
Abstract
Soft-tissue preservation in molluscs is generally rare, particularly in bivalves and gastropods. Here, we report a three-dimensionally preserved specimen of the limid Acesta clypeiformis from the Cenomanian of France that shows preservation of organic structures of the adductor muscles. Examination under UV-light revealed likely phosphatisation of organic remains, which was corroborated by EDX-analyses. We suggest that the parts of the adductor muscles that are very close to the attachment are particularly resistant to decay and thus may be preserved even under taphonomic conditions usually not favouring soft-tissue fossilisation.
Similar content being viewed by others


Introduction
Bivalves have a long evolutionary history that dates back to the Early Cambrian (Geyer & Streng, 1998). Their diversity, abundance and metabolic activity steadily increased (Payne et al., 2014) and they have been a dominant taxon of benthic marine ecosystems since the Triassic (Friesenbichler et al., 2021). The autecology of extinct bivalve taxa is usually well-understood because of a close correlation between shell morphology and mode of life. However, preservation of soft parts is rare (e.g., Klug et al., 2005). Here, we report the preservation of soft tissue from the adductor muscle of the limid bivalve Acesta clypeiformis from the Cenomanian of France.
The adductores are usually differentiated in two parts that correspond to two different functions: (1) quickly contracting to close the valves under threat (quick muscle; fast, strong and short) and (2) keeping the valves closed firmly (catch muscle; slow, continuous, e.g., Bowden, 1958; Millman, 1967; Chantler, 2006; Simone, 2019; Eggermont et al., 2020; Castro-Claros et al., 2021). Accordingly, these muscle fibres can be differentiated functionally and morphologically into striated muscular fibres and smooth fibres. Their attachment to the shells is usually very strong (Castro-Claros et al., 2021), reflecting their importance in protecting the animal. Because of the required strength of this connection, the basal part of the adductors might become fossilized more easily than other soft tissues.
Fossilisation of mollusc soft parts is rare. Soft tissue-preservation is reasonably common in cephalopods (e.g., Kear et al., 1995; Klug et al., 2021a; Klug et al., 2021b; Hoffmann et al., 2021), it is very rare in gastropods (Sutton et al., 2006) and quite rare in bivalves (a list of references is given in Table 1).
Here we report soft tissue remains of a Late Cretaceous bivalve from France assigned to Acesta clypeiformis. We include analyses of its chemical composition and the significance of this discovery.
Materials and methods
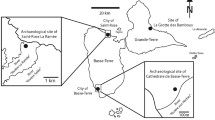
Only one specimen of the limid bivalve Acesta clypeiformis with preserved soft-tissue is available. The specimen was found by L.H. at the Falaises des Vaches Noires. These cliffs are 110 m high and extend over 5 km along the coast of the Calvados Department (Normandy, France), between Villers-sur-Mer and Houlgate. Research on the Falaises des Vaches Noires began in 1776 with the work of the monk Jean-Francois Dicquemare (1775). Since then, it became a classic fossil locality of palaeontology (e.g., Brignon, 2015, 2017). The sediments composing these cliffs range in age from the late Callovian (Jurassic) to the Late Oxfordian. Oxfordian sediments crop out on the natural cut between the lowest level of the beach and the top of the lower cliff (Dugué et al., 1998). Set back further and more difficult to access, because of intense erosion, the early Cenomanian chalk (occasionally also late Aptian to late Albian) occurs at a height of 30–40 m above the beach (Beaugrand, 1884). Periodically, natural landslides are caused by groundwater emerging from the Cenomanian sediments above the Oxfordian marls (Costa et al., 2006; Duperret et al., 2005). These exposed parts of the section deliver scree, which slides down the slopes and ravines, sometimes all the way to the beach. The name “Vaches Noires” derives from large Cenomanian blocks that fell on the beach and were subsequently covered by algae and shells, reminiscent of a herd of cows (Fig. 1). The specimen of Acesta clypeiformis with fossilized soft-tissue remains was collected by L.H. from one of these blocks of glauconitic chalk from the Cenomanian. It is now stored with the number PIMUZ 37855 at the Paläontologisches Institut und Museum of the Universiät Zürich.

Falaises des Vaches Noires, France, with the early Cenomanian blocks on the beach, where the here described Acesta clypeiformis (d’Orbigny, 1847) was found (photo by LH)
The remains of the right adductor muscle were examined by Andres Käch (Zürich) using a JEOL JSM-6010 tungsten cathode SEM with EDX v. 3.01, using a spot size of 50 μm without and with a high vacuum.
R.R. took photos of the bivalve under white artificial light and a UVA-handlamp (Hönle UV technology) with a Nikon D3X with a Nikon AF-S Micro Nikkor 105 mm 1:2.8 objective and a UV (UV-Filter MC Lotus from Kaiser Fototechnik) and polarising filters (Nikon Circular Polarizing Filter II). The colour of the UV-photos was corrected in Adobe PhotoShop 2021.
Results
Description
Specimen PIMUZ 37855 (Fig. 2) is slightly deformed and is preserved with the external (calcitic) shell layer of both valves, which flaked off over a larger part of the right valve and, to a lesser extent, also over parts of the left valve. The internal filling consists of glauconitic marl. The specimen has been identified as Acesta clypeiformis (d’Orbigny, 1847), a widespread species of the family Limidae, figured, e.g., by Woods (1904, p. 26–27, Fig. 5, who included the species in the genus Lima). The articulated valves are 82 mm high, 73 mm long and 34 mm wide, although the latter value is altered by compaction. The shell of both valves flaked off around the attachment of the adductor, thereby revealing phosphatized remains of their bases, which is better visible on the right valve. There, the brown and beige remains cover an irregular triangular surface measuring 13.1 mm in length and 11.5 mm in height. The brown and beige surface shows a reticulate pattern of polygonal sectors between 1.5 and 2.5 mm wide, surrounded by brown lines of 0.3–0.4 mm width. These lines become lighter in colour postero-ventrally and show a faint striation with the same orientation (running postero-ventrally). The lines are referred to as brown parts, while the inside of these polygonal fields are referred to as beige parts. Within the beige parts, comma-shaped patches are distributed irregularly. They are about 0.5 mm long and between 0.1 and 0.2 mm wide. The reticulate pattern corresponds to the bundles of muscle fibres that made up the muscle.

Acesta clypeiformis (d’Orbigny, 1847) with adductor muscle scars retaining phosphatized muscle remains. Cenomanian, Falaises des Vaches Noires, France. B taken under UV-light, A, B adductor muscle of the right valve (marked with a rectangle in D). C–F Photos of the entire specimen. C Anterior; D right; E posterior; F left view. G Adductor muscle of the left valve (marked with a rectangle in F)
Chemical analyses of the adductor muscle remains
We ran several EDX analyses, four of which are presented here (Fig. 3, Table 2.). Three of the analyses were made in the brown parts and two of these three were made in the beige parts. The latter two analyses revealed similar amounts of oxygen (42–43 wt.%), calcium (32–35 wt.%), phosphorous (12–13 wt.%), carbon (4–7 wt.%) and fluorine (close to 4 wt.%). The analysis from the brown part revealed higher percentages of oxygen and carbon, but relatively less calcium and phosphorous. The analysis from the sedimentary matrix of the internal mould indicates a high content of calcium, carbon and oxygen, corroborating a CaCO3 composition.

Element analyses of the adductor remains and the sedimentary filling of the bivalve shown in Fig. 2. For the weight percent values, see Table 2. a Detail of the bivalve showing the part that was measured (black rectangle). b Magnified detail taken by SEM, showing the spots, where the EDX-measurements were taken. c and d EDX measurements from the beige parts. e EDX measurement from the brown parts. f EDX measurement from the internal mould (glauconitic marl)
Discussion
What is preserved?
Limids are monomyarian (Cox & Hertlein, 1969); their single adductor is homologous with the posterior adductor of dimyarian pteriomorphs. On both valves, only the posteroventral parts of the proximal part of the adductor muscle is preserved. We assume that not the entire muscle is preserved but only the part very close to the shell attachment.
How can its preservation be explained?
As demonstrated by Castro-Claros et al. (2021), Ca2+-ions play an important role in the attachment of muscles in bivalves. This suggests that the richness in calcium carbonate made the part of the muscle that was closest to the shell more resistant to decay and thus increased the likelihood of becoming phosphatized.
Acesta is a byssate limid bivalve. Modern representatives of this genus live in cold water or at great depths (Cox & Hertlein, 1969). Low temperatures would slow down disintegration of soft-tissues. However, Merles (2011) suggested that the water was rather warm during the Cenomanian in that region (see also O’Brien et al., 2017).
Phosphatisation of soft-tissues is known to occur in the sediment near the redox boundary (Allison, 1988a, 1988b; Briggs & Wilby, 1996; Briggs et al., 1993), often in combination with bacterial activity. In contrast to most other bivalves with fossilized soft-tissues (see Table 1), Acesta is not infaunal. Limids are usually attached to the substratum by the byssus, but they can release the byssus for swimming when attacked by a predator (Stanley, 1970).
Accordingly, we hypothesize rapid burial, which inhibited the decay of soft-tissues and brought the remains to the redox-boundary, thus enabling phosphatisation. It is also conceivable that local conditions within the closed shell were more important than the position of the redox boundary in the sediment and favoured phosphatisation. In addition, due to the proximity to the muscle insertion, we suggest that the resistance to decay of this part of the muscle is linked with the presence of raised levels of Ca2+-ions(Castro-Claros et al., 2021).
Conclusions
We describe a Late Cretaceous fossil of a byssate epifaunal bivalve, which preserves those parts of the posterior adductor muscle that are the closest to the shell. We suggest that the combination of the proximal muscle with its abundant collagen fibres and the shell carbonate at the muscle insertion inhibited decomposition. Rapid burial likely stopped decay and enabled phosphatisation of these organic remains. This shows that under certain conditions, soft tissues may become fossilized in taphonomic contexts, where such preservation would normally not be expected.
Availability of data and materials
The single specimen is incorporated in the collections of the Palaeontological Institute and Museum of the University of Zurich (PIMUZ 37855).
References
Allison, P. A. (1988a). Konservat-Lagerstätten: Cause and classification. Paleobiology, 14, 331–344.
Allison, P. A. (1988b). Soft bodied squids from the Jurassic Oxford Clay. Lethaia, 21, 403–410.
Beaugrand, C. (1884). Le Cénomanien de Villers-sur-Mer. Bulletin De La Société Géologique De La Normandie, 9(1882), 16–18.
Bowden, J. (1958). The structure and innervation of lamellibranch muscle. International Review of Cytology, 7, 295–335.
Briggs, D. E. G., Kear, A. J., Martill, D. M., & Wilby, P. R. (1993). Phosphatization of soft-tissue in experiments and fossils. Journal of the Geological Society, London, 150, 1035–1038.
Briggs, D. E. G., & Wilby, P. R. (1996). The role of the calcium carbonate-calcium phosphate switch in the mineralization of soft-bodied fossils. Journal of the Geological Society, London, 153, 665–668.
Brignon, A. (2015). Les débuts de la paléoichthyologie en Normandie et dans le Boulonnais. Revue Française De Paléontologie, 21, 43–62.
Brignon, A. (2017). Les « ossements pétrifiés » du Jurassique des Vaches Noires dans les cabinets d’histoire naturelle du XVIIIe et du début du XIXe siècles. L’écho Des Falaises, Association Paléontologique De Villers-Sur-Mer, 21, 7–35.
Castro-Claros, J. D., Checa, A., Lucena, C., Pearson, J. R., & Salas, C. (2021). Shell-adductor muscle attachment and Ca2+ transport in the bivalves Ostrea stentina and Anomia ephippium. Acta Biomaterialia, 120, 249–262.
Chantler, P. D. (2006). Scallop adductor muscles: Structure and function. In S. E. Shumway & G. J. Parsons (Eds.), Scallops: Biology, ecology and aquaculture (pp. 229–316). Elsevier.
Chiba, T., Sato, S., & Yamada, T. (2014). Fossilized intestine casts located within closed bivalve shells: Implications for palaeoecological and sedimentological studies. Lethaia, 12, 341–352. https://doi.org/10.1111/let.12111
Costa, S., Henaff, A., & Lageat, Y. (2006). The gravel beaches of North-West France and their contribution to the dynamic of the coastal cliff-shore platform system. Zeitschrift Für Geomorphologie, 144, 194–199.
Cox, L. R. (1960). The preservation of moulds of the intestine in fossil Nuculana (Lamellibranchia) from the Lias of England. Palaeontology, 2, 262–269.
Cox, L. R., & Hertlein, L. G. (1969). Family Limidae Rafinesque, 1815. N385. In R. C. Moore (Ed.), Treatise on invertebrate paleontology. Part N. Mollusca 6. Bivalvia 1. (p. 489). Geological Society of America, Boulder, and University of Kansas Press.
Whyte, M. A., Wimbledon, W. A., & Morris, N. J. (1984). The preservation of soft-tissues in Portlandian trigoniids. Berlin: Palaeontological Association annual conference, abstracts, Fitzwilliam College, University of Cambridge.
Woods, H. (1904). A monograph of the Cretaceous Lamellibranchia of England (Vol. 2, pp. 1–7). Palaeontographical Society.
De la Beche, H. T. (1848). Anniversary address of the president. Quarterly Journal of the Geological Society of London, 4, 21–120.
De la Beche, H. T. (1849). Anniversary address of the president. Quarterly Journal of the Geological Society of London, 5, 19–116.
d’Orbigny, A. (1847). Paléontologie française—Terrains crétacé—Tome 3, Lamellibranchia (liv 114) (pp. 537–544). Arthus Bertrand.
Dicquemare, J. F. (1775). Observations sur les coquilles fossiles, & particulièrement sur les cornes d’ Ammon. Observations Sur La Physique, Sur L’histoire Naturelle Et Sur Les Arts, 5, 435–439.
Dugué, O., Fily, G., & Rioult, M. (1998). Le Jurassique des côtes du Calvados. Biostratigraphie, sédimentologie, paléoécologie, paléogéographie et stratigraphie séquentielle. Bulletin Trimestriel De La Société Géologique De Normandie Et Des Amis Du Muséum Du Havre, 85, 1–132.
Duperret, A., Taibi, S., Mortimore, R. N., & Daigneault, M. (2005). Effect of groundwater and sea weathering cycles on the strength of chalk rock from unstable coastal cliffs of NW France. Engineering Geology, 78, 321–343.
Eggermont, M., Cornillie, P., Dierick, M., Adriaens, D., Nevejan, N., Bossier, P., Van den Broeck, W., Sorgeloos, P., Defoirdt, T., & Declercq, A. M. (2020). The blue mussel inside: 3D visualization and description of the vascular-related anatomy of Mytilus edulis to unravel hemolymph extraction. Scientific Reports, 10, 6773. https://doi.org/10.1038/s41598-020-62933-9
Friesenbichler, E., Hautmann, M., & Bucher, H. (2021). The main stage of recovery after the end-Permian mass extinction: Taxonomic rediversification and ecologic reorganization of marine level-bottom communities during the Middle Triassic. PeerJ, 9, e11654.
Gavey, G. E. (1853). On the railway cuttings at the Mickleton Tunnel and at Aston Magna, Gloucestershire. Quarternary Journal of the Geological Society of London, 9, 29–37.
Geyer, G., & Streng, M. (1998). Middle Cambrian pelecypods from the Anti-Atlas, Morocco. Revista Española De Paleontologia, No Extr. Homenaje Al Prof. Gonzalo Vidal, 5, 83–96.
Harper, E. M., & Todd, J. A. (1996). Preservation of the adductor muscle of an Upper Jurassic oyster. Paläontologische Zeitschrift, 69, 55–59.
Hoffmann, R., Morón-Alfonso, D., Klug, C., & Tanabe, K. (2021). Ammonoid soft-tissue remains revealed by computed-tomography. Swiss Journal of Palaeontology, 140, 14.
Kear, A. J., Briggs, D. E. G., & Donovan, D. T. (1995). Decay and fossilisation of non-mineralised tissue in coleoid cephalopods. Palaeontology, 38, 105–131.
Kiel, S., Götz, S., Pascual-Cebrian, E., & Hennhöfer, D. K. (2012). Fossilized digestive systems in 23 million-year-old wood-boring bivalves. Journal of Molluscan Studies, 78, 349–356.
Klug, C., Hagdorn, H., & Montenari, M. (2005). Phosphatised soft-tissue in Triassic bivalves from Germany. Palaeontology, 48, 833–852.
Klug, C., Pohle, A., Roth, R., Hoffmann, R., Wani, R., & Tajika, A. (2021a). Preservation of nautilid soft parts inside and outside the conch interpreted as central nervous system, eyes, and renal concrements from the Lebanese Cenomanian. Swiss Journal of Palaeontology, 140, 11.
Klug, C., Schweigert, G., Tischlinger, H., & Pochmann, H. (2021). Failed prey or peculiar necrolysis? Isolated ammonite soft body from the Late Jurassic of Eichstätt (Germany) with complete digestive tract and male reproductive organs. Swiss Journal of Palaeontology, 140, 1–15. https://doi.org/10.1186/s13358-020-00215-7
Lopes De Simone, L. R., & Mezzalira, S. (1993). Vestígios de partes moles em um bivalve fóssil (Unionida, Mycetopodidae) do Grupo Baro (Cretáceo Superior), São Paulo, Brasil. Anais Da Academia Brasileira De Ciěncias, 65, 155–159.
Mantell, G. A. (1843). Notice on the fossilized remains of the soft parts of Mollusca. Proceedings of the Geological Society of London, 4, 35–36.
Mehl, J., & Rehfeld-Kiefer, U. (1992). Sipho-Erhaltung bei Pholadomya fidicula (Pelecypoda) aus dem Mittleren Jura (Bajocium), ein Vergleich mit der rezenten Pholadomya candida. Senckenbergiana Lethaea, 71, 367–375.
Merles, B. (2011). Les falaises des Vaches Noires de Cuvier au Paléospace (p. 129). Éditions des Falaises.
Millman, B. M. (1967). Mechanisms of contraction in molluscan muscle. American Zoologist, 7, 583–591.
O’Brien, C. L., Robinson, S. A., Pancost, R. D., Sinninghe Damsté, J. S., Schouten, S., Lunt, D. J., Alsenz, H., Bornemann, A., Bottini, S., Brassell, S. C., Farnsworth, A., Forster, A., Huber, B. T., Inglis, G. N., Jenkyns, H. C., Linnert, C., Littler, K., Markwick, P., & Wrobel, N. E. (2017). Cretaceous sea-surface temperature evolution: Constraints from TEX86 and planktonic foraminiferal oxygen isotopes. Earth-Science Reviews, 172, 224–247.
Payne, J. L., Heim, N. A., Knope, M. L., & McClain, C. R. (2014). Metabolic dominance of bivalves predates brachiopod diversity decline by more than 150 million years. Proceedings of the Royal Society B, 281, 1–9. https://doi.org/10.1098/rspb.2013.3122
Simone, L. R. L. (2019). Modifications in adductor muscles in bivalves. Malacopedia, 2, 1–12.
Spamer, E. E., & Bogan, A. E. (1989). Recovery of the Etheldred Benett collection of fossils mostly from Jurassic-Cretaceous strata of Wiltshire, England, analysis of the taxonomic nomenclature of Benett (1831), and notes and figures of type specimens contained in the collection. Proceedings of the Academy of Natural Sciences of Philadelphia, 141, 115–180.
Stanley, S. M. (1970). Relation of shell form to life habits of the Bivalvia (Mollusca). Geological Society of America, 125, 296.
Wilby, P. R., & Whyte, M. A. (1995). Phosphatized soft-tissues in bivalves from the Portland Roach of Dorset (Upper Jurassic). Geological Magazine, 132, 117–120.
Stewart, J. D. (1990). Preliminary account of halecostome-inoceramid commensalism in the Upper Cretaceous of Kansas. In A. J. Boucot (Ed.), Evolutionary paleobiology of behaviour and coevolution (pp. 51–60). Elsevier.
Sutton, M. D., Briggs, D. E. G., Siveter, D. J., & Siveter, D. J. (2006). Fossilized soft tissues in a Silurian platyceratid gastropod. Proceedings of the Royal Society B, 273, 1039–1044. https://doi.org/10.1098/rspb.2005.3403
Todd, J. A. & Palmer, T. J. (2002). The Jurassic bivalve genus Placunopsis: new evidence on anatomy and affinities. Palaeontology, 45, 487–510.
Torrens, H. S., Benamy, E., Daeschler, E. B., Spamer, E. E., & Bogan, A. E. (2000). Etheldred Benett of Wiltshire, England, the first lady geologist—Her fossil collection in the Academy of Natural Sciences of Philadelphia, and the rediscovery of “lost” specimens of Jurassic Trigoniidae (Mollusca: Bivalvia) with their soft anatomy preserved. Proceedings of the Academy of Natural Sciences of Philadelphia, 150, 59–123.
Whyte, M. A. (1992). Phosphate gill supports in living and fossil bivalves. In S. Suga & H. Nakahava (Eds.), Mechanisms and phylogeny of mineralisation in biological systems (pp. 427–431). Springer.
Acknowledgements
We thank the two anonymous reviewers for their constructive remarks.
Funding
This study was supported by SNF grant 200021-162402/1 (to MH).
Author information
Authors and Affiliations
Contributions
LH found the specimen and had the idea to publish it. CK supervised the SEM-work and the chemical analyses. RR produced the photographs of the specimen using white und UV-light. CK, LH, and RR produced the figures. MH provided information about bivalve palaeobiology. All authors wrote parts of the text, proof read earlier and the final version and approved of it. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
We have no competing interests.
Additional information
Editorial handling: Daniel Marty
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Klug, C., Hüne, L., Roth, R. et al. Phosphatized adductor muscle remains in a Cenomanian limid bivalve from Villers-sur-Mer (France). Swiss J Palaeontol 141, 10 (2022). https://doi.org/10.1186/s13358-022-00252-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13358-022-00252-4