
Abstract
Key Message
We present data from a new delimited 1-ha forest plot on the Sierra Nevada de Santa Marta in Colombia and describe its tree diversity, composition, and structure. This data can be used in regional analyses and help to refine remote sensing models. Future censuses will provide data on the demographics of rare, endangered, and endemic trees that can be used for their conservation. Dataset access is at https://doi.org/10.15472/ftwol2. Associated metadata are available at https://metadata-afs.nancy.inra.fr/geonetwork/srv/fre/catalog.search#/metadata/a4a7855f-4cb2-459b-a1a2-287bd1e6d587.
Similar content being viewed by others


1 Background
The Sierra Nevada de Santa Marta (SNSM) is an isolated mountain on Colombia’s Caribbean coast. Rising to 5775 m asl, this immense massif is the tallest peak in Colombia and the tallest coastal mountain in the world. The SNSM is separated from the Andes mountains by hot and dry lowland Caribbean savannas to the southeast and by a large wetland, the Ciénaga Grande de Santa Marta, to the southwest. Owing to its unique location, isolation, and complex topography, the mountain is home to a mosaic of distinct ecosystems with high levels of endemism (Adams 1973; Carbono and Lozano-Contreras 1997; Cardona and Ojeda 2010).
Unfortunately, our understanding of the mountain’s flora is incomplete. Although there have been important botanical expeditions to the SNSM (Van der Hammen and Ruiz 1984; Ayers and Boufford 1998), no study has implemented a standardized tree plot with which to compare the cloud forests of SNSM to others in the region (ForestPlots.net et al. 2021). Therefore, our knowledge of the diversity, composition, and structure of tree communities on the SNSM remains severely limited. To address this problem, we installed a permanent 1-ha forest plot on the SNSM, and we here present baseline data from the plot. Our work represents the first standardized tree plot above 600 m asl on the SNSM and will contribute to our understanding of both local and regional cloud forest diversity and composition.
2 Methods
2.1 Study site
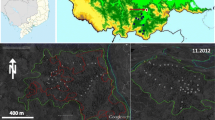
The 1-ha plot is located on Cerro Kennedy, a high point reaching 2830 m asl on the northwest side of the SNSM. On the southern face of Cerro Kennedy, in Reserva El Dorado, is a strip of relict old-growth forest totaling just over 250 ha. The plot is located in this remnant forest at 2200-m elevation (11° 6′4.37″ N, 74° 2′22.20″ W, Fig. 1). Mean annual temperature is ~ 15 °C, and mean annual precipitation is ~ 1900 mm.

Map showing the location of the 1-ha plot in Colombia and on the Sierra Nevada de Santa Marta
2.2 Plot demarcation
The specific location of the plot was chosen pseudo-randomly, avoiding difficult terrain and steep topography, in a location easy to access from the trail, and ensuring that each plot edge was at least 100 m from a forest edge to avoid as many edge effects as possible (Laurance et al. 2002). We demarcated the plot boundaries with PVC tubes every 10 m following cardinal directions with a compass. We then installed 100 10 × 10 m subplots following the same methods. The shape of the plot is irregular due to a cliff we encountered in the southwest corner and is steep, with an ENE to WSW slope exceeding 30°. Because each 10-m interval was demarcated using a planar projection and the ground distance was corrected with a digital clinometer, the total ground area of the plot is approximately 1.21 ha.
2.3 Tree measurements
We tagged, measured, mapped, and identified every stem ≥ 10-cm DBH following standard protocols (Lopez-Gonzalez et al. 2011; Phillips et al. 2018). For trees with buttressed trunks or other irregularities, diameter was measured above the irregularities, and measurement height was noted. We collected voucher specimens for species identifications and stored them in the herbarium at the Cartagena Botanical Garden “Guillermo Piñeres” (JBGP).
We also scored each tree for liana infestation, sun exposure, and canopy damage. Liana infestation scores were 0 for trees without lianas and 1, 2, 3, or 4 for trees with 1–25%, 26–50%, 51–75%, or 76–100% of their canopy covered by lianas, respectively (Clark & Clark 1990). Sun exposure scores were from 1 to 5, with 1 for plants lacking direct sunlight except during sun flecks and 5 for canopy emergents (Dawkins 1978). Crown damage scores were 0 for trees with intact crowns and 1, 2, 3, or 4 for trees with 1–25%, 26–50%, 51–75%, or 76–100% of the crown broken, respectively. Finally, we estimated tree height.
2.4 Access to the data and metadata description
The datasets generated during the present study are available in the SiB Colombia (Contreras et al. 2024), URL: https://ipt.biodiversidad.co/sib/resource?r=parcelasnsm#anchor-downloads, DOI: https://doi.org/10.15472/ftwol2. Metadata files are included with this dataset and also accessible at https://metadata-afs.nancy.inra.fr/geonetwork/srv/fre/catalog.search#/metadata/a4a7855f-4cb2-459b-a1a2-287bd1e6d587.
2.5 Technical validation
All variables were examined for inconsistencies. Any outliers, impossible tree scores, and mapping errors were revised by checking with field sheets and confirming with the field team. We made figures to summarize and visualize the data to ease error identification (Figs. 2–3). Species identifications that were made in the field were confirmed or refined by comparing our collections to physical material in the Jardín Botánico de Cartagena “Guillermo Piñeres” herbarium (JBPG). When physical material was not available, we reviewed digital herbarium specimens in the online repositories of the New York Botanical Garden (NY), Missouri Botanical Garden (MO), Field Museum Herbarium (F), the Royal Botanic Gardens Kew (K), and the Herbario Forestal de la Universidad Distrital “Francisco José de Caldas” (UDBC).

A Stem map of the 1-ha plot. Each dot is a stem, with the size of the dot corresponding to its diameter. The four most common species are color coded. B Bar graph of the most common species, ranked by number of individuals. The four most common species are color coded as in A. C Bar graph of the 10 most species-rich families, ranked by number of species

Histogram of stem diameters in the 1-ha plot. The blue dashed line indicates the mean diameter (23.3 cm) at breast height
In total, we measured, tagged, mapped, and identified a total of 924 stems representing 846 individual trees and 85 species in 41 families. Four species represent nearly half of total stems, and four families account for 32% of all species (Table 1, Fig. 2). We calculated three diversity metrics including Shannon, Simpson, and inverse Simpson indices, which were 3.36, 0.93, and 14.3, respectively. Even though each metric is calculated differently, they all illustrate relatively high diversity within the plot.
Using measured tree diameters (D), we calculated the basal area of each stem as (D/2)^2*π. We then assigned wood density for each species from a wood density database (Zanne et al. 2009). When a species-level wood density was not available, we assigned a species the genus- or family-level wood density average instead. We then estimated biomass for the entire stand using an allometric model via the R package BIOMASS (Chave et al. 2014; Réjou-Méchain et al. 2017). Tree diameters range from 10 to 123.5 cm, with a mean diameter of 23.3 cm (Fig. 3). The total basal area for the entire plot is 55.96 m2. The four species with the largest basal area are Pouteria espinae (18% of total basal area, Table 1), Ficus insipida (Moraceae, 11%), Chrysochlamys colombiana (8%), and Calatola costaricensis (7%). Total plot biomass is ~ 451 t.
2.6 Reuse and potential limits
Plot data is essential for analyses of species composition across space and through time. Our plot data will be uploaded into ForestPlots.net (Lopez-Gonzalez et al. 2011), a pantropical network of tree plots which facilitates studies that investigate patterns of tree species composition and diversity. Data on tree-level liana infestation, sun exposure, and crown damage in 1-ha plots is scarce in tropical montane cloud forests, so our plot will be useful when doing cross-site examinations of these variables. We plan to do a full recensus of the plot in 2028 and every 5 years thereafter in order to characterize compositional changes and calculate growth and mortality rates through time.
Data on the structure and composition of trees such as those presented here are important components of earth systems modeling. Indeed, forest plots help to inform and calibrate remote sensing estimations of aboveground biomass and carbon sequestration (Chave et al. 2019). Thus, our ground-based data can be used in future studies that estimate biomass on large spatial scales using remote sensing techniques.
Some species in our dataset are rare, endangered, and/or endemic (e.g., Pouteria espinae, the most common species in the plot) to the SNSM. Therefore, the occurrence data (and future growth, mortality, and recruitment data) of these species can be useful for their management, for protection, and for future assessments for the Red List of the International Union for the Conservation of Nature (IUCN).
Availability of data and materials
The datasets generated during the current study are available as a Darwin Core Archive in the SiB Colombia, DOI: https://doi.org/10.15472/ftwol2, URL: https://ipt.biodiversidad.co/sib/resource?r=parcelasnsm#anchor-downloads. The data can also be accessed through the Global Biodiversity Information Facility (GBIF) at https://www.gbif.org/dataset/caf76652-9bb6-4d47-8a67-4c473cf56130.
References
Adams M (1973) Ecological zonation and the butterflies of the Sierra Nevada de Santa Marta, Colombia. J Nat Hist 7:699–718
Ayers TJ, Boufford DE (1998) Index to the vascular plant types collected by H. H. Smith near Santa Marta Colombia. Brittonia 40:400–432
Carbono E, Lozano-Contreras G (1997) Endemismos y otras singularidades de la Sierra Nevada de Santa Marta, Colombia. Posibles causas de origen y necesidad de conservarlos. Rev Acad Colombiana Ciencias 21:409–419
Cardona A, Ojeda GY (2010) Special volume: geological evolution of the Sierra Nevada de Santa Marta and adjacent basins, Colombian Caribbean region
Chave J, Réjou-Méchain M, Búrquez A, Chidumayo E, Colgan MS, Delitti WBC, Duque A, Eid T, Fearnside PM, Goodman RC, Henry M, Martínez-Yrízar A, Mugasha WA, Muller-Landau HC, Mencuccini M, Nelson BW, Ngomanda A, Nogueira EM, Ortiz-Malavassi E, Pélissier R, Ploton P, Ryan CM, Saldarriaga JG, Vieilledent G (2014) Improved allometric models to estimate the aboveground biomass of tropical trees. Glob Change Biol 20:3177–3190
Chave J, Davies SJ, Phillips OL, Lewis SL, Sist P, Schepaschenko D, Armston J, Baker TR, Coomes D, Disney M, Duncanson L, Hérault B, Labrière N, Meyer V, Réjou-Méchain M, Scipal K, Saatchi S (2019) Ground data are essential for biomass remote sensing missions. Surv Geophys 40:863–880
Clark DB, Clark DA (1990) Distribution and Effects on Tree Growth of Lianas and Woody Hemiepiphytes in a Costa Rican Tropical Wet Forest. J Trop Ecol 6:321–331
Contreras MP, Bernal-Escobar M, Kullberg A, Osorio-Ospina JC, Herrera Y, Aragón L, Palacios C, Fortier RP (2024) Composición y estructura de una parcela permanente en la Sierra Nevada de Santa Marta. Jardín Botánico de Cartagena "Guillermo Piñeres". [dataset]. V1.1. https://doi.org/10.15472/ftwol2
Dawkins HC, Field DRB (1978) A long-term surveillance system for British woodland vegetation. Commenwealth Forestry Institute, University of Oxford, Oxford
ForestPlots.net, Blundo C, Carilla J, Grau R, Malizia A, Malizia L, et al (2021) Taking the pulse of Earth’s tropical forests using networks of highly distributed plots. Biol Conserv 260:1–27
Laurance WF, Lovejoy TE, Vasconcelos HL, Bruna EM, Didham RK, Stouffer PC, Gascon C, Bierregaard RO, Laurance SG, Sampaio E (2002) Ecosystem decay of Amazonian forest fragments: A 22-year investigation. Conserv Biol 16:605–618
Lopez-Gonzalez G, Lewis SL, Burkitt M, Phillips OL (2011) ForestPlots.net: a web application and research tool to manage and analyse tropical forest plot data. J Veg Sci 22:610–613
Phillips O, Baker T, Feldpausch T, Brienen R (2018) RAINFOR field manual for plot establishment and remeasurement
Réjou-Méchain M, Tanguy A, Piponiot C, Chave J, Hérault B (2017) Biomass: an R package for estimating above-ground biomass and its uncertainty in tropical forests. Methods Ecol Evol 8:1163–1167
Van der Hammen T, Ruiz PM (eds) (1984) La Sierra Nevada de Santa Marta (Colombia) Transecto Buritaca-La Cumbre. J. Cramer, Berlin-Stuttgart
Zanne AE, Lopez-Gonzalez G, Coomes DA, Ilic J, Jansen S, Lewis SL, Miller RB, Swenson NG, Wiemann MC, Chave J (2009) Data from: towards a worldwide wood economics spectrum. Dryad, Dataset
Acknowledgements
We thank Fundación ProAves for the use of their forest and their support during the field campaign. We also thank Marcela Yacomelo, Rubén Sanguino Arciniegas, Christian Vasquez, Erandi Garcia, Olivia Lockyear, and Luis Gabriel Mosquera for their assistance in the field.
Code availability
Not applicable.
Funding
Funding was provided by a Botany in Action Fellowship from the Phipps Conservatory and Botanical Gardens, a Tinker Field Research Grant from the University of Miami Institute of the Americas, the Savage, Evoy, and Kushlan funds from the University of Miami Department of Biology, and by the Colombian Bank of the Republic Foundation “Fundación para la promoción de la investigación y la tecnología” agreement 202316.
Author information
Authors and Affiliations
Contributions
RPF, MBE, and ATK conceptualized the study. RPF, MBE, ATK, MPC, JCOO, YHM, LA, and CP conducted field work. MPC and RPF curated, organized, and uploaded data to the repository. RPF led the production of the manuscript, and all the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Some species in the dataset are rare, endangered, and/or endemic to the Sierra Nevada de Santa Marta. The authors declare that they obtained the approval of Fundación ProAves for conducting the study in their private reserve, Reserva El Dorado.
Plants were collected under the joint permit endorsed by the Colombian National Environmental Authority by resolution 00571 of May 18, 2017, to the Jardín Botánico de Cartagena “Guillermo Piñeres,” “Permiso Marco de Recolección de Especímenes de Especies Silvestres de la Diversidad Biológica con Fines de Investigación Científica No Comercial.” All specimens are stored in the herbarium at the Jardín Botánico de Cartagena “Guillermo Piñeres” (JBGP).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Handling editor: Véronique Lesage.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fortier, R.P., Bernal-Escobar, M., Kullberg, A.T. et al. Diversity, composition, and structure of a 1-hectare tree plot in the cloud forest of the Sierra Nevada de Santa Marta, Colombia. Annals of Forest Science 81, 24 (2024). https://doi.org/10.1186/s13595-024-01239-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13595-024-01239-6