was reinserted into the genome (retroposon) [1].
In a previous study, we created Ins1 knock-out (Ins1-KO) and Ins2-KO mice on the nonobese diabetes (NOD) background. The Ins1-KO prevented the majority of progression to autoimmune type 1 diabetes, whereas the Ins2-KO accelerated the development of type 1 diabetes [2]. Similarly, Thebault-Baumont et al. have shown that Ins2-KO mice bred onto the NOD background develop accelerated insulitis and diabetes [3]. These studies have suggested that the role of each gene encoding insulin in autoimmune diabetes is different.
More recently, we have described double insulin KO (Ins1-KO and Ins2-KO) NOD mice with a mutated gene encoding preproinsulin (B16:Ala) that rescues the mice from metabolic diabetes [4]. These mice lacking a native B:9–23 sequence do not develop autoimmune diabetes. During evaluation and creation of these mice, we unexpectedly found that male NOD mice with the genotype Ins1+/−, Ins2−/− (NODIns1+/−,Ins2−/−) developed early onset of diabetes (<10 weeks of age). In this study, we describe the NODIns1+/−,Ins2−/− mice, a novel strain with non-autoimmune insulin-deficient diabetes.
Materials and methods
Mice
We constructed four lines of mice: (1) mice with two copies of Ins1 (NODIns1+/+,Ins2−/−; Ins2-KO); (2) mice with a single copy of Ins1 (NODIns1+/−,Ins2−/−); (3) mice with two copies of Ins2 (NODIns1−/−,Ins2+/+; Ins1-KO); and (4) mice with a single copy of Ins2 (NODIns1−/−,Ins2+/−). The mice were established by breeding the original insulin knock-outs kindly provided by J. Jami (Cochin and Saint Vincent de Paul Hospital, Paris, France) on to NOD/Bdc mice using marker-assisted congenic methods [2]. NODIns1+/−,Ins2−/− and NODIns1−/−,Ins2+/− were produced by mating (Ins1-KO×Ins2-KO)F1 with Ins2-KO and mating (Ins1-KO×Ins2-KO)F1 with Ins1-KO, respectively. NODIns1+/−,Ins2−/− with a mutated transgene encoding proinsulin (NODIns1+/−,Ins2−/−,Tg(+)) were constructed by mating with NODIns1+/+,Ins2−/− and NODIns1−/−,Ins2−/− with the transgene encoding B16:Ala proinsulin [4, 5]. Mice were housed in a pathogen-free animal colony at Barbara Davis Center for Childhood Diabetes with an approved protocol from the University of Colorado Health Sciences Center Animal Care and Use Committee. All mice had free access to tap water in an air-conditioned room (22–25°C) with a 12-h light–darkness cycle (06.00–18.00 h). In addition, we produced control strains with the genetic region of 129S1/SvImj mice surrounding Ins1 and Ins2 bred onto the NOD mice and these mice did not develop ‘early’ diabetes but typical insulitis associated with later onset of diabetes as reported [4].
Genotype analysis
Genomic DNA was extracted from mouse tails. The genotyping for the Ins1-knock-out gene, the Ins1-wild-type gene, the Ins2-knock-out gene and the Ins2-wild-type gene was performed by using PCR [5]. The PCR products were electrophoresed on 2% agarose gels and visualised by ethidium bromide staining.
Diagnoses of diabetes
Glucose was measured weekly with the FreeStyle blood glucose monitoring system (TheraSense, Alameda, CA, USA), and the mice were considered diabetic after two consecutive blood glucose values >13.9 mmol/l. After development of diabetes, the mice were killed immediately and the pancreas was fixed in 10% formalin to perform histological analysis.
Insulin autoantibody (IAA) assay
IAA was measured with a 96-well filtration plate micro-IAA assay as previously described [6] and expressed as an index. A value of 0.01 or greater is considered positive and exceeds the 99th percentile of normal controls.
Insulin content in pancreas
Mice were analysed for insulin content at the age of 4–5 weeks, and blood glucose was measured before and after overnight fasting to confirm lack of diabetes. Insulin was extracted with 4 ml acid–ethanol with an overnight incubation at 4°C. Insulin concentration of the supernatant after centrifuging and diluting (×1,000) was measured with an ELISA-based insulin kit (Mercodia, Uppsala, Sweden). Calculated insulin content was corrected for pancreatic weight or body weight.
Response to exogenous insulin
To detect insulin resistance, an insulin tolerance test (ITT) was performed by injecting human insulin (0.75 IU/kg) i.p. into overnight-fasted mice at the age of 10 weeks and blood glucose levels were measured at 0, 15, 30, 45 and 60 min.
Intraperitoneal glucose tolerance test (ipGTT)
To assess glucose tolerance in NODIns1−/−,Ins2+/+ and NODIns1−/−,Ins2+/−, an ipGTT (2 g glucose/kg body weight) was performed in overnight-fasted mice, and blood glucose levels were measured at 0, 30, 60, 90 and 120 min. We calculated the glucose AUCs according to the trapezoidal rule from the glucose measurements at baseline (0 min), 30, 60, 90 and 120 min.
Histology
The pancreata obtained from the mice were fixed in 10% formalin and paraffin-embedded. Paraffin-embedded tissue sections were stained with a monoclonal mouse anti-insulin antibody (Sigma, St Louis, MO, USA) followed by incubation with a peroxidase-labelled anti-mouse IgG antibody (DakoCytomation, Carpinteria, CA, USA), and also with a peroxidase-labelled broad-spectrum secondary antibody (Zymed/Invitrogen Corporation, Carlsbad, CA, USA) for peroxidase staining on adjacent sections. For immunofluorescence staining, the secondary antibody incubation took place with anti-guinea-pig AMCA (blue)-, anti-mouse Texas Red (red)-, and anti-rabbit Cy2 (green)-conjugated antibodies (Jackson ImmunoResearch, West Grove, PA, USA).
Statistics
Data are shown as means±SEM. Statistical analyses of insulin content, body weight, ITTs and ipGTTs were performed by Mann–Whitney’s U test. Survival curves were analysed with the log-rank test. Statistical tests used PRISM software (Graphpad, San Diego, CA, USA). p<0.05 was regarded as significant.
Results
We produced four lines of mice varying their number of genes encoding insulin and monitored the mice for the development of diabetes (Fig. 1). As shown in Fig. 1a, all male mice with only a single Ins1 gene (NODIns1+/−,Ins2−/−) developed diabetes (12 out of 12) by 10 weeks of age, whereas no male mice with a single Ins2 gene (NODIns1−/−,Ins2+/−) (0 out of 12) developed diabetes by this age (p<0.0001). In addition, no male NODIns1+/+,Ins2−/− and NODIns1−/−,Ins2+/+ had early-onset diabetes (0 out of 9 and 0 out of 18, respectively). No female mice developed early-onset diabetes, independently of the number of genes encoding insulin, even in female NODIns1+/−,Ins2−/− (0 out of 17, p<0.0001) (Fig. 1b).

Development of diabetes with differential Ins knock-outs. a Male NODIns1+/−,Ins2−/− (n=12), NODIns1+/+,Ins2−/− (n=9), NODIns1−/−,Ins2+/− (n=12), NODIns1−/−,Ins2+/+ (n=18); b female NODIns1+/−,Ins2−/− (n=17), NODIns1+/+,Ins2−/− (n=8), NODIns1−/−,Ins2+/− (n=14), NODIns1−/−,Ins2+/+ (n=15). By 10 weeks of age, only male NODIns1+/−,Ins2−/− developed diabetes (12 out of 12). p<0.0001 (log-rank test) for survival curves between male NODIns1+/−,Ins2−/− and female NODIns1+/−,Ins2−/−, and between male NODIns1+/−,Ins2−/− and male NODIns1−/−,Ins2+/−
After 10 weeks of age, female Ins2-KO (NODIns1+/+,Ins2−/− and NODIns1+/−,Ins2−/−) mice had a high prevalence of diabetes (Fig. 1b) and severe insulitis. However, no Ins1-KO (NODIns1−/−,Ins2+/+ and NODIns1−/−,Ins2+/−) mice in either the males or females developed diabetes until 36 weeks of age (Fig. 1).
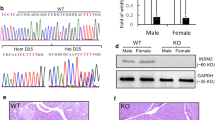
Since male NOD mice with a single copy of Ins1 and lacking Ins2 developed diabetes, we analysed the histology of the male NODIns1+/−,Ins2−/− mice with diabetes. Despite the presence of diabetes, islets on haematoxylin–eosin staining appeared normal and there was no lymphocytic infiltration (Fig. 2a). We also measured IAA (Fig. 2b). The level of IAA in male NODIns1+/−,Ins2−/− mice was the same as in wild-type male NOD and less than male NODIns1+/+,Ins2−/−. A sex-related difference in IAA expression for mice <10 weeks of age was not detected in NODIns1+/−,Ins2−/− mice. These results suggest that diabetes in male NODIns1+/−,Ins2−/−, which develops prior to 10 weeks of age, is not associated with enhanced autoimmunity, especially the lack of islet infiltrates.

a Histology of a diabetic NODIns1+/−,Ins2−/− mouse with diabetes that developed before 10 weeks of age (haematoxylin–eosin staining). b–d Longitudinal analysis of IAA. The level of IAA in male (M) NODIns1+/−,Ins2−/− mice was the same as in wild-type male NOD and less than male NODIns1+/+,Ins2−/−. A sex-related difference in IAA before 10 weeks of age was not detected in NODIns1+/−,Ins2−/− mice. F Female. Horizontal line, cut-off for mIAA (0.01)
We next measured pancreatic insulin content of mice at 4–5 weeks of age, before the development of diabetes (Fig. 3). The insulin content in male NODIns1+/−,Ins2−/− was significantly lower than that in mice with any other insulin genotype corrected for pancreatic weight (Fig. 3a) or body weight (Fig. 3b). In marked contrast, NODIns1−/−,Ins2+/− mice, with only one copy of the Ins2 gene, had almost the same insulin content as wild-type NOD insulin mice or NODIns1−/−,Ins2+/+ mice, suggesting NODIns1−/−,Ins2+/− mice compensate for loss of genes while NODIns1+/−,Ins2−/− mice cannot. NODIns1+/+,Ins2−/− mice had an intermediate insulin content compared with lower levels for NODIns1+/−,Ins2−/− and NODIns1−/−,Ins2+/− mice (Fig. 3c). Thus a single copy of Ins2 results in greater insulin content than two copies of Ins1. In addition, as shown in Fig. 3, at this age, Ins2-KO mice (including NODIns1+/−,Ins2−/−, NODIns1+/+,Ins2−/−) weighed slightly less compared with Ins1-KO (including NODIns1−/−,Ins2+/−, NODIns1−/−,Ins2+/+) (p<0.05 in both male and female mice), and wild-type NOD (p<0.05 in male and p<0.01 in female mice). However, the body weight of Ins1-KO mice with Ins2 was not significantly different from that of wild-type NOD mice, suggesting that low pancreatic insulin was associated with delayed growth and/or fat accumulation.

a,b Pancreatic insulin content of NODIns1+/−,Ins2−/− (n=8, 6, male and female, respectively), NODIns1+/+,Ins2−/− (n=5, 6), NODIns1−/−,Ins2+/− (n=8, 9), NODIns1−/−,Ins2+/+ (n=8, 8) and wild-type NOD mice (n=7, 7). Filled bars, male mice; open bars, female mice. Insulin content was per milligram of pancreatic weight (a) or gram of body weight (BW) (b). NODIns1+/−,Ins2−/− had extremely low pancreatic insulin content. No sex-related difference in pancreatic insulin content was detected. c Body weight of Ins2-KO (combining NODIns1+/−,Ins2−/− and NODIns1+/+,Ins2−/− [n=13, 12, male and female, respectively]), Ins1-KO (combining NODIns1−/−,Ins2+/− and NODIns1−/−,Ins2+/+ [n=16, 17]) and wild-type NOD mice (n=7, 7). Black bars, Ins2-KO mice; grey bars, Ins1-KO mice; open bars, wild-type NOD mice. *p<0.05, **p<0.01, # p<0.0001 combined male and female
Given the low content of insulin at a young age before diabetes, we evaluated insulin expression by islets with immunocytochemistry (Fig. 4). Compared with wild-type NOD mice, mice with a single copy of Ins1 had weak and heterogeneous (between beta cells) insulin expression. These results suggest that the diabetes of male NODIns1+/−,Ins2−/− results from deficient insulin expression. To confirm this, we genetically rescued male NODIns1+/−,Ins2−/− mice from diabetes with a transgene encoding proinsulin that we had previously developed to study a specific epitope of insulin (B:9–23). This transgene, driven off the rat insulin promoter, produces biologically active insulin despite a single amino acid change (B16:Ala). Male NODIns1+/−,Ins2−/− mice with the transgene (n=17) did not develop early onset of diabetes (p<0.0001; log-rank test). In addition, mice with the transgene recovered pancreatic insulin content at least to the same level as NODIns1+/+,Ins2−/− (Fig. 5). These results strongly suggest that rescue from diabetes was possible with the introduction of insulin message independently of the wild-type insulin loci and that male NODIns1+/−,Ins2−/− developed diabetes because of low insulin expression.

No insulitis and weak and heterogeneous staining for insulin in male NODIns1+/−,Ins2−/− islets with insulin immunoperoxidase staining (a) and insulin immunofluorescence staining (e) despite the presence of normal islet architecture. Female NODIns1+/−,Ins2−/− (b,f) had similar signal levels to male NODIns1+/−,Ins2−/−, whereas male NODIns1−/−,Ins2+/− (c, g), and wild-type NOD (d, h) had more uniform and brighter insulin staining compared with male NODIns1+/−,Ins2−/−. All samples were collected at 5–6 weeks of age before development of diabetes. a–d Insulin immunoperoxidase staining (brown: insulin). e–h Insulin immunofluorescence staining (blue: insulin, red: glucagon, green: somatostatin)

Pancreatic insulin content of NODIns1+/−,Ins2−/−,Tg(−) (n=8, 6, male and female, respectively), NODIns1+/−,Ins2−/−,Tg(+) (n=11, 11), and NODIns1+/+,Ins2−/−,Tg(−) (n=5, 6). Pancreatic insulin content in the NODIns1+/−,Ins2−/−,Tg(+) was significantly higher than that in the NODIns1+/−,Ins2−/−,Tg(−) (# p<0.0001) and similar to that in the NODIns1+/+,Ins2−/−,Tg(−) (p=0.28), suggesting that insulin content in the pancreas was rescued by the transgene encoding B16:Ala mutated proinsulin. Filled bars, male mice; open bars, female mice
As shown in Fig. 3, insulin content in male and female NODIns1+/−,Ins2−/− mice was similar, although only male mice developed diabetes. We hypothesised that male NOD mice might be more insulin-resistant than female NOD mice. Therefore we evaluated the glucose response to i.p. insulin in male and female NOD mice (Fig. 6). Male NOD mice had significantly higher blood glucose than female mice (Fig. 6a), and the blood glucose at 15 min (%basal) after i.p. insulin was also significantly higher, and the slope of glucose decline was decreased compared with female mice (Fig. 6b). In addition, as shown in Fig. 7, even among male and female NODIns1−/−,Ins2+/+ that did not develop any diabetes and had no differences in pancreatic insulin content, the male mice have higher glucose on an ipGTT than females. These results suggest that sex differences in insulin resistance may contribute to the metabolic differences between male and female knock-out mice.

ITTs in NOD mice at 10 weeks of age. Filled squares, male mice (n=10); filled circles, female NOD mice (n=12). Male NOD mice had significantly higher blood glucose than female (*p<0.05, **p<0.01) (a) and the blood glucose at 15 min (percentage of basal) was also significantly higher than that in the female (p<0.05) (b)

a,b Intraperitoneal GTTs in male NODIns1−/−,Ins2+/+ (n=10, filled squares) mice, female NODIns1−/−,Ins2+/+ (n=6, filled circles) mice, and male NODIns1−/−,Ins2+/− (n=3, open squares) mice at 10 weeks of age. *p<0.05, # p<0.005 compared with male NODIns1−/−,Ins2+/+
Discussion
In this study, we report that male NOD mice with only a single copy of Ins1 (NODIns1+/−,Ins2−/−) develop ‘metabolic diabetes’ at less than 10 weeks of age. The pathogenesis is probably a defect of insulin production with low and heterogeneous expression of insulin that can be corrected by additional transgenic expression of proinsulin. A single copy of Ins2 gene is sufficient to prevent diabetes and is associated with normal insulin content. This suggests that Ins1 gene is a functionally defective gene relative to Ins2 gene and interestingly, there is no compensation sufficient to prevent diabetes with only a single Ins1 gene.
It has been reported that Ins1-KO and Ins2-KO, when on a C57BL/6 strain background and with two copies of each gene encoding insulin, have insulin content in the pancreas similar to wild-type mice, in addition to normal glucose tolerance, because of the compensatory response of insulin transcription [7, 8]. In contrast to these studies, our NODIns1+/+,Ins2−/− mice have lower insulin content compared with wild-type NOD mice. A major difference between these studies may be related to the background genome, namely NOD background in our study vs C57BL/6 background. We are surprised that there is not compensation for insulin deficiency with the single copy of Ins1 and with two copies of Ins1. However, a strain derived from the same colony as the NOD mouse, the Nagoya–Shibata–Yasuda mouse [9, 10], a model of type 2 diabetes, which may share background genome with NOD [11, 12], has no compensational hypertrophy of pancreatic islets despite increasing insulin resistance with ageing [9]. In addition, Kulkarni et al. reported the importance of background genome in the induction of type 2 diabetes [13].
Male and female NODIns1+/−,Ins2−/− mice had similarly decreased levels of pancreatic insulin content in young non-diabetic mice. However, diabetes incidence between male and female NODIns1+/−,Ins2−/− mice was significantly different. We hypothesise that small but statistically significant differences in response to insulin as evidenced by our i.p. ITT may relate to sex differences in development of overt diabetes of these insulin-deficient NODIns1+/−,Ins2−/− mice. We believe that greater insulin resistance for male mice with marginal insulin production leads to hyperglycaemia. In fact, most animal models of type 2 diabetes (including Nagoya–Shibata–Yasuda mice) show a high prevalence of type 2 diabetes in male mice [9, 10].
In humans, a polymorphism upstream of the INS promoter correlates with insulin expression in the thymus, but has little effect on pancreatic insulin expression [14, 15]. Therefore, loci other than the insulin locus may contribute to pancreatic insulin expression. Further analysis of crosses of NOD and B6 mice with insulin gene knock-outs may help clarify such genetic loci that possibly relate to regulation of insulin production. Of note, humans have only the insulin 2 homologous gene.
Analysing structural features of the gene encoding insulin, Soares et al. reported that Ins1 was generated by an RNA-mediated duplication–transposition event involving a transcript of Ins2 [1]. In addition, only Ins2 is expressed in the NOD thymus with both genes expressed in islet beta cells [3, 16, 17]. This study clearly indicates that Ins1 (on the NOD genetic background) is a functionally defective gene compared with Ins2, and provides a novel model of sex-dimorphic insulin-deficient diabetes.
Abbreviations
- IAA:
-
insulin autoantibody
- Ins1 :
-
gene encoding insulin 1
- Ins2 :
-
gene encoding insulin 2
- ipGTT:
-
intraperitoneal glucose tolerance test
- ITT:
-
insulin tolerance test
- KO:
-
knock-out
- NOD:
-
nonobese diabetic
References
Soares MB, Schon E, Henderson A et al (1985) RNA-mediated gene duplication: the rat preproinsulin I gene is a functional retroposon. Mol Cell Biol 5:2090–2103
Moriyama H, Abiru N, Paronen J et al (2003) Evidence for a primary islet autoantigen (preproinsulin 1) for insulitis and diabetes in the nonobese diabetic mouse. Proc Natl Acad Sci USA 100:10376–10381
Thebault-Baumont K, Dubois-Laforgue D, Krief P et al (2003) Acceleration of type 1 diabetes mellitus in proinsulin 2-deficient NOD mice. J Clin Invest 111:851–857
Nakayama M, Abiru N, Moriyama H et al (2005) Prime role for an insulin epitope in the development of type 1 diabetes in NOD mice. Nature 435:220–223
Nakayama M, Moriyama H, Abiru N et al (2004) Establishment of native insulin-negative NOD mice and the methodology to distinguish specific insulin knockout genotypes and a B:16 alanine preproinsulin transgene. Ann N Y Acad Sci 1037:193–198
Yu L, Robles DT, Abiru N et al (2000) Early expression of antiinsulin autoantibodies of humans and the NOD mouse: evidence for early determination of subsequent diabetes. Proc Natl Acad Sci USA 97:1701–1706
Leroux L, Desbois P, Lamotte L et al (2001) Compensatory responses in mice carrying a null mutation for Ins1 or Ins2. Diabetes 50(Suppl 1):S150–S153
Chentoufi AA, Polychronakos C (2002) Insulin expression levels in the thymus modulate insulin-specific autoreactive T-cell tolerance: the mechanism by which the IDDM2 locus may predispose to diabetes. Diabetes 51:1383–1390
Ueda H, Ikegami H, Yamato E et al (1995) The NSY mouse: a new animal model of spontaneous NIDDM with moderate obesity. Diabetologia 38:503–508
Rees DA, Alcolado JC (2005) Animal models of diabetes mellitus. Diabet Med 22:359–370
Ikegami H, Fujisawa T, Ogihara T (2004) Mouse models of type 1 and type 2 diabetes derived from the same closed colony: genetic susceptibility shared between two types of diabetes. ILAR J 45:268–277
Babaya N, Ikegami H, Fujisawa T et al (2005) Susceptibility to streptozotocin-induced diabetes is mapped to mouse chromosome 11. Biochem Biophys Res Commun 328:158–164
Kulkarni RN, Almind K, Goren HJ et al (2003) Impact of genetic background on development of hyperinsulinemia and diabetes in insulin receptor/insulin receptor substrate-1 double heterozygous mice. Diabetes 52:1528–1534
Pugliese A, Zeller M, Fernandez A Jr et al (1997) The insulin gene is transcribed in the human thymus and transcription levels correlated with allelic variation at the INS VNTR-IDDM2 susceptibility locus for type 1 diabetes. Nat Genet 15:293–297
Vafiadis P, Bennett ST, Todd JA et al (1997) Insulin expression in human thymus is modulated by INS VNTR alleles at the IDDM2 locus. Nat Genet 15:289–292
Chen W, Bergerot I, Elliott JF et al (2001) Evidence that a peptide spanning the B–C junction of proinsulin is an early autoantigen epitope in the pathogenesis of type 1 diabetes. J Immunol 167:4926–4935
Brimnes MK, Jensen T, Jorgensen TN, Michelsen BK, Troelsen J, Werdelin O (2002) Low expression of insulin in the thymus of non-obese diabetic mice. J Autoimmun 19:203–213
Acknowledgements
This work was supported by grants from the National Institutes of Health (DK32083, DK55969, DK62718), Diabetes Endocrine Research Center (P30 DK57516), the American Diabetes Association, the Juvenile Diabetes Foundation, and the Children’s Diabetes Foundation. N. Babaya was supported by the Osaka Medical Research Foundation for Incurable Diseases. M.Nakayama was supported by a fellowship from the Juvenile Diabetes Foundation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Babaya, N., Nakayama, M., Moriyama, H. et al. A new model of insulin-deficient diabetes: male NOD mice with a single copy of Ins1 and no Ins2 . Diabetologia 49, 1222–1228 (2006). https://doi.org/10.1007/s00125-006-0241-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-006-0241-4