
Abstract
Having to look for multiple targets typically results in switch costs. However, using a gaze-contingent eyetracking paradigm with multiple color-defined targets, we have recently shown that the emergence of switch costs depends on whether observers can choose a target or a target is being imposed upon them. Here, using a similar paradigm, we tested whether these findings generalize to the situation in which targets are specified across different feature dimensions. We instructed participants to simultaneously search for, and then fixate, either of two possible targets presented among distractors. The targets were defined as either two colors, two shapes, or one color and one shape. In one condition, only one of the two targets was available in each display, so that the choice was imposed. In the other condition, both targets would be present in each display, which gave observers free choice over what to search for. Consistent with our earlier findings, switch costs emerged when targets were imposed, whereas no switch costs emerged when target selection was free, irrespective of the dimension in which the targets were defined. The results are consistent with the operation of different modes of control in multiple-target search, with switch costs emerging whenever reactive control is required and being reduced or absent when displays allow for proactive control.
Similar content being viewed by others

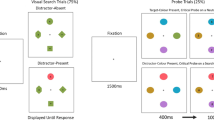
Individuals are often required to find multiple target objects. In some situations, the outcome of such multiple-target searches can be critical—for example, when airport baggage screeners check bags for multiple potential threats. Considering its importance, it is striking that multiple-target search is still poorly understood.
Searching for two or more target objects typically comes with performance costs relative to search for a single target, in terms of both speed and accuracy (Barrett & Zobay, 2014; Biderman, Biderman, Zivony, & Lamy, 2017; Houtkamp & Roelfsema, 2009; Menneer, Barrett, Phillips, Donnelly, & Cave, 2007; Stroud, Menneer, Cave, Donnelly, & Rayner, 2011). Performance suffers in particular when the search target changes from one trial to the next, resulting in what is known as a switch cost (Dombrowe, Donk, & Olivers, 2011; Found & Müller, 1996; Juola, Botella, & Palacios, 2004; Kristjánsson & Campana, 2010; Maljkovic & Nakayama, 1994; Monsell, 2003; Olivers & Humphreys, 2003; Olivers & Meeter, 2006; Wolfe, Butcher, Lee, & Hyle, 2003). These findings suggest that looking for multiple search targets does not allow for the same strength of attentional guidance as looking for a single target does, perhaps even limiting active visual search to a single item at a time (e.g., Olivers, Peters, Houtkamp, & Roelfsema, 2011).Footnote 1
We recently found evidence that the availability of targets in the search display is an important factor in the occurrence of switch costs (Ort, Fahrenfort, & Olivers, 2017). We hypothesized that switch costs emerge when only one of two possible targets is present in the search display. In these cases, observers need to change selection to the other target feature when they happen to be set for the wrong feature on a given trial. In contrast, when both target features are available in each search display, it does not matter which target feature the observer prepares to search for, since either will be available. In such cases no switch costs are expected. To test this explanation, we used a gaze-contingent eyetracking paradigm, in which we instructed participants to simultaneously look for two target colors presented among distractor colors. On every trial, they then selected one of those targets by fixating it. Upon fixation the display disappeared, followed by a new search display. Importantly, in one condition both target colors would be present in the display, so that there would always be a match with whichever target the observer prioritized (if any). In the other condition, only one target color was present, so that the prepared-for and the actual target would often not match. Across three experiments, we found that switch costs occurred when only one target was available per display, whereas switch costs were either strongly reduced or absent when both target colors were available.
The influence of target availability on switch costs in visual search can be explained from a cognitive control framework that distinguishes between reactive and proactive control (Braver, 2012). Whereas proactive control refers to sustained activation of task-relevant information in anticipation of a task, reactive control occurs when task goals need to be adjusted when no events match the current task goals. Importantly, reactive control always leads to increased processing times because, by definition, it is invoked after a conflict. In the case of search for multiple targets, this would mean that if only one target is present in the display and it does not match the anticipated target, reactive control will be required in order to swap to a different search template, and switch costs will emerge. In contrast, if all potential targets are available for selection, proactive control mechanisms allow for preparation for any of the possible targets without running into selection conflicts. Depending on the time that such preparation takes, switch costs should be reduced or absent, as compared to the case of reactive control. As we argued earlier (Ort et al., 2017), the idea of a role for proactive control when observers can prepare for any one target, because all targets are always available anyway, may also explain the earlier findings of a lack of switch costs during multiple-target search (Beck & Hollingworth, 2017; Beck, Hollingworth, & Luck, 2012). We note as well here that preparing the search does not need to be a deliberate, willful act. Although target expectations and selections may be explicit and voluntary, they need not be (Awh, Belopolsky, & Theeuwes, 2012; Kristjánsson & Campana, 2010), and it remains an open question in which precise way observers would employ control over selection.
Here we extend the reactive-versus-proactive control framework to visual search for objects defined across different feature dimensions. In daily life, objects are typically composed of features in multiple dimensions, and each of these features could potentially be used to guide attention. We asked whether the effects of target availability on switch costs that we had previously observed for targets defined within a feature dimension (color) would also hold when targets were defined in different dimensions (color and shape). On the one hand, it is known that cross-dimension search is associated with costs, with some studies showing them to be higher than for within-dimension search (e.g., Found & Müller, 1996; Menneer et al., 2007; Müller, Heller, & Ziegler, 1995; Olivers & Meeter, 2008; Williams, 1967). However, these studies used single-target displays and did not compare performance to conditions in which multiple targets were available in the search display. It may be that when targets vary across dimensions, switch costs would also emerge for multiple-target displays.
On the other hand, there is reason to believe that switch costs may actually be reduced for cross-dimension search. Influential theories of attention have maintained that different object dimensions are processed largely separately and in parallel, in so-called feature maps (e.g., Treisman & Gelade, 1980; Wolfe, 1994). It is possible that although preparing for multiple target features within a feature map incurs costs, preparing for multiple target features across different feature maps may preclude such costs, by tapping into separate resources. It has been shown that searching for a target that is defined by two features from the same dimension (e.g., a conjunction of two colors) is more difficult than searching for a target defined by two features from different dimensions (e.g., a conjunction of orientation and color; Allport, 1971; Wolfe et al., 1990). However, these studies always combined features within a single target, and thus did not investigate multiple-target search and the accompanying switch costs.
To investigate the role of target availability on cross-dimension search, we adapted the gaze-contingent eyetracking paradigm we had used in Ort et al. (2017), as is illustrated in Fig. 1. We instructed observers to look for two features and to make an eye movement toward one of them in subsequent search displays. Participants either had to search for two colors, two shapes, or one color and one shape. Importantly, the feature dimensions were always kept fully separate, such that the target color and target shape were never presented in the same stimulus. In separate blocks, either one or both of the targets would be present in the search display. If target availability has the same effect on cross-dimension as on within-dimension search, we would expect to find switch costs when only one target was present in the search display, and no switch costs when both targets were present. If, however, multiple-target search is more efficient when the targets are defined in different feature dimensions, we would expect to observe reduced switch costs for the one-target-available condition, relative to within-dimension search. Finally, switching across feature dimensions could also be harder than within feature dimensions, potentially leading to switch costs even when two targets are in the display.

Illustration of the study design. (a) Example sequence for a cross-dimension search block. Each block began with a cue indicating the two target features for the subsequent sequence of search displays. Depending on the condition, each of the 40 search displays contained either one target feature or both target features. Participants were required to fixate one of the targets (indicated here by an arrow, which was not present in the display); this triggered the next display, which appeared in an imaginary circle surrounding the location of the previously fixated target. (b) Example search displays for each experimental condition
In addition, with the present design it is possible to measure the effects of top-down processes while keeping the bottom-up signal in the search display constant across conditions. Note that dimension switch costs have typically been studied with singleton search displays (e.g., Found & Müller, 1996; Krummenacher, Müller, & Heller, 2001; Müller et al., 1995). In those studies, a cross-dimension switch always coincided with a substantial change in the bottom-up characteristics of the search display, affecting the contrast between the target and distractors. In such cases, any cross-dimension switch costs cannot unambiguously be attributed to top-down factors, since part of the effect may be driven by low-level sensory changes (the same also goes for the efficiency of cross-dimension search; see Quinlan & Humphreys, 1987). In the present design, the search display composition was such that the target was never associated with a unique bottom-up signal, thus providing a more direct measure of the cognitive processes involved in target changes both across and within feature dimensions.
Method
Participants
A sample of 20 participants (age: 18–34 years, M = 24.4; 11 females, nine males) was recruited from the Vrije Universiteit Amsterdam and participated for course credit or money (sample size based on Ort et al., 2017, Exps. 2 and 3). All participants gave written consent according to the Declaration of Helsinki. All of them reported normal or corrected-to-normal visual acuity and color vision and were naive to the purpose of the experiment. The study was approved by the ethics board of the Faculty of Behavioural and Movement Sciences at Vrije Universiteit Amsterdam.
Apparatus
The experiment was designed and presented using the OpenSesame software package (version 3.0.7; Mathôt, Schreij, & Theeuwes, 2012) in combination with PyGaze (Version 0.6), an eyetracking toolbox (Dalmaijer, Mathôt, & Van der Stigchel, 2013). Stimuli were presented on a 22-in. Samsung Syncmaster 2233RZ screen with a resolution of 1,680 × 1,050 pixels and a refresh rate of 60 Hz, at a viewing distance of 75 cm. Eye movements were recorded with the EyeLink 1000 eyetracking system (SR Research, Mississauga, Ontario, Canada) at a sampling rate of 1000 Hz. The experiment took place in a dimly lit, sound-attenuated room. The experimenter received real-time feedback on system accuracy on a second monitor located in an adjacent room. After every block, eyetracker accuracy was assessed and improved as needed, by applying a 9-point calibration and validation procedure.
Stimuli and procedure
The stimulus set consisted of all possible combinations of five colors and five shapes. The shapes were a square, a triangle, a five-pointed star, a plus, and a diamond (i.e., a square rotated 45 deg) and spanned on average 1.0 deg of visual angle (SD = 0.15 deg). These canonical shapes were chosen because they were easy to remember. The colors were blue (RGB values: 0, 130, 150), red (240, 0, 0), green (70, 135, 0), brown (175, 100, 75), and purple (180, 80, 170). All these colors were isoluminant (M = 21 cd/m2). The background color was a uniform gray (197, 197, 197, luminance = 59 cd/m2).
A block began with a white fixation cross for 500 ms, followed by the cue display for 2,500 ms and another fixation cross for 500 ms (see Fig. 1a). In the cue display, the two task-relevant target features were presented 1.9 deg of visual angle to the left and right of fixation. To disambiguate task relevance, the target colors were presented on a disk and target shapes were colored black, because neither disks nor black items ever appeared in the search displays as stimuli. The cue display was followed by a sequence of 40 search displays. Each search display consisted of five color–shape conjunctions. The five objects were placed on an imaginary circle around fixation with a radius randomly drawn from values between 3.8 and 4.7 deg of visual angle around the starting point, and an angular distance between each of the elements of at least 45 deg. Any single target object (whether one or both targets were available) never contained both target features, always one of the two.
Participants were instructed to make an eye movement toward a stimulus that contained one of the target features. After target fixation, the search display disappeared from the screen, and the fixated target was replaced by a black ring to provide participants with a fixation point while waiting for the next search display, which appeared after 600 to 800 ms (uniformly jittered). Because the coordinates of the previously fixated target served as the starting point for the next display, the search traveled across the screen throughout a block. Considering this dynamic character of the paradigm, it could happen that when the search approached an edge or corner of the computer screen, there was not sufficient space on the full imaginary circle to present all stimuli spaced at least 45 deg apart. In these cases, objects were placed in closer proximity to each other, away from the edge or corner, with a minimal separation of 31 angular degrees, but always keeping the same distance from fixation for all objects. Fixations had to land in an area with a radius of 0.74 deg of visual angle, centered on a target, to trigger the next search display. If participants fixated one of the distractors, they received auditory feedback and were required to make a corrective eye movement toward a target. The search was aborted if no target was fixated within 3,000 ms, and a new search display appeared.
There were three main factors in this experiment: feature dimensionality (the dimension in which the targets were defined), target availability (whether only one or both targets were present in the search display), and trial type (whether the target selection switched or repeated from one trial to the next). Feature dimensionality was manipulated at the block level: Per block, the two target features could both be colors (within-color condition), both be shapes (within-shape condition), or be one color and one shape (cross-dimension condition). To reduce interference across blocks, all within-dimension blocks were presented randomly intermixed in one half of the experiment, and all cross-dimension blocks in the other half. The order in which these halves were presented was counterbalanced across participants. Other than the relevant feature dimensions imposed by the target instructions, the specific target features were randomly selected for every block.
The target availability factor was implemented by manipulating whether only one or both of the target features were presented in the display. Target availability was also varied at the block level (previous findings having suggested that manipulating target availability at the block or the trial level does not qualitatively change the results; Ort et al., 2017). In the both-targets condition, both cued targets appeared in the search display. In the one-target condition, only one of the cued targets was present. The trial type factor (target repeat vs. target switch) was determined by the observer (both-targets blocks) or by a random sampling procedure (one-target blocks). This sampling procedure randomly selected (with replacement) a sequence of repeat and switch trials from a pool of sequences that were recorded during both-targets blocks. The motivation behind that procedure was to match one-target blocks and both-targets blocks in terms of switch rate and the number of consecutive repeat trials. Only the sequence of switch and repeat trials was replayed, not the features or positions of the stimuli. Because we did not yet have any sequences to present in one-target blocks at the outset of the experiment, we initialized a pool with four prespecified random sequences of switch and repeat trials (one each for eight, 12, 16, and 20 switches per block). Completed both-targets blocks were added to this pool throughout the experiment. The sequence of repeat and switch trials for a given one-target block was then randomly chosen from that pool. In this way the repeat–switch sequences in one-target blocks had the same distribution as the sequences in both-targets blocks, while also remaining unpredictable. A paired-samples t test confirmed that the switch rates did not differ significantly between target availability conditions [both targets available: 12.5 switches; one target available: 12.6 switches per block; t(19) = 0.30, p = .77, Cohen’s d = 0.06], which was confirmed by a Bayesian t test suggesting more evidence for the absence of a difference than for its presence (unless otherwise noted, all Bayes factor analyses were performed with the alternative hypothesis in the numerator; hence, the result here of BF = 0.24 indicates greater evidence for the null).
Both-targets-available and one-target-available blocks would differ not only in terms of target availability, but also in the mere number of targets in the display, which would make the one-target-available condition more difficult than the both-targets-available condition. Therefore, we included trials in the one-target-available condition in which there were two target objects, but both carried the same target feature, so that still only one target feature was present in the search display (i.e., target duplicate; e.g., during search for a red or a blue target in the one-target-available condition, there could be trials with two red targets or two blue targets, but never with a red and a blue target). In addition, we included trials in which two distractors shared a feature in the task-relevant dimension (i.e., distractor duplicate; e.g., during search for a red or a blue target, there could be trials with two green distractors), so that participants could not identify the target by merely detecting a feature duplication. Likewise, both-targets blocks also contained target duplicate trials (in which two out of three targets shared a target feature; e.g., during search for a red or blue target, we had trials with one red and two blue target objects), as well as distractor duplicate trials (e.g., trials with one red target, one blue target, and two green distractors). As a result, one target feature was duplicated on half of the trials of each target availability condition, and one distractor feature was duplicated on the other half of the trials. In this way, neither the number of targets nor the number of unique features in the display was predictive of target availability. Nevertheless, we also statistically examined whether this factor (duplicate type) affected our main findings, and it did not (see the Results). In the within-color and within-shape conditions, all stimuli were unique in terms of the task-irrelevant dimensions (e.g., if the targets were defined by color, all stimuli had a unique shape). Hence, any target duplicates and distractor duplicates were defined only in the task-relevant dimension. Because both dimensions were task-relevant in the cross-dimension condition, the dimension that contained a duplicate was chosen randomly across trials.
Participants completed 24 cross-dimension blocks, 12 within-color blocks, and 12 within-shape blocks, for 48 experimental blocks in total. In each feature dimensionality condition, both-targets-available and one-target-available blocks appeared equally often, but randomly intermixed. Before the experimental blocks started, participants practiced all of the six unique block types once.
Data preprocessing
For all experimental conditions, we compared the saccade latencies of eye movements (dwell times before a saccade was executed) for repeat trials (current target category the same as the previous one) with those for switch trials (current target category different from the previous one). We took the first saccade after search-display onset with a minimum amplitude threshold of 1 deg of visual angle around the initial fixation, provided that the saccade was directed toward the selected target (i.e., it was directed more to the target than to any of the other items in the display, and its direction was no more than 30 angular degrees away from the direction of the target). This resulted in an average of 33.6% of all search displays being removed. Furthermore, a saccade latency filter was applied, in which saccades quicker than 100 ms and slower than three standard deviations above the block mean (based on all trials) for that participant were excluded (4.8% of all search displays). If no target was being fixated, as could have happened when the eyetracker calibration deteriorated, both the current and the next search display were excluded, because neither could be labeled as a switch or repeat (6.4% of all search displays). For the same reason, we excluded the first search display of each block (2.5% of all search displays). In total, 33.7% of all search displays were removed during preprocessing (note that a single trial could meet multiple exclusion criteria).Footnote 2 Analyses were carried out with the afex R package (Singmann, Bolker, & Westfall, 2015).
Results
Cross-dimension search versus within-dimension search
Because we were mostly interested in the comparison of the overall switch costs for cross-dimension search with those for within-dimension search, for the main analysis we collapsed the within-shape and within-color conditions.
Figure 2a shows the mean saccade latencies on switch trials and repeat trials as a function of target availability, separately for within-dimension and cross-dimension blocks. To investigate this data pattern, we ran a three-way repeated measures analysis of variance (ANOVA) on the mean saccade latencies, with target availability (one vs. both), feature dimensionality (within- vs. cross-dimension), and trial type (repeat vs. switch) as factors, which revealed significant main effects of target availability, F(1, 19) = 69.0, p < .001, ηp2 = .78, and trial type, F(1, 19) = 17.2, p < .001, ηp2 = .47. All two-way interactions were also significant [Target Availability × Feature Dimensionality: F(1, 19) = 16.7, p < .001, ηp2 = .47; Target Availability × Trial Type: F(1, 19) = 23.6, p < .001, ηp2 = .55; Feature Dimensionality × Trial Type: F(1, 19) = 9.9, p = .005, ηp2 = .34]. Neither the main effect of feature dimensionality, F(1, 19) = 3.3, p = .09, ηp2 = .15, nor the three-way interaction, F(1, 19) = 0.28, p = .61, ηp2 = .01, reached significance. Overall, saccade latencies were longer for target switches than for target repetitions. This difference originated primarily in the one-target conditions and was strongly reduced in the both-targets conditions, as suggested by the two-way interaction between trial type and target availability. Furthermore, saccade latencies were generally greater for one-target than for both-targets blocks.

Main results. (a) The bar plots represent the mean saccade latencies on switch trials and repeat trials for each level of target availability (one target vs. both targets) and feature dimensionality (cross-dimension vs. within-dimension). The gray lines represent the mean saccade latencies for each observer individually. (b) The within-dimension search blocks were split up into blocks in which the targets were color-defined (within-color search) or shape-defined (within-shape search) and here are plotted separately. Error bars represent the upper limit of the within-subjects 95% confidence intervals (Morey, 2008)
To check whether any of these effects were mitigated by block order or by the fact that sometimes the target color and sometimes the distractor color duplicated, we ran two four-way ANOVAs, one with block order (cross-dimension search first vs. within-dimension search first), the other with duplicate type (target duplicate vs. distractor duplicate), as additional factor. However, although there were a main effect of duplicate type [F(1, 19) = 11.3, p = .003, ηp2 = .37] and a two-way interaction between duplicate type and target availability [F(1, 19) = 8.0, p = .01, ηp2 = .3]—with saccade latencies on target duplicate trials being smaller than those on distractor duplicate trials, particularly on one-target-available blocks—neither block order nor duplicate type interacted with trial type or feature dimensionality. This is consistent with our previous study, in which we also found that duplicate type did not modulate the interaction between trial type and target availability (Ort et al., 2017).
A Bayes factor (BF) analysis yielded similar results: The model supported most by the data was the one that included all three main effects, the two-way interaction between feature dimensionality and target availability, and the two-way interaction between trial type and target availability (BF = 1.96 × 1019). The next best models contained all main effects and all two-way interaction effects (BF = 1.32 × 1019) or all main effects with only the two-way interaction between target availability and trial type (BF = 1.64 × 1019).
Further support comes from participants’ fixation accuracy, shown in Table 1. The pattern points toward the same conclusion drawn on the basis of saccade latencies, which precluded an interpretation in terms of speed–accuracy trade-offs. The same ANOVA on accuracy scores yielded significant main effects of target availability, F(1, 19) = 18.0, p < .001, ηp2 = .49, and trial type, F(1, 19) = 23.6, p < .001, ηp2 = .55, as well as a significant interaction between them, F(1, 19) = 28.7, p < .001, ηp2 = .60. All other effects were nonsignificant (p values > .4). Overall, performance was worse in the one-target than in the both-targets condition, but again, especially after a switch.
The nonsignificant three-way interaction in the three-way ANOVA on saccade latencies suggests that the interaction between trial type and target availability was not different for cross-dimension and within-dimension search. To further test this conclusion and compare to the results from our previous study (Ort et al., 2017), we ran two two-way ANOVAs with the factors target availability and trial type on saccade latencies, separately for the cross-dimension and within-dimension conditions. Both main effects as well as the interaction were significant in both ANOVAs [within-dimension: main effect of target availability, F(1, 19) = 45.7, p < .001, ηp2 = .71; main effect of trial type, F(1, 19) = 17.5, p < .001, ηp2 = .48; Target Availability × Trial Type interaction, F(1, 19) = 27.0, p < .001, ηp2 = .59; cross-dimension: main effect of target availability, F(1, 19) = 73.4, p < .001, ηp2 = .79; main effect of trial type, F(1, 19) = 16.1, p < .001, ηp2 = .46; Target Availability × Trial Type interaction, F(1, 19) = 15.2, p = .001, ηp2 = .44]. These patterns were confirmed by two Bayesian ANOVAs with the same factors, in which the data provided most evidence for those models that included both main effects and the two-way interaction, both for the within-dimension condition (BF = 1.9 × 109, 26 times as likely as the next best model) and for the cross-dimension condition (BF = 3.9 × 109, two times as likely as the next best model).
These results show that switch costs were larger in the one-target-available condition than in the both-targets-available condition for both cross-dimension (one-target: M = 91 ms; both-targets: M = 30 ms) and within-dimension (one-target: M = 64 ms; both-targets: M = 8 ms) search, although the cross-dimension search switch costs never completely disappeared. Classical t tests indeed indicated significant switch costs in all one-target-available conditions [cross-dimension: t(19) = 4.20, p < .001; within-dimension: t(19) = 4.98, p < .001], and also for the cross-dimension, both-targets condition, t(19) = 2.88, p < .001, but not for the within-dimension, both-targets condition, t(19) = 1.29, p = .21. Bayesian t tests further confirmed the pattern, with very strong evidence for the presence of switch costs for both types of feature dimensionality when only one target was available (cross-dimension: BF = 68; within-dimension: BF = 326), whereas when both targets were available, there was still moderate evidence for the presence of switch costs in the cross-dimension search (BFSwitchCosts = 5.3), whereas there was anecdotal evidence for the absence of switch costs in the within-dimension search (BFNoSwitchCosts = 2.0). Furthermore, classical t tests indicated significantly larger switch costs for cross-dimension than for within-dimension search (cross-dimension: M = 91 ms; within-dimension: M = 64 ms), t(19) = 2.43, p = .03, whereas the Bayesian t test indicated that the evidence for this difference was only anecdotal (BF = 2.1), casting doubt as to whether this difference is really meaningful.
Finally, we also examined the switch rates for within-dimension and cross-dimension blocks when both targets were available. In line with previous findings on voluntary task switching (e.g., Arrington & Logan, 2004, 2005), observers showed a repetition bias. In within-dimension blocks, target switches occurred on 35.0% of all trials; in cross-dimension blocks, on 28.3% of all trials; and this difference was significant, t(19) = 2.6, p = .02, Cohen’s d = 0.58, BF = 3.3.
Comparison of search for color-defined versus shape-defined targets
It is conceivable that the observed effects in the within-dimension conditions could have been caused by either the within-color or the within-shape search blocks alone. For example, because shape has been shown to guide attention less well than color (Williams, 1967; Wolfe & Horowitz, 2004), saccades to shape-defined targets could have been slow regardless of whether the target repeated or switched or how many targets were available for selection, but this might also have interacted with switch costs. Therefore, we next focused on the within-dimension conditions, but instead of collapsing them, we directly compared within-dimension search for two colors to within-dimension search for two shapes. Figure 2b shows the mean saccade latencies for this analysis.
We ran a three-way repeated measures ANOVA on mean saccade latencies, with target availability (one vs. both), feature dimensionality (here, within-color vs. within-shape), and trial type (repeat vs. switch) as factors. This yielded significant main effects of target availability, F(1, 19) = 46.8, p < .001, ηp2 = .71; feature dimensionality, F(1, 19) = 40.7, p < .001, ηp2 = .53; and trial type, F(1, 19) = 21.3, p < .001, ηp2 = .53, as well as significant two-way interactions between target availability and feature dimensionality, F(1, 19) = 15.4, p < .001, ηp2 = .45, and between target availability and trial type, F(1, 19) = 26.6, p < .001, ηp2 = .58. Neither the two-way interaction between feature dimensionality and trial type nor the three-way interaction reached significance (p values > .10). These results were supported by a Bayes factor analysis, which indicated that the data provided the most evidence for the model that included all main effects as well as the two-way interactions between target availability and feature dimensionality and between target availability and trial type (BF = 3.3 × 1023); this model was 2.4 times more likely than the next best model (which included all main effects and all two-way interactions).
Saccade latencies were considerably longer on within-shape than on within-color blocks, and they were also longer on one-target than on both-targets blocks. Here we also ran classical and Bayesian t tests, to inspect in which conditions switch costs were present. For within-color blocks, classical t tests indicated significant switch costs in the one-target condition (M = 58 ms), t(19) = 4.41, p < .001, but not in the both-targets condition (M ≈ 0 ms), t(19) = 0.001, p < 1. Bayesian t tests confirmed this result by indicating strong evidence for the presence of switch costs when only one target was available (BFSwitchCosts = 104), and moderate evidence for the absence of switch costs when both targets were present (BFNoSwitchCosts = 4.3). For within-shape blocks, we observed significant switch costs for both one-target blocks (M = 88 ms), t(19) = 3.93, p < .001, and both-targets blocks (M = 17 ms), t(19) = 2.32, p = .03. Bayesian t tests indicated strong evidence for the presence of switch costs in the one-target condition (BF = 40), but only anecdotal evidence in the both-targets condition (BF = 2.0). The pattern of switch costs in the two within-dimension conditions confirmed that when one target was available, the switch costs observed in the collapsed within-dimension condition (Fig. 2a) originated in both feature dimensions, whereas the switch costs that were observed in the both-targets condition primarily originated in the within-shape blocks. Overall, however, selecting color targets appeared to be somewhat less demanding than selecting shape targets, as indicated by shorter saccade latencies. This was also confirmed by participants preferentially fixating color-defined targets (67.9% of all fixations) when they could choose which target to look at (both-targets-available condition).
We further tested whether color targets and shape targets contributed to the same extent to the presence of switch costs across all levels of feature dimensionality and target availability. To that end, we ran two separate two-way ANOVAs for within-color and within-shape blocks, with the factors trial type and target availability on saccade latencies, and also a three-way repeated measures ANOVA with the factors target availability (one vs. both), trial type (repeat vs. switch), and selected dimension (color vs. shape) on the mean saccade latencies for cross-dimension blocks. Overall, in all conditions the patterns did not differ between color-defined and shape-defined targets. The full ANOVAs and further statistical tests can be found in the supplemental material.
Influence of task-irrelevant features on search performance
It has been shown that features can be stored in visual working memory as part of an integrated object (Luck & Vogel, 1997; Wheeler & Treisman, 2002) and that attentional selection can operate on the object level (O’Craven, Downing, & Kanwisher, 1999; Roelfsema, Lamme, & Spekreijse, 1998). On the one hand, integration of features into an object might have the advantage that maintaining the features that comprise an object is less taxing than having to maintain two separate representations. On the other hand, teasing apart the features whenever they are needed for attentional guidance might cause interference and substantial costs. Considering that we did not include conjunctions of target color and target shape in our search displays, there appeared to be no reason for observers to integrate the target features into a single representation. Nevertheless, we wanted to test what influence the currently task-irrelevant feature had on attentional selection. Therefore, we tested whether participants could efficiently ignore the task-irrelevant feature during search.
We ran an exploratory analysis to tease apart the effects that switches of the task-relevant and task-irrelevant features had on saccade latencies. For example, if observers were looking for two colors, irrespective of a target switch or target repeat, the shape of the target object could also randomly change or repeat. To that end, we split target switch and target repeat trials on the basis of whether the currently task-irrelevant feature repeated or changed, separately for the combined within-dimension conditions and the cross-dimension condition, as well as for the one-target and both-targets block conditions. Since task-irrelevant switch trials could not be separated from task-relevant switch trials in the cross-dimension condition (where a switch in the task-relevant feature always coincided with a task-irrelevant feature switch), we analyzed only the task-relevant repeat trials across both levels of feature dimensionality. Figure 3 shows the data for this analysis.

Influence of the task-irrelevant feature on saccade latencies, shown separately for both-targets and one-target blocks as well as for cross-dimension and within-dimension search. Error bars represent the upper limit of the within-subjects 95% confidence intervals (Morey, 2008)
We ran a three-way repeated measures ANOVA with the factors target availability (one vs. both), feature dimensionality (within-dimension vs. cross-dimension), and task-irrelevant feature (repeat vs. switch) on mean saccade latency. This ANOVA yielded significant main effects of target availability, F(1, 19) = 37.4, p < .001, ηp2 = .66, and task-irrelevant feature, F(1, 19) = 37.5, p < .001, ηp2 = .66, as well as a significant two-way interaction between target availability and feature dimensionality, F(1, 19) = 11.2, p = .003, ηp2 = .37. None of the other effects reached significance (p values > .37). This pattern was largely confirmed by a Bayesian ANOVA with the same factors. The data provided most evidence for the model that included all main effects and the two-way interaction between target availability and feature dimensionality (BF = 6.3 × 1015). This model was four times as likely as the next best models.
Whereas general saccade latencies on cross-dimension blocks were longer than on within-dimension blocks for one-target blocks, there was no difference for both-targets blocks. Task-irrelevant switch trials were consistently slower than task-irrelevant repeat trials across all levels of feature dimensionality and target availability (M = 14 ms). This suggests that the irrelevant feature could not be ignored entirely, and that feature integration happened to some extent. Interestingly, this was also the case for the within-dimension conditions, in which the task-irrelevant feature dimension could have been ignored for an entire block.
Discussion
Having to look for multiple target objects typically comes with switch costs when the target changes between presentations. We have shown previously that the occurrence of such switch costs depends on target availability, with switch costs occurring when one of the two potential targets is present in the search display, but not when both targets are available for selection. The present results replicate this pattern for color-defined targets and show that it extends to shape-defined targets as well as to cross-dimension search when the targets are defined in terms of either color or shape. We found reliable switch costs when only one target was available, whereas switch costs were strongly reduced when both targets were available, regardless of whether targets were defined within or across feature dimensions. These findings provide further support for a role of different modes of cognitive control during multiple-target search. When both targets are available, observers can prioritize either target over the other prior to search display onset. Whichever target is being prioritized, there will be a match. In contrast, when only one target is available, reactive control mechanisms are necessary whenever the present target object does not match the anticipated target.
Unlike in our previous study (Ort et al., 2017), small but reliable switch costs also emerged when both targets were available for selection. This finding is consistent with the idea that proactive preparation of a target switch is in itself time-consuming, but that the process can start earlier than in the case of reactive control. Interestingly, whereas we found slight switch costs even when both targets were available, Beck and Hollingworth (2017) found no switch costs in one of their conditions even when only one target was available—despite their experiment being quite similar to ours. In this condition, observers were asked to fixate a target drawn from a set of two alternatives, in which on each trial a single target was presented together with a distractor. Upon fixation of the first target, a new target would appear, together with another distractor. The target could either be the same or different, but this did not affect the saccade latency. However, there were several differences between their task and ours, such as the length of the eye movement sequences, and the timing as well as the task demands after the target had been found. We are currently investigating some of the factors that might have contributed to these contradictory findings.
We found that switch costs tended to be somewhat larger for shape-defined targets than for color-defined targets, although this difference was not significant. This might reflect the fact that visual search for shape-defined targets is typically more difficult than search for color-defined targets (Williams, 1967; Wolfe & Horowitz, 2004), and the switch costs might scale accordingly. Indeed, our data revealed longer saccade latencies and lower accuracy scores whenever observers had to fixate a shape-defined rather than a color-defined target, and when our participants had the choice, they showed a preference for color-defined targets. This preference for color likely also led to a lower switch rate for cross-dimension than for within-dimension blocks whenever observers had the choice, since they were less inclined to switch to the more difficult search for the shape-defined target. This is in accordance with guidance toward shape being reduced here, as compared to guidance toward color.
The present results provide little indication that visual search for targets drawn from different dimensions is qualitatively different from search for targets drawn from the same feature dimension. Previous research has produced mixed findings on the efficiency of cross-dimension search relative to within-dimension search. On the one hand, a number of studies have not looked at switch costs, but instead at the overall efficiency of search as a function of the number of targets to be looked for. Wolfe and colleagues (Wolfe, Cave, & Frenzel, 1989; Wolfe et al., 1990) found more efficient attentional guidance toward two targets from different dimensions than toward two targets from the same dimension, suggesting that two dimensions, but not two features, may provide for attentional guidance in parallel. However, apart from previous research not investigating switch costs, the difference from our experiments is that the search targets used by Wolfe and colleagues were always conjunctions of both target features, whereas distractors carried either one of the target features. Observers thus had to integrate both features, and the designs did not allow for the measurement of attentional guidance toward each of the target features in isolation. In contrast, our cross-dimension search setup encouraged participants to separately look for either of the target features, as the target was never a conjunction of target shape and target color, and the cue displays always presented the two target features separated in space. Under these conditions, attentional guidance toward two target features appears to be limited, even when the features originate in different dimensions. Nevertheless, this leaves open the possibility that if observers could combine target features into a single object representation (e.g., looking for “red” and “square” by looking for “a red square”), search could become more efficient. Future studies should further investigate whether having two separate feature templates (as we studied here) is any different from having an integrated object template consisting of multiple features—in particular, in the context of the distinction between proactive and reactive control.
Another study that suggested that search can occur in parallel across feature dimensions was reported by Quinlan and Humphreys (1987). They found that searching for two distinct target items that were specified in different feature dimensions (their both condition) was as efficient as searching for the less guiding target alone (their feature conditions) in terms of search slopes. However, even though the search slopes were comparable, search for two targets was slower overall than search for a single feature. They interpreted this as an additional, non-search-related process, such as a switch from one target to the other that needs to be invoked in order to make the decision about target presence. However, here, too, switch costs were not directly investigated. Moreover, the tasks differed, in that Quinlan and Humphreys instructed observers to always find both targets in each search display, whereas the observers in the present study only made a single selection. Finally, display differences also preclude a straightforward comparison with the present study, since the difference between two-target and single-target conditions in Quinlan and Humphreys’s study also involved changes in the bottom-up input in terms of target–distractor similarity, and thus target salience.
On the other hand, many studies that have looked at switch costs have shown that switching between feature dimensions comes at a larger cost than switching between features within the same dimension (e.g., Found & Müller, 1996; Krummenacher et al., 2001; Müller et al., 1995; Olivers & Meeter, 2008). In these studies, the targets were always singletons presented within a homogeneous array of distractors. The enhancement of switch costs may therefore partly have been due to the fact that changes of the target dimension always coincided with changes in the bottom-up signal—specifically, the stimulus contrast between target and distractor—rather than to cognitive factors, such as presumed difference in the task sets. Furthermore, those previous studies measured manual response times, and additional costs may therefore have arisen from postselection stages. Our design employed very heterogeneous displays, with changes in the bottom-up signal being independent of the target set, and we took the first eye movement as a measure of selection. This arguably provides a more direct measure of the top-down processes involved in target changes across and within feature dimensions, and our study suggests that when sensory and response factors are controlled for, the switch costs associated with dimension changes are similar to those for feature changes.
Single target capacity or limited resource?
The observation of switch costs when only one target was available in the display is consistent with the idea that observers do not prepare equally for both targets at the same time. Previous research has attributed dual-target costs to either a reduction in the specificity of the target representations or a limit on simultaneously active target representations. According to the former account, trying to look for multiple targets at the same time decreases the amount of resources that is available for maintaining the target representations, so that overall guidance by both target representations is reduced, relative to one-target search (Barrett & Zobay, 2014; Roper & Vecera, 2012). However, this account is not supported by the present data. If the dual-target cost simply originated in an overall reduction of target representation specificity, then switch trials as well as repeat trials should have been affected to similar extents. Likewise, the difference between the target availability conditions is hard to reconcile with an account that is based on less precise target representations, because guidance to all targets in a search display should be reduced, irrespective of how many targets are available for selection. One might argue that observers were able to simultaneously prepare for both target colors only during both-targets blocks, whereas they chose to prepare for just one target during one-target blocks. However, this goes against the findings in Experiment 3 of Ort et al. (2017), in which we manipulated target availability on a trial-by-trial level, so that observers could not predict whether a particular trial would include two targets or only one. Critically, the same data pattern emerged there, as well: switch costs for one-target trials, and no switch costs for both-targets trials.
We believe that the present data are more consistent with models of working memory that propose that only a single item (the target template) in memory can drive visual attention, whereas other memorized items stay dormant in the background, where they are shielded from perceptual input (Huang & Pashler, 2007; Oberauer, 2002; Olivers et al., 2011). If more than one target could be fully prepared for, and thus could guide attention at a given time, we should not have found switch costs in the both-targets-available conditions. Irrespective of whether one or both targets are available for selection in the search display, two equally active search templates should efficiently guide attention toward either target. Furthermore, the present data indicate that this limitation is not restricted to features within a feature dimension, but also holds for features that are specified in independent feature dimensions. However, the assumption that only a single target template is active at a time may be too strict. The same data can also be captured by a hybrid account that assumes that resources are distributed across multiple target representations, but in an unequal fashion, without the need of imposing a limitation of a single slot. In this sense, while one is looking for multiple targets, resources are being reallocated between target representations according to the task requirements and selection history, but at any moment in time, one representation receives the majority of resources, so that attentional guidance is stronger by this item than by the others. Although in that case there would be no fundamental difference in the representational state, the implication would then still be that observers are unable to spread resources equally among different target representations, even if this would be more optimal under a task’s circumstances.
Relationship with long-term memory
So far, we have discussed the results in relation to working memory. However, there is the distinct possibility that we were measuring not just the effects of working memory, but also of long-term memory. It has been shown that working memory representations can already be transferred to long-term memory after five to seven target repetitions (Carlisle, Arita, Pardo, & Woodman, 2011). Given the sometimes relatively long streaks of target repetitions in our experiment, the same transfer might have occurred here. However, we believe this was not the case. First, in a post-hoc confirmatory analysis, we examined saccade latencies as a function of repetition streak length and could not find any indication of decreasing saccade latencies with increasing target repetitions (see Fig. S2 for this analysis). One reason for this difference might be that, in contrast to Carlisle et al., the participants in the present study always had to maintain two search templates throughout a block. This might have required continuous reactivation of each target feature in order not to forget or confuse them with the target features of previous trials, which would be distractors on the current trial. Finally, given the matched streak lengths for both-targets and one-target blocks, it is unclear how a transfer to long-term memory could explain why switch costs would selectively emerge in one-target blocks only, making it unlikely that transfer to long-term memory formed the basis for the observed effects.
The role of cognitive control in visual search
As we have argued, the present findings are in line with an explanation based on two modes of cognitive control, one proactive, one reactive. So far there has been little research on the roles of these forms of control in visual search. In a recent review, Geng (2014) argued for the involvement of proactive and reactive control in distractor suppression. According to this framework, reactive control becomes necessary in order to disengage from a distractor once observers’ attention has been captured by it, whereas proactive control may enable observers to avoid having attention captured in first place. One piece of evidence for reactive suppression of distractors is provided by the distractor positivity (Pd), an event-related potential that occurs contralateral to a suppressed location (Hickey, Di Lollo, & McDonald, 2009). There is also evidence for proactive control of distractor suppression, as observers try to adopt an attentional set against distractor locations or distractor features (e.g., Arita, Carlisle, & Woodman, 2012; Awh, Matsukura, & Serences, 2003; Poole & Kane, 2009; Reeder, Olivers, & Pollmann, 2017). Our results extend these findings by demonstrating that observers can use available information not only to avoid attending unwanted stimuli, but also to facilitate the shift of attention toward stimuli that match a current task set.
However, it is important to point out that we did not actually independently measure cognitive control. We merely assumed that observers could proactively prioritize one of two potential targets in the intertrial interval and respond reactively if the target were not there, but we have no direct evidence for this claim. To gain further insight into what actually happens before and during search, we are currently investigating the dynamics of multiple-target search with both fMRI and electroencephalographic measures. If the two modes of control indeed affect visual search in the way we propose here, neural activity related to template switching should increase prior to search display onset when both targets are available, whereas such activity should be observed after a search display onset whenever only one target is present. Furthermore, these modes might also dissociate in terms of the type and extent of the control networks involved.
Conclusion
We found switch costs whenever only one of two potential targets was available in each display, whereas no switch costs emerged when both targets were present in the search display—irrespective of the feature dimension within which the targets were defined. These results support a role for different modes of cognitive control during multiple-target search. Given free choice, observers can proactively prioritize a single target. In contrast, when only one target is available, reactive control mechanisms are necessary whenever the present target object does not match the anticipated target, irrespective of the feature dimension in which the target is defined.
Notes
To make sure that the findings were not dependent on these selection criteria, we also ran all analyses with less stringent criteria—that is, dropping the restriction that the very first saccade had to be directed toward the eventually fixated target. This led to the exclusion of only 12% of all trials. Importantly, loosening the selection criteria had no impact on the data pattern. Therefore, we report only the analyses with the more conservative criteria.
References
Allport, D. A. (1971). Parallel encoding within and between elementary stimulus dimensions. Perception & Psychophysics, 10, 104–108. https://doi.org/10.3758/BF03214327
Arita, J. T., Carlisle, N. B., & Woodman, G. F. (2012). Templates for rejection: configuring attention to ignore task-irrelevant features. Journal of Experimental Psychology: Human Perception and Performance, 38, 580–584. https://doi.org/10.1037/a0027885
Arrington, C. M., & Logan, G. D. (2004). The cost of a voluntary task switch. Psychological Science, 15, 610–615. https://doi.org/10.1111/j.0956-7976.2004.00728.x
Arrington, C. M., & Logan, G. D. (2005). Voluntary task switching: Chasing the elusive homunculus. Journal of Experimental Psychology: Learning Memory and Cognition, 31. 683–702. https://doi.org/10.1037/0278-7393.31.4.683
Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top-down versus bottom-up attentional control: A failed theoretical dichotomy. Trends in Cognitive Sciences, 16, 437–443. https://doi.org/10.1016/j.tics.2012.06.010
Awh, E., Matsukura, M., & Serences, J. T. (2003). Top-down control over biased competition during covert spatial orienting. Journal of Experimental Psychology: Human Perception and Performance, 29, 52–63. https://doi.org/10.1037/0096-1523.29.1.52
Barrett, D. J. K., & Zobay, O. (2014). Attentional control via parallel target-templates in dual-target search. PLoS ONE, 9, e86848. https://doi.org/10.1371/journal.pone.0086848
Beck, V. M., & Hollingworth, A. (2017). Competition in saccade target selection reveals attentional guidance by simultaneously active working memory representations. Journal of Experimental Psychology: Human Perception and Performance, 43, 319–335. https://doi.org/10.1037/xhp0000306
Beck, V. M., Hollingworth, A., & Luck, S. J. (2012). Simultaneous control of attention by multiple working memory representations. Psychological Science, 23, 887–898. https://doi.org/10.1177/0956797612439068
Biderman, D., Biderman, N., Zivony, A., & Lamy, D. (2017). Contingent capture is weakened in search for multiple features from different dimensions. Journal of Experimental Psychology: Human Perception and Performance, 43, 1974–1992. https://doi.org/10.1037/xhp0000422
Braver, T. S. (2012). The variable nature of cognitive control: A dual mechanisms framework. Trends in Cognitive Sciences, 16, 106–113. https://doi.org/10.1016/j.tics.2011.12.010
Carlisle, N. B., Arita, J. T., Pardo, D., & Woodman, G. F. (2011). Attentional templates in visual working memory. Journal of Neuroscience, 31, 9315–9322. https://doi.org/10.1523/JNEUROSCI.1097-11.2011
Dalmaijer, E. S., Mathôt, S., & Van der Stigchel, S. (2013). PyGaze: An open-source, cross-platform toolbox for minimal-effort programming of eyetracking experiments. Behavior Research Methods, 46, 913–921. https://doi.org/10.3758/s13428-013-0422-2
Dombrowe, I., Donk, M., & Olivers, C. N. L. (2011). The costs of switching attentional sets. Attention, Perception, & Psychophysics, 73, 2481–2488. https://doi.org/10.3758/s13414-011-0198-3
Found, A., & Müller, H. J. (1996). Searching for unknown feature targets on more than one dimension: investigating a “dimension-weighting” account. Perception & Psychophysics, 58, 88–101. https://doi.org/10.3758/BF03205479
Geng, J. J. (2014). Attentional mechanisms of distractor suppression. Current Directions in Psychological Science, 23, 147–153. https://doi.org/10.1177/0963721414525780
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009). Electrophysiological indices of target and distractor processing in visual search. Journal of Cognitive Neuroscience, 21, 760–775. https://doi.org/10.1162/jocn.2009.21039
Houtkamp, R., & Roelfsema, P. R. (2009). Matching of visual input to only one item at any one time. Psychological Research, 73, 317–326. https://doi.org/10.1007/s00426-008-0157-3
Huang, L., & Pashler, H. (2007). A Boolean map theory of visual attention. Psychological Review, 114, 599–631. https://doi.org/10.1037/0033-295X.114.3.599
Juola, J. F., Botella, J., & Palacios, A. (2004). Task-and location-switching effects on visual attention. Attention, Perception, & Psychophysics, 66, 1303–1317. https://doi.org/10.3758/BF03195000
Kristjánsson, Á., & Campana, G. (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72, 5–18. https://doi.org/10.3758/APP.72.1.5
Krummenacher, J., Müller, H. J., & Heller, D. (2001). Visual search for dimensionally redundant pop-out targets: Evidence for parallel-coactive processing of dimensions. Attention, Perception, & Psychophysics, 63, 901–917. https://doi.org/10.3758/BF03194446
Luck, S. J., & Vogel, E. K. (1997). The capacity of visual working memory for features and conjunctions. Nature, 390, 279–281. https://doi.org/10.1038/36846
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. role of features. Memory & Cognition, 22, 657–672. https://doi.org/10.3758/BF03209251
Mathôt, S., Schreij, D., & Theeuwes, J. (2012). OpenSesame: An open-source, graphical experiment builder for the social sciences. Behavior Research Methods, 44, 314–324. https://doi.org/10.3758/s13428-011-0168-7
Menneer, T., Barrett, D. J. K., Phillips, L., Donnelly, N., & Cave, K. R. (2007). Costs in searching for two targets: Dividing search across target types could improve airport security screening. Applied Cognitive Psychology, 21, 915–932. https://doi.org/10.1002/acp.1305
Monsell, S. (2003). Task switching. Trends in Cognitive Sciences, 7, 134–140. https://doi.org/10.1016/S1364-661300028-7
Morey, R. D. (2008). Confidence intervals from normalized data: A correction to Cousineau (2005). Tutorials in Quantitative Methods for Psychology, 4, 61–64. https://doi.org/10.3758/s13414-012-0291-2
Müller, H. J., Heller, D., & Ziegler, J. (1995). Visual search for singleton feature targets within and across feature dimensions. Attention, Perception, & Psychophysics, 57, 1–17. https://doi.org/10.3758/BF03211845
Oberauer, K. (2002). Access to information in working memory: Exploring the focus of attention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 28, 411–421. https://doi.org/10.1037/0278-7393.28.3.411
O’Craven, K. M., Downing, P. E., & Kanwisher, N. (1999). fMRI evidence for objects as the units of attentional selection. Nature, 401, 584–587. https://doi.org/10.1038/44134
Olivers, C. N. L., & Humphreys, G. W. (2003). Attentional guidance by salient feature singletons depends on intertrial contingencies. Journal of Experimental Psychology: Human Perception and Performance, 29, 650–657. https://doi.org/10.1037/0096-1523.29.3.650
Olivers, C. N. L., & Meeter, M. (2006). On the dissociation between compound and present/absent tasks in visual search: Intertrial priming is ambiguity driven. Visual Cognition, 13, 1–28. https://doi.org/10.1080/13506280500308101
Olivers, C. N. L., & Meeter, M. (2008). Feature priming in visual search does not depend on the dimensional context. Visual Cognition, 16, 785–803. https://doi.org/10.1080/13506280701428658
Olivers, C. N. L., Peters, J., Houtkamp, R., & Roelfsema, P. R. (2011). Different states in visual working memory: When it guides attention and when it does not. Trends in Cognitive Sciences, 15, 327–334. https://doi.org/10.1016/j.tics.2011.05.004
Ort, E., Fahrenfort, J. J., & Olivers, C. N. L. (2017). Lack of free choice reveals the cost of having to look for more than one object. Psychological Science, 28, 1137–1147. https://doi.org/10.1177/0956797617705667
Poole, B. J., & Kane, M. J. (2009). Working-memory capacity predicts the executive control of visual search among distractors: The influences of sustained and selective attention. Quarterly Journal of Experimental Psychology, 62, 1430–1454. https://doi.org/10.1080/17470210802479329
Quinlan, P. T., & Humphreys, G. W. (1987). Visual search for targets defined by combinations of color, shape, and size: an examination of the task constraints on feature and conjunction searches. Perception & Psychophysics, 41, 455–472. https://doi.org/10.3758/BF03203039
Reeder, R. R., Olivers, C. N. L., & Pollmann, S. (2017). Cortical evidence for negative search templates. Visual Cognition, 25, 278–290. https://doi.org/10.1080/13506285.2017.1339755
Roelfsema, P. R., Lamme, V. A. F., & Spekreijse, H. (1998). Object-based attention in the primary visual cortex of the macaque monkey. Nature, 395, 376–381. https://doi.org/10.1038/26475
Roper, Z. J. J., & Vecera S. P. (2012). Searching for two things at once: Establishment of multiple attentional control settings on a trial-by-trial basis. Psychonomic Bulletin & Review, 19, 1114–21. https://doi.org/10.3758/s13423-012-0297-8
Singmann, H., Bolker, B., & Westfall, J. (2015). afex: Analysis of factorial experiments (R package version 0.15-2). Retrieved from https://cran.r-project.org/web/packages/afex/index.html
Stroud, M. J., Menneer, T., Cave, K. R., Donnelly, N., & Rayner, K. (2011). Search for multiple targets of different colours: Misguided eye movements reveal a reduction of colour selectivity. Applied Cognitive Psychology, 25, 971–982. https://doi.org/10.1002/acp.1790
Treisman, A. M., & Gelade, G. (1980). A feature-integration theory of attention. Cognitive Psychology, 12, 97–136. https://doi.org/10.1016/0010-0285(80)90005-5
Wheeler, M. E., & Treisman, A. M. (2002). Binding in short-term visual memory. Journal of Experimental Psychology: General, 131, 48–64. https://doi.org/10.1037/0096-3445.131.1.48
Williams, L. (1967). The effects of target specification on objects fixated during visual search. Acta Psychologica, 27, 355–360. https://doi.org/10.1016/0001-6918(67)90080-7
Wolfe, J. M. (1994). Guided Search 2.0 A revised model of visual search. Psychonomic Bulletin & Review, 1, 202–238. https://doi.org/10.3758/BF03200774
Wolfe, J. M. (2012). Saved by a log: How do humans perform hybrid visual and memory search? Psychological Science, 23, 698–703. https://doi.org/10.1177/0956797612443968
Wolfe, J. M., Butcher, S. J., Lee, C., & Hyle, M. (2003). Changing your mind: On the contributions of top-down and bottom-up guidance in visual search for feature singletons. Journal of Experimental Psychology: Human Perception and Performance, 29, 483–502. https://doi.org/10.1037/0096-1523.29.2.483
Wolfe, J. M., Cave, K. R., & Frenzel, S. L. (1989). Guided search: An alternative to the feature integration model for visual search. Journal of Experimental Psychology: Human Perception and Performance, 15, 419–433. https://doi.org/10.1037/0096-1523.15.3.419
Wolfe, J. M., & Horowitz, T. S. (2004). What attributes guide the deployment of visual attention and how do they do it? Nature Reviews Neuroscience, 5, 495–501. https://doi.org/10.1038/nrn1411
Wolfe, J. M., Yu, K. P., Stewart, M. I., Shorter, A. D., Friedman-Hill, S. R., & Cave, K. R. (1990). Limitations on the parallel guidance of visual search: Color × Color and Orientation × Orientation conjunctions. Journal of Experimental Psychology: Human Perception and Performance, 16, 879–892. https://doi.org/10.1037/0096-1523.16.4.879
Author information
Authors and Affiliations
Contributions
This work was supported by Open Research Area Grant 464-13-003 from the Netherlands Organization for Scientific Research, and by European Research Council Consolidator Grant ERC-2013-CoG-615423 to C.N.L.O. The authors declare that they have no conflicts of interest with respect to their authorship or the publication of this article.
Corresponding author
Electronic supplementary material
All data and materials have been made publicly available via the Open Science Framework and can be accessed at www.osf.io/d9er6.
ESM 1
(DOCX 306 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Ort, E., Fahrenfort, J.J. & Olivers, C.N.L. Lack of free choice reveals the cost of multiple-target search within and across feature dimensions. Atten Percept Psychophys 80, 1904–1917 (2018). https://doi.org/10.3758/s13414-018-1579-7
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-1579-7