
Abstract
This study investigated whether initial self-control exertion by dogs would affect behavioral approach toward an aggressive threat. Dogs were initially required to exert self-control (sit still for 10 min) or not (caged for 10 min) before they were walked into a room in which a barking, growling dog was caged. Subject dogs spent 4 min in this room but were free to choose where in the room they spent their time. Approaching the unfamiliar conspecific was the predisposed response, but it was also the riskier choice (Lindsay, 2005). We found that following the exertion of self-control (in comparison with the control condition), dogs spent greater time in proximity to the aggressor. This pattern of behavior suggests that initial self-control exertion results in riskier and more impulsive decision making by dogs.
Similar content being viewed by others

The potential for danger is ubiquitous. To avoid danger, people often exert self-control over their behavior (Baumeister, 1998; Baumeister, Heatherton & Tice, 1994). When people fail to exert self-control and behave more impulsively, they may unintentionally put themselves in harm’s way (Freeman & Muraven, 2010). Pedestrians jaywalk across busy streets, children stick objects into electrical outlets, and teenagers join dangerous gangs. The failure to exert self-control and avoid these dangerous activities is likely affected by many individual variables (e.g., demographics, personality). However, a common mechanism that may be responsible for human and nonhuman self-control vigor may also play a role (Miller, Pattison, DeWall, Rayburn-Reeves & Zentall, 2010). The present research tested this hypothesis by examining whether dogs approach dangerous situations when their ability to exert self-control is compromised.
Research with human and nonhuman animals suggests that self-control relies on a limited resource (Baumeister & Heatherton, 2004; Miller et al., 2010). Exerting self-control depletes this resource, and once depleted, subsequent efforts to control behavior become impaired. For example, when humans control their impulse to eat fresh cookies (in comparison to when they inhibit eating radishes), they then persist for a shorter time on an unsolvable puzzle task (Baumeister, Bratslavsky, Muraven & Tice, 1998). Dogs behave similarly. When dogs control their physical movement (in comparison with when self-control is not needed because they are physically constrained by a cage), they persist for a shorter duration on a subsequent unsolvable puzzle task (Miller et al., 2010).
Extensive research with humans suggests that this phenomenon is domain general, suggesting that tasks that require self-control negatively affect performance on a wide variety of subsequent tasks (for a review, see Baumeister, Schmeichel & Vohs, 2007). Decision making, for example, is negatively affected by initial self-control exertion. Depleted subjects, as compared with their nondepleted counterparts, take more risks and gamble more (Bruyneel, DeWitte, Franses & Dekimpe, 2009; Freeman & Muraven, 2010; Molet, Miller, Laude, Kirk, Manning & Zentall, 2012). To date, however, there is no evidence that initial self-control exertion affects subsequent behavior in more than one domain for nonhuman animals, nor is there evidence that depleted humans or nonhuman animals are more likely to inadvertently subject themselves to risks that may result in physical harm.
Some recent evidence suggests this possibility, because self-regulatory depletion increases approach motivation (Schmeichel, Harmon-Jones & Harmon-Jones, 2010) and behaviors with an approach-related motivational direction, such as aggression (Denson, Pedersen, Friese, Hahm & Roberts, 2011; DeWall, Baumeister, Stillman & Gailliot, 2007; Finkel, DeWall, Slotter, Oaten & Foshee, 2009). Consequently, the purpose of the present investigation was to determine whether self-regulatory depletion has domain-general consequences on nonhuman animal behavior and to examine the likelihood that depletion would cause dogs to put themselves in harm’s way by approaching a physically threatening target. We chose dogs, in part, because, like humans, they are highly social animals that establish social dominance hierarchies and need to be sensitive to social cues provided by other members of their species.
We adopted a two-task procedure used widely in the human self-control literature (Baumeister, Bratslavsky, Muraven & Tice, 1998). Dogs were tested twice, individually, and we varied the requirements of the first task between sessions. In the first task, either the dog was required to sit still for 10 min (self-control depletion), or the movement of the dog was constrained by placing it inside of a cage for the same duration (control). Next, the dog was brought into a room in which it encountered a caged, barking, and growling dog. Dogs spent time in this room, but their behavior was unconstrained. Thus, dogs could choose to spend their time near the aggressor, or they could stay farther away. Although dogs are predisposed to approach and investigate unknown conspecifics (Lindsay, 2000), in this context, it was the more dangerous thing to do. Greater proximity to a confined aggressive dog, despite the confinement, is associated with a greater risk of an aggressive encounter (American Veterinary Medical Association, 2011; Lindsay, 2001, 2005; Sacks, Sattin & Bonzo, 1989). Consequently, staying near the aggressor was defined as the riskier (more impulsive) choice, and avoidance was defined as the safer (less impulsive) choice. We predicted that self-control depletion, as compared with the control condition, would increase approach-related behavior toward the threatening dog.
Method
Subjects
We recruited 10 dogs (Canis familiaris; 4 males, 6 females) ranging from 12 to 120 months of age (M = 48.8 months). All dogs belonged to private owners, would immediately approach a friendly caged dog, and had been trained to maintain an out-of-sight sit–stay for 10 min. They had also been trained to remain calm inside a cage for as long as 6 h.
Apparatus
A bath mat was placed on the floor in front of an empty dog cage (1.2 m long × 0.8 m wide × 0.9 m high) that was surrounded by a ProSelect™ exercise pen (see Fig. 1). The dogs sat on this mat during the self-control manipulation. This mat was placed inside a second dog cage (0.9 m long × 0.6 m wide × 0.7 high) at the same location during the control condition. A mirror was placed strategically on the wall so that the experimenter could watch the dogs from outside of the room through a small opening in the door. To increase the difficulty of the self-control depletion phase, an electronic “hamster” (Zhu Zhu pets®) was placed inside an Adventure Ball™ and was activated inside the room during the self-control depletion phase (see Fig. 1).

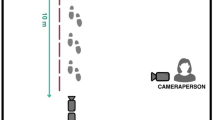
The experimental room as it was set up for the self-control manipulation that preceded the impulsivity test
A highly dominant dog (an 11-year-old female bull terrier) with a disposition for guarding territory was placed in a dog cage (1.2 m long × 0.8 m wide × 0.9 m high) that was surrounded by the ProSelect™ exercise pen. This dog would bark and growl continuously whenever it was confined and another dog was visible. The intensity of this display was greater when its owner was close by, suggesting that the dog included its owner as part of its territory to defend (Borchelt, 1983; Lindsay, 2001). For this reason, the owner (O) stood next to the dog during testing.
For safety reasons, the pen that was placed around the cage provided an additional distance of 0.3 m between the aggressive dog and the subject dog. The room (3.9 m long × 3.8 m wide) was demarcated into zones by lines made with Scotch® masking tape (2.5 cm wide; see Fig. 2). The first of four lines was 61 cm from the screen (zone 1), and zone was 61 cm deep (zones 2, 3, and 4). Because there were doors in zones 1 and 4 and standing near a door should be considered an escape behavior rather than an approach behavior, we chose to define this zone separately. The dog was considered in door space if its front two feet were within 30 cm of the door and the dog's body was pointing in the direction of the door.

The experimental room as it was set up for the impulsivity test
In the first phase of the experiment, the door (B) was left slightly ajar; however, both doors were closed during the second phase of the experiment. Two video cameras were used to record each testing session; one was mounted on a tripod to the left of the aggressive dog, and an experimenter who was to the right of the dog operated the other camera.
Procedure
Pretesting
At least 1 week before testing, all subject dogs were introduced to a friendly caged dog. All subject dogs approached the caged dog upon being released from leashed restraint. The purpose of this testing was to ensure that subject dogs were predisposed to approach an unknown conspecific and that some level of inhibition would be required for avoidance. In addition, subject dogs were leashed and were exposed to the Zhu Zhu® hamster to ensure that they were interested in but not frightened by it when it moved. This assessment eliminated the possibility that fear would confound the self-control manipulation.
Testing
All dogs were tested with both conditions of the self-control manipulation (order was counterbalanced). In the self-control depletion condition, the dog was cued to “sit” and “stay” by Experimenter 1 (E1). Following the cue, E1 activated the Zhu Zhu® hamster and exited the room through door A while the dog maintained its position. The hamster was used to increase the difficulty of the self-control task. With the door slightly ajar, E1 watched the dog (without being seen by the dog) via a strategically placed mirror. The dog was allowed to keep visual track of the hamster but if the dog moved from its position, E1 returned and gave the sit–stay cue again. A second experimenter (E2), who stood outside of the testing room (behind door A), recorded the number of cues and the time at which each cue was given. The dog remained alone in the room with the electronic hamster for a total of 10 min.
When the dog was released, it was given a small piece of wiener (1 g), was moved to an adjacent room, and was praised for 30 s by E1. After the subject dog left the testing room, the aggressive dog was led into the newly vacant room through door B by its owner (O) who was blind to the self-control manipulation. The aggressive dog was placed inside the cage, and O stood next to the cage behind the exercise pen during testing. Once the aggressive dog was situated, E1 entered the room with the subject dog (which caused the aggressive dog to start barking and growling), quietly walked the subject dog across the room on leash, and unleashed it at a predesignated spot without further interaction (black dot in Fig. 2). E1 then filmed the subject dog’s behavior for 4 min while quietly standing still and looking at the dog only via the screen of the digital camera. At the end of 4 min, E1 leashed the subject dog and removed it from the experimental room through door B.
In the control condition, E1 placed the subject dog inside a dog cage and was told by E2 to return to the dog and “recue” it to get inside of the cage at the same times she had previously cued the dog during the self-control condition. If the dogs’ first test session was in the control condition, it was revisited by E1 three times at minutes 1, 5, and 7 during the 10 min. This was the average number of revisits required in a previous experiment (Miller et al., 2010). During the control condition, the electronic hamster was inside the room but was not activated. Pilot research indicated that the hamster attracted attention from dogs during the sit–stay but that, when activated while caged, it caused some dogs to whine, circle, and paw. The remaining procedural variables were held constant between conditions. Dogs were tested between the hours of 09:00 and 15:00. A video clip displaying the dogs’ behavior during the control manipulation can be found at http://www.youtube.com/watch?v=849mWDrXnC8.
An observer, who was blind to the subjects’ condition, used a digital stopwatch to time the duration that subject dogs spent in each zone. A second blind observer coded 10% of the observations to obtain a reliability measure. There was a positive Pearson correlation between the two observers, r = .99, p < .01.
Results
The dogs responded differently in the two conditions. The dogs generally spent more time in zone 1 and near the doors, especially following the self-control condition. The proportion of time spent in each zone and near the door appears in Fig. 3.

Mean proportion of time in the different zones as a function of self-control condition (self-control vs. control)
A two-way repeated measures analysis of variance (ANOVA) analyzed the differences of time spent in the zones (+doors) across conditions (self-control, control). Time spent in the room did not differ as a function of condition because total time was equated, but the time spent in the zones was significantly different, F(4, 36) = 5.06, p < .01, and the dispersion of time spent in zones was significantly affected by self-control condition, F(4, 36) = 2.56, p = .05. Subsequent analyses were run to examine the observed differences. A one-way repeated measures ANOVA examining the effect of zone for the self-control condition found that there was a very reliable effect of zone following self-control exertion, F(4, 51) = 6.08, p < .01, but not following the control condition, F(4, 51) = 1.88, p = .13. When the simple effects of zone within each condition (self-control, control) were analyzed, it was observed that following self-control exertion, dogs reliably spent more time in zone 1 than in 2, 3, and 4, F(9, 36) = 13.29, 15.75, 16.17, respectively, all ps < .01, but not more time than near the doors, F(9, 36) = 3.67, p = .06. Following the control condition, dogs did not spend a significantly different amount of time between zones 1 and 2, F(9, 36) = 2.13, p = .15, but they did spend a significantly greater time in zone 1 than in zones 3 and 4, F(9, 36) = 5.67, 5.54, all ps = .02. Here again, time spent in zone 1 did not differ from time spent at the doors, F(9, 36) = 1.95, p = .17.
Planned comparisons using two-tailed correlated samples t-tests were performed on the difference scores in each zone between the two conditions. The difference was significant in zone 1, t(9) = 3.11, p = .01 (dogs spent more time in zone 1 following the self-control condition), and in zone 2, t(9) = 2.39, p = .04 (dogs spent more time in zone 2 following the control condition), but it was not significant in either of the other two zones (or near the doors), all ts < 1.2. As was predicted, self-control depletion caused dogs to spend a greater percentage of time in the zone nearest to the aggressive dog (58.9%), as compared with the control condition (41.8%). The data from individual dogs in both the self-control and control conditions appear in Fig. 4.

Proportion of time that individual dogs spent in the area nearest the threatening conspecific (zone 1) as a function of self-control condition (self-control vs. control)
Discussion
Avoiding danger enhances an animal’s ability to survive and reproduce. Yet there are often occasions when the need to avoid danger is paired with a natural tendency to approach. To keep out of harm’s way, animals override their natural impulse to approach in order to remain safe and secure. When animals have limited self-control resources, they may make more impulsive decisions that put them in harm’s way. The present experiment examined whether initial exertion of self-control would increase impulsivity in dogs, resulting in risky decisions.
Our results supported this prediction. When dogs were depleted, as compared with when they were not, they were less able to inhibit their predisposed approach behaviors. As a result, dogs approached an aggressive dog more when depleted than when nondepleted.
Our interpretation of the present results rests on the assumption that it is dangerous to approach a confined aggressive dog and that subject dogs were risking their safety by approaching. Although in the context of the present experiment, strict precautions ensured the safety of all of the dogs, confinement is not fail-proof in the natural world, and it is quite possible that a confined aggressor could escape and attack. Mail carriers are often attacked by presumably confined dogs, as are children, and a significant percentage of pet-related human fatalities result when a restrained aggressive dog is approached (Sacks et al., 1989; U.S. Postal Service, 2011). Moreover, moving away from an aggressor defending its territory reduces the motivation for attack (Lindsay, 2001). In the context of the present experiment, experimenters observed that the aggressor displayed a more intense threat display when subject dogs were near the fence and the intensity decreased as subject dogs moved farther away. This observation is similar to others regarding territorial behavior and confinement (Calhoun, 1962; Klopfer, 1969; Lindsay, 2001; Pettijohn, Davis & Scott, 1980). It is also evidence that approaching was relatively more risky than avoiding the caged aggressor.
Another way of looking at the dogs’ behavior is as an increase in counter aggression, provoked by the confined aggressor. This complementary hypothesis is founded on research demonstrating that humans, who typically suppress emotionally driven aggressive responses, are more likely to retaliate aggressively when they have depleted their self-control. More specifically, when students initially deplete their self-control by inhibiting their consumption of a donut (but not a radish), they are less able to control their aggressive behavior when they are subsequently provoked (i.e., they are negatively evaluated on a previously written essay). Depleted students retaliate by adding more hot sauce to food intended for the essay evaluator. Similarly, when students are initially required to control their attention (inhibit reading words displayed to them) and are subsequently given a negative essay evaluation, they are more likely to administer aversive noises to the essay evaluator. Furthermore, following self-control exertion, students are more likely to report the desire to inflict physical harm on someone who provokes them (DeWall et al., 2007).
In the context of the present experiment, it is difficult to delineate the role that increased aggression may have played. Increased approach behavior may reflect counter aggression, or it may reflect an increase in approach motivation. Self-regulatory depletion increases approach motivation in humans in the absence of aggressive intentions and, thus, may have also contributed to the pattern of behavior we observed in dogs (Schmeichel et al., 2010).
The commonality between human and nonhuman animals is great, and the present research is further evidence that human self-control has phylogenetic roots. It is also further evidence that a phenomenon (i.e., depletion) once believed to be uniquely human can be modeled with dogs. Such modeling may have great empirical value, since it may provide greater insight into the physiological and neurobiological processes that affect self-control vigor. Research with animal models will not supplant that on cognitive factors involved in human self-regulation, but it will augment our understanding of the fundamental and biological rudiments of a phenomenon that is clearly multifactorial in nature. Therefore, we believe that social psychology will benefit by incorporating work with nonhuman animals to extend existing theories.
References
American Veterinary Medical Association. (2011). National dog bite prevention week (press release). Retrieved from http://www.avma.org/press/releases/110512-NDBPW-prevention-campaign-pf.asp
Baumeister, R. F. (1998). The self. In D. T. Gilbert, S. T. Fiske, & G. Lindzey (Eds.), Handbook of social psychology (4th ed., pp. 680–740). New York: McGraw-Hill.
Baumeister, R. F., Bratslavsky, E., Muraven, M., & Tice, D. M. (1998). Ego depletion: Is the active self a limited resource? Journal of Personality and Social Psychology, 74, 1252–1265.
Baumeister, R. F., & Heatherton, T. (2004). Self-regulation failure: An overview. In R. M. Kowalski & M. R. Leary (Eds.), The interface of social and clinical psychology: Key readings (pp. 51–69). New York: Psychology Press.
Baumeister, R. F., Heatherton, T. F., & Tice, D. M. (1994). Losing control: How and why people fail at self-regulation. San Diego, CA: Academic Press.
Baumeister, R. F., Schmeichel, B. J., & Vohs, K. D. (2007). Self-regulation. In A. W. Kruglanski & E. T. Higgins (Eds.), Social psychology: Handbook of basic principles (pp. 516–539). New York: Guildford.
Borchelt, P. L. (1983). Aggressive behavior of dogs kept as companion animals: Classification and influence of sex, reproductive status and breed. Applied Animal Ethology, 10, 45–61.
Bruyneel, S. D., DeWitte, S., Franses, P. H., & Dekimpe, M. G. (2009). I felt low and my purse feels light: Depleting mood regulation attempts affect risk decision making. Journal of Behavioral Decision Making, 22, 153–170.
Calhoun, J. B. (1962). Population density and social pathology. Scientific American, 206, 139–148.
Denson, T. F., Pedersen, W. C., Friese, M., Hahm, A., & Roberts, L. (2011). Understanding impulsive aggression: Angry rumination and reduced self-control capacity are mechanisms underlying the provocation–aggression relationship. Personality and Social Psychology Bulletin. Advance online publication. doi:10.1177/0146167211401420
DeWall, C. N., Baumeister, R. F., Stillman, T. F., & Gailliot, M. T. (2007). Violence restrained: Effects of self-regulatory capacity and its depletion on aggressive behavior. Journal of Experimental Social Psychology, 43, 62–76.
Finkel, E. J., DeWall, C. N., Slotter, E. B., Oaten, M., & Foshee, V. A. (2009). Self-regulatory failure and intimate partner violence perpetration. Journal of Personality and Social Psychology, 97, 483–499.
Freeman, N., & Muraven, M. (2010). Self-control depletion leads to increased risk taking. Social Psychological Science, 2, 175–181.
Klopfer, P. H. (1969). Habitats and territories: A study of the use of space by animals. New York: Basic.
Lindsay, S. R. (2000). Handbook of applied dog behavior and training: Vol. 1. Adaptation and learning. Ames: Iowa State University Press.
Lindsay, S. R. (2001). Handbook of applied dog behavior and training: Vol. 2. Etiology and assessment of behavior problems. Ames: Iowa State University Press.
Lindsay, S. R. (2005). Handbook of applied dog behavior and training: Vol. 3. Procedures and protocols. Ames: Iowa State University Press.
Miller, H. C., Pattison, K. P., DeWall, C. N., Rayburn-Reeves, R., & Zentall, T. R. (2010). Self-control without a self? Common self-control processes in humans and dogs. Psychological Science, 21, 534–538.
Molet, M., Miller, H. C., Laude, J. R., Kirk, C., Manning, B., & Zentall, T. R. (2012). Decision-making by humans in a behavioral task: Do humans, like pigeons, show suboptimal choice? Learning & Behavior. Advance online publication. doi:10.3758/s13420-012-0065-7
Pettijohn, T. F., Davis, K. L., & Scott, J. P. (1980). Influence of living area space on agonistic interacion in Telomian dogs. Behavioral and Neural Biology, 28, 343–349.
U.S. Postal Service (2011). Postal news (Press Release 11-050). Retrieved from about.usps.com/news/national-releases/2011/pr11_050.pdf.
Sacks, J. J., Sattin, R. W., & Bonzo, S. E. (1989). Dog bite-related fatalities from 1979 through 1988. Journal of the American Medical Association, 262, 1489–1492.
Schmeichel, B. J., Harmon-Jones, C., & Harmon-Jones, E. (2010). Exercising self-control increases approach motivation. Journal of Personality and Social Psychology, 99, 162–172.
Author Note
We thank Byron Nelson for his help with data analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Miller, H.C., DeWall, C.N., Pattison, K. et al. Too dog tired to avoid danger: Self-control depletion in canines increases behavioral approach toward an aggressive threat. Psychon Bull Rev 19, 535–540 (2012). https://doi.org/10.3758/s13423-012-0231-0
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-012-0231-0