
Abstract
Aims/hypothesis
The aim of this study was to investigate whether dynamic strength training modifies the control of lipolysis, with particular attention paid to the involvement of the antilipolytic adrenergic alpha 2A receptor (ADRA2A) pathway.
Methods
Twelve obese men (age: 47.4±2.8 years; BMI: 32.7±0.9) were investigated during a 210-min euglycaemic–hyperinsulinaemic clamp conducted before and after 3 months of dynamic strength training. Before and during the third hour of the clamp, the lipolytic effect of a perfusion of isoproterenol or adrenaline (epinephrine) alone or associated with the ADRA2A antagonist phentolamine was evaluated using the microdialysis method of measuring extracellular glycerol concentration (EGC) in subcutaneous abdominal adipose tissue (SCAAT). In addition, biopsies of SCAAT were carried out before and after training to determine mRNA levels
Results
The training increased insulin sensitivity in adipose tissue. The decrease of EGC was more pronounced during the clamp conducted after the training period than during the clamp done in pre-training conditions. Before and after the training, catecholamines induced an increase in EGC, the increase being lower during the clamp on each occasion. The isoproterenol-induced increase in EGC was higher after the training. Adrenaline-induced lipolysis was potentiated by phentolamine after but not before the training. There were no training-induced changes in mRNA levels of key genes of the lipolytic pathway in SCAAT.
Conclusions/interpretation
In obese subjects, dynamic strength training improves whole-body and adipose tissue insulin responsiveness. It increases responsiveness to the adrenergic beta receptor stimulation of lipolysis and to the antilipolytic action of catecholamines mediated by ADRA2As.
Similar content being viewed by others


Introduction
Adrenergic receptor subtypes control lipolysis in human adipose tissue. Studies in isolated human fat cells have shown that the activation of adrenergic alpha 2A receptor (ADRA2A) by adrenaline (epinephrine) and noradrenaline (norepinephrine) impairs the beta-adrenergic component of catecholamine-induced lipolysis. The nature of the interplay between the two pathways has been clearly established in vitro. In human subcutaneous fat cells, where ADRA2As outnumber adrenergic beta receptors, preferential recruitment of the ADRA2A leads to inhibition of lipolysis [1, 2]. Physiological and pathological studies have shown that the possible deregulation of adrenergic control in adipose tissue lipolysis in obese subjects is linked to an alteration of the beta-adrenergic pathway. Although less investigated, ADRA2A has also been shown to contribute to lipolysis dysfunction [3].
Adipose tissue is a major target for insulin effects. Insulin inhibits lipolysis and counters the action of adrenergic beta receptors by activating phosphodiesterase-3B, which hydrolyses cyclic AMP and also desensitises beta-adrenergic responsiveness [4]. Microdialysis allows the in situ study of the interaction of insulin with the adrenergic pathways in subcutaneous adipose tissue. When delivered through microdialysis probes, catecholamines promote stimulation of lipolysis in adipose tissue without affecting circulating catecholamine concentrations. Insulin counteracts the beta-adrenergic lipolytic effect in adipose tissue [5]. In previous studies in healthy normal-weight individuals, we have shown a decrease in the lipolytic response to adrenaline during a hyperinsulinaemic–euglycaemic clamp in human subcutaneous abdominal adipose tissue (SCAAT) [6]. Several mechanisms have been proposed to explain the lipolytic changes promoted by insulin administration. The adrenergic-beta-receptor-dependent lipolytic pathway is reduced, presumably through the activation of phosphodiesterase-3B, leading to cAMP reduction [7, 8]. In addition, the concomitant activation of the antilipolytic ADRA2A-dependent pathway by adrenaline also contributes to the reduction of the adrenergic-beta-receptor-dependent lipolytic response [6]. These studies [6–8] demonstrated that insulin regulates lipolysis in human subcutaneous adipose tissue by affecting the functional balance between the adrenergic beta and ADRA2A-dependent pathways.
The present study was performed to expand our in vivo investigations of the mechanisms involved in the control of lipid mobilisation in SCAAT in obese subjects. Insulin resistance is common in obese patients and increasing insulin sensitivity has become a major target outcome for clinical trials examining lifestyle modifications including physical activity and diet. The interplay between adrenergic beta receptor and ADRA2A pathways has never been investigated, using in situ microdialysis, in insulin-resistant obese patients. Since aerobic training is known to improve metabolic parameters in insulin-resistant individuals, we investigated whether another popular training protocol, dynamic strength training, improves insulin responsiveness and modifies the interplay between the adrenergic beta receptor and ADRA2A pathways in insulin-resistant obese subjects. Thus the main aims of this study were to investigate: (1) the effect of dynamic strength training on lipolysis regulation; and (2) the effect of this type of training on insulin resistance as evaluated using an insulin-clamp technique.
Based on in situ microdialysis use in SCAAT, we investigated the counteracting effect of clamp-induced hyperinsulinaemia on the lipolytic action of isoproterenol, a selective adrenergic beta receptor agonist or adrenaline, a catecholamine exhibiting both ADRA2A and adrenergic beta receptor agonist properties, in obese subjects with insulin resistance. The second objective was to evaluate the effect of a 3-month dynamic strength training programme on adipose tissue insulin responsiveness and the interplay between the adrenergic beta receptor and ADRA2A pathways in the control of lipid mobilisation. In addition, we investigated the expression of key genes in lipolysis regulation. This was done in biopsy samples of SCAAT by quantifying mRNA levels with reverse transcription-quantitative PCR (RT-qPCR).
Subjects, materials and methods
Subjects
For the study we recruited 12 obese men, who were aged 47.4±2.8 years (range 28–63 years), had a mean BMI of 32.7±0.9 kg/m2 (range 30.7–39.1), and who had not been submitted to any pharmacological or nutritional protocol prior to our study. All had had a stable weight during the previous 3 months. Selection of the subjects was based on detailed medical history, a physical examination, and several blood chemistry analyses. The Ethical Committee of the Third Faculty of Medicine, Charles University, Prague approved the study. All subjects gave their informed consent after detailed explanation of the experimental procedure.
Training programme
Participants exercised in a bodybuilding gym for 12 weeks. Exercise sessions were three times a week, and lasted 1 h, including warm-up and stretching. The dynamic training was performed using equipment (Technogym, Gambettola, Italy) arranged in a circuit (different multi-joint and single-joint exercises, e.g.: bench press, leg press, chest press, chest crossover, lateral pull-down, back extension, abdominal curl, triceps extension, biceps curl, leg extension, leg curl, toe raise, lateral raise). The subjects completed one set of the circuit during the initial week, passing on to two sets gradually after 3–4 weeks. The prescription of the dynamic strength training was performed according to conventional guidelines [9] The intensity was set at 60–70% of the one-repetition maximum strength (1-RM) for each exercise (i.e. for each muscle group), and 12–15 repetitions were performed. The work-load was adjusted to the increasing maximal strength throughout the training period in order to maintain the intensity at 60–70% of 1-RM. Participants were supervised by professional fitness instructors, initially twice a week, and later on once a week. All participants kept an exercise diary.
Experimental protocol
Studies were performed on two separate occasions (before and after 3 months of dynamic strength training). During this period, patients were instructed to maintain their nutritional habits. Four to five days before the investigation, each subject did a maximum exercise test on a bicycle ergometer (Ergometrics er800S; Ergoline, Bitz, Germany) to determine peak oxygen consumption using a Vmax (Sensor Medics, Yorba Linda, CA, USA). This test began with an initial workload of 60 W, followed by a sequential increase in workload of 30 W every 3 min until exhaustion. Verbal encouragement was given to attain maximal performance. Heart rate was continuously monitored. The second of the two investigations took place 48–72 h after the last exercise session in order to eliminate the possible acute effect of that session on the variables investigated.
On the day of the experiment, the subjects, who had fasted overnight, came to the hospital at 08:00 hours. During the experimental period they remained in a semi-recumbent position. An indwelling polyethylene catheter was inserted into the antecubital vein of each arm. At 08:30 hours four microdialysis probes (Carnegie Medicine, Stockholm, Sweden) of 20 × 0.5 mm and 20,000 molecular weight cut-off were inserted percutaneously into the abdominal SCAAT after epidermal anaesthesia (200 μl of 1% lidocaine; Roger-Bellon, Neuilly-s-Seine, France). The probes were separated by at least 5 cm, and inserted at a distance of 10 cm to the right or the left of the umbilicus. They were connected to a micro-perfusion pump (Harvard Apparatus, Les Ulis, France) and infused with Ringer’s solution (139 mmol/l sodium, 2.7 mmol/l potassium, 0.9 mmol/l calcium, 140.5 mmol/l chloride, 2.4 mmol/l bicarbonate). The extracellular glycerol concentration (EGC) found in the present study fits with previous determinations performed in obese subjects [3, 10]. Ethanol was added to the perfusate to estimate changes in the blood flow, as previously described [11–13]. The ethanol ratio was calculated as: ethanol ratio (%)=(ethanol concentration in outgoing dialysate/ethanol concentration in ingoing perfusate)×100. Variations of the ethanol ratio were taken as an index of variations of adipose tissue blood flow. The outgoing dialysate was collected throughout the experiment in all the probes to measure EGC. The scheme of the perfusions in the four microdialysis probes in relation to the hyperinsulinaemic clamp is depicted in Fig. 1. After the equilibration period, all the probes were perfused for 30 min with Ringer solution. During the following 30 min probe 1 was perfused with isoproterenol (1 μmol/l) in Ringer solution, probe 2 with adrenaline (10 μmol/l) in Ringer solution, probe 3 with adrenaline and phentolamine (100 μmol/l) in Ringer solution and probe 4 (control) with Ringer solution. Thereafter, the hyperinsulinaemic–euglycaemic clamp was started and all the probes were perfused with Ringer solution. At 150 min after initiation of the clamp (210 min after beginning of the protocol), the perfusions using adrenaline, isoproterenol and adrenaline plus phentolamine were performed again with concentrations identical to those before the clamp (Fig. 1). As previously shown, this time lapse between the infusions is necessary in order to avoid desensitisation of catecholamine-induced lipolysis [6]. Two days after this investigation, at 09:00 hours, a needle microbiopsy (200–300 mg) of adipose tissue was performed under local anaesthesia (1% xylocaine) 10–15 cm from the umbilicus for mRNA evaluation of adrenergic beta 2 receptor (ADRB2), ADRA2A, hormone-sensitive lipase and phosphodiesterase-3B (PDE3B).

Scheme of the experimental protocol: time course of microdialysis perfusions and hyperinsulinaemic clamp
Euglycaemic–hyperinsulinaemic clamp
The euglycaemic–hyperinsulinaemic clamp was performed according to the de Fronzo method [14]. A catheter for insulin and glucose infusion was inserted into an antecubital vein and a second catheter for blood sampling was placed in a dorsal vein of the ipsilateral hand. The hand was kept in a warm box (60°C) to provide arterialisation of venous blood. Priming plus continuous infusion of crystalline human insulin (Actrapid Human; Novo, Bagsvaerd, Denmark), 40 mU m2 min−1, was given for 210 min. Euglycaemia (the fasting blood glucose concentration) was maintained by a variable 20% glucose perfusion. The perfusion rate was determined by measuring arterialised plasma glucose at 5-min intervals using a glucose analyser (Beckman Instruments, Fullerton, CA, USA). Plasma concentrations of glucose, glycerol, NEFA and free insulin were analysed before and during the clamp.
Quantification of messenger RNAs in human adipose tissue biopsies using RT-qPCR
Total RNA was extracted from adipose tissue biopsies as previously described [15]. RT-qPCR was performed as described [15], except that first-strand cDNAs were first synthesised from 200 ng total RNA instead of 1 μg. For ADRA2A, PDE3B and hormone-sensitive lipase a set of primers (Genset-Proligo, Paris, France) was designed using Primer Express 1.5 (Applied Biosystems, Foster City, CA, USA) and used at a final AR concentration of 900 nmol/l with SYBR-Green-based chemistry. For ADRB2, Taqman was used. Both primers and TaqMan probes were obtained from Applied Biosystem. We used 18S rRNA as control to normalise gene expression using the Ribosomal RNA Control TaqMan Assay kit. All reactions were performed in duplicate.
Drugs and biochemical determinations
The following drugs were used: isoproterenol hydrochloride (Abbot, Rungis, France), adrenaline hydrochloride (Stallergenes, Antony, France) and phentolamine methanesulfonate (Novartis, Basel, Switzerland). Ethanol in the dialysate and perfusate was determined with an enzymatic method [16]. Glycerol was determined in plasma and in the dialysate using an ultrasensitive radiometric method [17]. Plasma glucose was assayed with a glucose oxidase technique (Biotrol, Paris, France). NEFA were assayed with an enzymatic method (Unipath, Dardilly, France). Plasma insulin was measured using an Insulin Irma kit (Immunotech, Prague, Czech Republic).
Data analysis
All values are given as means±SEM. A statistical comparison of the values was performed using the non-parametric Wilcoxon’s test for paired data. Values were considered statistically significant when p<0.05. Statistical analyses were performed using software packages (Statview 4.5 and SuperAnova 1.11; Abacus Concepts, Berkeley, CA, USA).
Results
Biological and anthropometric data before and after dynamic strength training
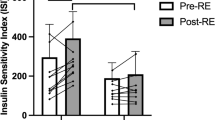
Table 1 reports the baseline characteristics of the patients and the changes observed after training. The main changes induced by dynamic strength training were the reduction in plasma leptin concentration and the increase in insulin sensitivity as assessed by homeostasis model assessment for insulin resistance or the insulin sensitivity coefficient (SIClamp) calculated according to Katz et al. [18].
Euglycaemic–hyperinsulinaemic clamp
Insulin infusion induced a steady-state insulin level (83.4±6.8 mU/l before training and 85.4±7.9 mU/l after training) with average individual insulin level variations of 8.6±3.1%. The subjects were clamped at their individual fasting glucose levels. The coefficient of variation for the glucose levels during the clamp was (5.67±0.75% before training vs 4.67±0.63% after training). Glucose consumption, calculated from the exogenous glucose infusion rates during the clamp, was significantly increased after the training period (Table 1).
Effect of training on gene expression
We quantified mRNA levels in SCAAT for key genes involved in lipolysis regulation in adipose tissue, i.e. ADRA2A, ADRB2, hormone-sensitive lipase and PDE3B. This was done by real-time RT-qPCR in samples taken from subjects before and at the end of the training period. The training did not induce changes in the gene expression profile (Table 1).
Extracellular glycerol, plasma glycerol and NEFA concentrations during the euglycaemic–hyperinsulinaemic clamp
The EGC before and during the clamp was evaluated in the probe infused with Ringer solution. EGC was higher before than after the training period (Fig. 2a). The decrease of EGC (Fig. 2b) during the insulin clamp was more pronounced after training. Basal plasma NEFA concentrations were significantly higher after the training programme than before (387±47 vs 538±63 μmol/l respectively) (Fig. 3a) while no changes were observed for plasma glycerol levels (98±13 vs 110±7 μmol/l respectively) (Fig. 3c). The relative decreases in NEFA and glycerol concentrations during the insulin clamp were more pronounced after dynamic strength training (Fig. 3b,d).

Extracellular glycerol concentration in subcutaneous adipose tissue during the euglycaemic–hyperinsulinaemic clamp before (filled bars) and after (open bars) dynamic strength training in obese subjects with insulin resistance. a Absolute values of glycerol. b Changes in glycerol values during the insulin clamp. Calculations of % variation were performed versus basal values of samples (15–30 min). Values are means±SEM. * p<0.05 when compared to values measured before dynamic strength training

Plasma NEFA and glycerol concentrations during the euglycaemic–hyperinsulinaemic clamp before (filled bars) and after (open bars) dynamic strength training in obese subjects with insulin resistance. a, b Absolute values of plasma NEFA and glycerol concentrations during the insulin clamp. c, d Change in plasma NEFA and glycerol concentrations during the insulin clamp. Calculations of % variation were performed versus basal values of samples (30 min). Values are means±SEM. * p<0.05 when compared to values measured before dynamic strength training
Lipolytic response to isoproterenol and adrenaline (alone or associated with phentolamine) perfusion in subcutaneous adipose tissue during hyperinsulinemic–euglycaemic clamp: effect of dynamic strength training
In the probe perfused with isoproterenol as well as that perfused with adrenaline, basal EGC was lower after training than before (150±19 vs 287±47 μmol/l and 149±27 vs 331±53 μmol/l respectively) (Figs. 4 and 5). Perfusion for 30 min with isoproterenol or adrenaline in the probes induced an increase in EGC (Figs. 4 and 5). Immediately after the end of the first perfusion, the euglycaemic–hyperinsulinaemic clamp was started. Perfusions of identical isoproterenol or adrenaline concentrations during the third hour of insulin clamp induced markedly lower EGC increases than those before the clamp (Figs. 4 and 5). Isoproterenol- or adrenaline-induced increases in EGC were reduced by 18 and 36%, respectively, during the insulin clamp.

In situ effects of isoproterenol (Iso, 1 μmol/l) on extracellular glycerol concentration in subcutaneous abdominal adipose tissue during the euglycaemic–hyperinsulinaemic clamp, before (diamonds) and after (squares) dynamic strength training in obese subjects with insulin resistance. a Absolute values of extracellular glycerol concentration. b Changes in extracellular glycerol concentration (calculated as: mean glycerol value during the 30-min perfusion minus mean glycerol values from two 15-min fractions of dialysate obtained before the infusion). * p<0.05 when compared to values measured before the dynamic strength training

In situ effects of adrenaline (Adr, 10 μmol/l) alone (diamonds) or in the presence of phentolamine (100 μmol/l; squares) on extracellular glycerol concentration in subcutaneous abdominal adipose tissue during the euglycaemic–hyperinsulinaemic clamp, before and after dynamic strength training in obese subjects with insulin resistance. a, b Absolute values of extracellular glycerol concentration before and after training, respectively. c Changes in extracellular glycerol concentration observed with adrenaline (filled bars) and adrenaline+phentolamine (open bars). The glycerol changes were calculated as for Fig. 4. * p<0.05 when compared to values measured before the dynamic strength training; # p<0.05 when compared to values measured with adrenaline alone
After training, the isoproterenol-induced increase in EGC was higher than that before training (Fig. 4), while the effect of adrenaline was unchanged (Fig. 5). However, the inhibition of the isoproterenol-induced increase in EGC during the insulin clamp was significantly stronger (35%; p=0.05) than before dynamic strength training. In contrast, training did not modify the reduction of the adrenaline-induced increase in EGC (35%) during the insulin clamp (Fig. 5).
In the probe perfused with adrenaline plus phentolamine, EGC was lower after training than before (151±14 vs 252±40 μmol/l; p<0.05). Before training the addition of phentolamine to the perfusate did not modify the increase of EGC in response to adrenaline perfusion before or during the insulin clamp (Fig. 5). After training, phentolamine enhanced the lipid mobilising effect of adrenaline before and during the insulin clamp (Fig. 5a,b).
Modifications of regional adipose tissue blood flow
The local changes in the adipose tissue blood flow (ATBF) were evaluated using the method based on measurement of ethanol washout. The ethanol ratio was calculated as described in Methods and taken as an index of ethanol washout. A higher ethanol ratio corresponds to a lower ethanol washout and to a lower regional ATBF. Fig. 6 depicts the changes in ethanol ratio induced by perfusion with respective agents in the probes before and after training. In the probe perfused with Ringer alone, the ethanol ratio was similar before and after dynamic strength training (81.5±4.6 and 82.2±4.7%, respectively). During the clamp (measured between 165 and 195 min) a decrease in the ethanol ratio before and after dynamic strength training (74.1±7.1 and 73±5.9% respectively) was observed, the decrease being significant only after training (p=0.03). Isoproterenol induced a decrease of the ethanol ratio (indicating vasodilatation) before and during the insulin clamp. Training did not modify this adrenergic-beta-receptor-mediated effect. Adrenaline did not modify the ethanol ratio before or during the insulin clamp and training did not promote any changes in the effect of adrenaline on the ethanol ratio. The addition of phentolamine to adrenaline induced a decrease in the ethanol ratio. Again, training did not modify this additional effect of phentolamine.

In situ effects of various perfusions on the ethanol ratio in subcutaneous abdominal adipose tissue during the euglycaemic–hyperinsulinaemic clamp before (solid bars) and after (open bars) dynamic strength training in obese subjects with insulin resistance. a Control perfusion (Ringer solution alone). b Isoproterenol (Iso, 1 μmol/l) perfusion. c Adrenaline (Adr, 10 μmol/l) alone. d Adrenaline with phentolamine (Adr+ph100 μmol/l) values are means±SEM. *p<0.05 when compared to values measured before basal 1 or basal 2
Discussion
The main finding of this study is that in obese subjects, 3 months of dynamic strength training results in an improvement of insulin resistance. Insulin sensitivity was increased (e.g. antilipolysis) and major changes in the adrenergic regulation of lipolytic pathways were observed in subcutaneous adipose tissue. Plasma levels of lipids and catecholamines were unchanged, whereas leptin levels were reduced.
Lipolysis in human fat cells is controlled by insulin and catecholamines. Adrenaline and noradrenaline promote the activation of both lipolytic adrenergic beta receptors and antilipolytic ADRA2As. The antilipolytic potency of adrenaline, which exhibits a higher affinity for ADRA2A than noradrenaline, has been shown in human subcutaneous adipocytes. Consequently, adrenaline is less efficient in stimulating lipolysis than a selective adrenergic beta receptor agonist like isoproterenol. Moreover, the antilipolytic action of adrenaline is particularly important in subcutaneous adipocytes of obese subjects. We have previously demonstrated that, in obese subjects, the activation of ADRA2A is responsible for reducing the lipolytic effect of catecholamines both at rest [19] or during exercise [3, 19]. Moreover, interactions between the adrenergic and insulin-dependent pathways in the regulation of lipolysis in SCAAT have been demonstrated [6]. In vitro investigations in fat cells have shown that insulin modifies the functional balance between adrenergic beta receptors and ADRA2A in favour of the ADRA2A antilipolytic pathway in human fat cells [5, 7, 20].
Insulin resistance in the patients studied here was assessed using a euglycaemic–hyperinsulinaemic clamp; the patients were found to be in the insulin-resistant range (when compared with populations cited by Skrha et al. [21] or by Katz et al. [18]). In obese patients with insulin resistance, the increase in the lipolytic response induced by the adrenergic beta receptor agonist, isoproterenol, was lower than that previously observed in healthy subjects (1.8-fold vs 3-fold increase, respectively) [6]. Moreover, the effect of adrenaline was not increased under ADRA2A blockade by phentolamine previously found by us in obese women [19]. These results suggest that, in insulin-resistant obese patients, there is a reduction of the adrenergic-beta-receptor-dependent activation of lipolysis in SCAAT. It can also be proposed that, in the state of insulin resistance, the activation of the antilipolytic ADRA2A pathway by adrenaline did not counteract the adrenergic-beta-receptor-dependent lipolytic pathway, since phentolamine did not modulate the lipolytic effect of adrenaline. As expected, insulin infused through a euglycaemic–hyperinsulinaemic clamp reduced the lipolytic action of both isoproterenol and adrenaline. However, in contrast to previous results in healthy subjects [6], phentolamine did not increase adrenaline-induced lipolysis during the insulin clamp. This point confirms that the activation of the ADRA2A pathway by adrenaline cannot counteract the adrenergic-beta-receptor-dependent lipolytic pathway in insulin-resistant patients. Hence, the reduction of the lipolytic effect of adrenaline cannot be explained by the involvement, found in healthy subjects, of the antilipolytic component of adrenaline action related to ADRA2As [3].
The 3-month dynamic-strength training programme modified the lipolytic responses in SCAAT. The first point to note is that dynamic strength training induced a reduction of spontaneous lipolysis in SCAAT, assessed by the reduction of EGC in the control probe (Fig. 2a). During the insulin clamp, the inhibition of spontaneous glycerol release was stronger after training than before (Fig. 2b). We also observed an increase in the lipolytic effect of isoproterenol in the normal insulin state, i.e. before the insulin clamp (Fig. 4). Such an effect on the adrenergic beta receptor pathway in obese subjects has previously been observed during endurance training [22]. This result reveals that dynamic strength training improves the adrenergic beta receptor sensitivity of subcutaneous fat cells in insulin-resistant patients. The second point is that, whereas the insulin clamp reduced isoproterenol-induced lipolysis by 18% before training, the clamp-induced reduction was greater (35%) after training. This suggests that training improved insulin sensitivity in SCAAT, leading to an enhanced counteraction of the isoproterenol effect. However, before the insulin clamp in the post-training investigation, the lipolytic action of adrenaline was unchanged (Fig. 5). After training, blockade of ADRA2As by phentolamine enhanced the lipolytic effect of adrenaline before the beginning of the insulin clamp. This suggests that after training, activation of the antilipolytic ADRA2A pathway by adrenaline can counteract the adrenergic-beta-receptor-dependent lipolytic pathway through increased efficiency of the ADRA2A pathway. This interpretation is also supported by the fact that, during the post-training insulin clamp, the lipolytic effect of adrenaline was enhanced by phentolamine. This effect was not seen in the clamp before the training programme (Fig. 5). We can, therefore, conclude that training enhances the adrenergic beta receptor pathway (assessed by the effect of isoproterenol). Moreover, the restoration of an efficient antilipolytic ADRA2A pathway explains the counteraction of the adrenergic-beta-receptor-mediated response when using adrenaline.
Globally, these modifications are probably linked to the improvement by dynamic strength training of insulin sensitivity in adipose tissue. Apparently the benefits of dynamic strength training are quite similar to those of endurance training in improving insulin resistance status. The dynamic strength training did not promote any changes in ADRB2 or ADRA2A nor in hormone-sensitive lipase or PDE3B gene expression in adipose tissue. These results fit with those of a previous study in obese women, in which a 12-week aerobic training programme did not change the mRNA levels of these genes [19]. The changes observed during dynamic strength training are therefore not related to modifications of gene transcription.
For an accurate interpretation of lipolysis data based on the determination of glycerol concentrations in the dialysate outflow from the microdialysis probe, the regional ATBF must be taken into account. It is known that the EGC in human adipose tissue is influenced by the local blood flow [11]. The method used by us to assess changes in adipose tissue microcirculation was based on the evaluation of ethanol washout as measured using the ethanol ratio. An increase in ethanol ratio reflects a decrease in ATBF. As previously shown, insulin by itself, at the concentration used, does not change ATBF [23]. In this study, the isoproterenol- or adrenaline-induced changes in ATBF were not modified during the insulin clamp. In addition, dynamic strength training had no apparent effect on the ATBF. Finally, the vasodilatation observed when adrenaline was associated with the non-selective ADRA1A/ADRA2A antagonist phentolamine was not modified by the insulin clamp or by the training (Fig. 6).
In general, dynamic strength training appears to improve sensitivity in all major pathways of regulation of lipid mobilisation in subcutaneous adipose tissue in obese subjects, and thus modifies a state of metabolic inflexibility in adipose tissue characteristic for obese and/or diabetic patients. The increase in insulin sensitivity in adipose tissue is supported by a stronger reduction of plasma NEFA and glycerol levels during the insulin clamp after training (Fig. 3b,d). Moerover, it is known that endurance training is followed by a significant decrease in plasma NEFA and an increase in NEFA utilisation as substrate for muscle [24]. In contrast, it has also been reported that dynamic strength training did not improve the use of lipid as substrate at rest [25]. Strength training is supposed to lead mainly to carbohydrate utilisation as substrate for muscle activity, and to less NEFA utilisation, a result which probably explains the higher plasma NEFA levels observed after training in our study.
In summary, the present study demonstrates that dynamic strength training, like endurance training, improves whole-body insulin sensitivity and, simultaneously, sensitivity to the antilipolytic action of insulin in adipose tissue in obese subjects. Moreover, the training induced an increase in sensitivity of the adrenergic-beta-receptor-dependent lipolytic pathway, as well as changes in the functional balance between ADRA2As and adrenergic beta receptors. In insulin-resistant obese subjects, the ADRA2A-dependent pathway does not efficiently counteract the adrenergic beta pathway, and dynamic strength training might improve the fine interplay between these two adrenergic pathways of lipolysis regulation. In conclusion, dynamic strength training appears to improve sensitivity in all major pathways of regulation of lipid mobilisation in subcutaneous adipose tissue in obese subjects. thus modifying a state of metabolic inflexibility in adipose tissue, which is characteristic for obese patients. Consequently, we do not see any reason for preferring one or another type of training in general. Nevertheless, we consider the beneficial metabolic effects of dynamic strength training observed by us to be of clinical importance, as many men in the 40–50 years age group are more willing to engage in dynamic strength training than in classical aerobic training. However, larger comparative studies (and longer training periods) will be necessary before final conclusions can be drawn.
Abbreviations
- ADRA2A:
-
alpha 2A adrenergic receptor
- ADRB2:
-
adrenergic beta-2 receptor
- ATBF:
-
adipose tissue blood flow
- EGC:
-
extracellular glycerol concentration
- PDE3B:
-
phosphodiesterase-3B
- 1-RM:
-
one-repetition maximum strength
- RT-qPCR:
-
reverse transcription-quantitative PCR
- SCAAT:
-
subcutaneous abdominal adipose tissue
References
Arner P (1992) Adrenergic receptor function in fat cells. Am J Clin Nutr 55:228S–236S
Lafontan M, Berlan M (1995) Fat cell α2-adrenoceptors: the regulation of fat cell function and lipolysis. Endocrine Rev 16:716–738
Stich V, De Glisezinski I, Crampes F et al (2000) Activation of α2-adrenergic receptors impairs exercise-induced lipolysis in subcutaneous adipose tissue of obese subjects. Am J Physiol Regul Integr Comp Physiol 279:R499–R504
Cimmino M, Agosto A, Minaire Y, Géloën A (1995) In situ regulation of lipolysis by insulin and norepinephrine: a microdialysis study during euglycemic–hyperinsulinemic clamp. Metabolism 44:1513–1518
Engfedt P, Hellmer J, Wahrenberg H, Arner P (1998) Effect of insulin on adrenoceptor binding and the rate of catecholamine-induced lipolysis in isolated human fat cells. J Biol Chem 263:1553–1560
Stich V, Pelikanova T, Wohl P et al (2003) Activation of α2-adrenergic receptors blunts epinephrine-induced lipolysis in subcutaneous adipose tissue during a hyperinsulinemic–euglycemic clamp in men. Am J Physiol Endocrinol Metab 285:E599–E607
Lonnroth P, Smith U (1986) The antilipolytic effect of insulin in human adipocytes requires activation of the phosphodiesterase. Biochem Biophys Res Commun 141:1157–1161
Moberg E, Enokson S, Hagström-Toft E (1998) Importance of phosphodiesterase 3 for the lipolytic response in adipose tissue during insulin-induced hypoglycemia in normal man. Horm Metab Res 30:684–688
Heyward, VH (1998) Advanced fitness assessment & exercise prescription, 3rd edn. Human Kinetics Pub, Champaign, USA pp 105–144
Jansson PA, Larsson A, Smith U, Lönnroth P (1992) Glycerol production in subcutaneous adipose tissue of lean and obese humans. J Clin Invest 89:1610–1617
Enoksson S, Nordenström J, Bolinder J, Arner P (1995) Influence of local blood flow on glycerol levels in human adipose tissue. Int J Obesity Relat Metab Disord 19:350–354
Felländer G, Linde B, Bolinder J (1996) Evaluation of the microdialysis ethanol technique for monitoring of subcutaneous adipose tissue blood flow in humans. Int J Obesity Relat Metab Disord 20:220–226
Galitzky J, Lafontan M, Nordenström J, Arner P (1993) Role of vascular alpha2-adrenoceptors in regulating lipid mobilization from human adipose tissue. J Clin Invest 91:1997–2003
DeFronzo RA, Tobin JD, Andres R (1979) Glucose clamp technique: a method for quantifying insulin secretion and resistance. Am J Physiol Endocrinol Metab 273:E214–E223
Arvidsson E, Viguerie N, Andersson I, Verdich C, Langin D, Arner P (2004) Effects of different hypocaloric diets on protein secretion from adipose tissue of obese women. Diabetes 53:1966–1971
Bernst E, Gutman I (1974) Determination of ethanol with alcohol dehydrogenase and NAD. In: Bergmeyer HU (ed) Methods of enzymatic analysis, vol. 3. Verlag Chemie, Weinheim, pp 1499–1505
Bradley DC, Kaslow HR (1989) Radiometric assays for glycerol, glucose and glycogen. Anal Biochem 180:11–16
Katz A, Nambi SS, Mather K et al (2000) Quantitative insulin sensitivity check index: a simple, accurate method for assessing insulin sensitivity in humans. J Clin Endocrinol Metab 85:2402–2410
Richterova B, Stich V, Moro C et al (2004) Effect of endurance training on adrenergic control of lipolysis in adipose tissue of obese women. J Clin Endocrinol Metab 89:1325–1331
Moro C, Polak J, Richterova B et al (2005) Differential regulation of ANP- and adrenergic receptor-dependent lipolytic pathways in human adipose tissue. Metabolism 54:122–131
Skrha J, Haas T, Sindelka G et al (2004) Comparison of the insulin action parameters from hyperinsulinemic clamps with homeostasis model assessment and QUICKI indexes in subjects with different endocrine disorders. J Clin Endocrinol Metab 89:135–141
Stich V, de Glisezinski I, Galitzky J et al (1998) Endurance training increases the beta-adrenergic lipolytic response in subcutaneous adipose tissue in obese subjects. Int J Obesity 23:374–381
Stallknecht B, Larsen JJ, Mikines KJ, Simonsen L, Bülow J, Galbo H (2000) Effect of training on insulin sensitivity of glucose uptake and lipolysis in human adipose tissue. Am J Physiol Endocrinol Metab 279:E376–E385
Crampes F, Marion-Latard F, Zakaroff-Girard A et al (2003) Effects of a longitudinal training program on responses to exercise in overweight men. Obesity Res 11:247–256
Achten J, Jeukendrup AE (2004) Optimizing fat oxidation through exercise and diet. Nutrition 20:716–727
Acknowledgements
This study was supported by grants from the Internal Grant Agency of the Ministry of Health of the Czech Republic (IGA 6,836-3 and IGA NR 8,066-3) and by Alfediam/Merck Lipha Santé. Support was also provided by the Direction Générale de la Coopération Internationale et du Développement (Programme d’Action Intégrées Franco-Tchèque). We are grateful to M.-T. Canal and Z. Parizkova for their excellent technical assistance and Dr P. Winterton for revising the English version.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Polak, J., Moro, C., Klimcakova, E. et al. Dynamic strength training improves insulin sensitivity and functional balance between adrenergic alpha 2A and beta pathways in subcutaneous adipose tissue of obese subjects. Diabetologia 48, 2631–2640 (2005). https://doi.org/10.1007/s00125-005-0003-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-005-0003-8