
Abstract
The variability of metric traits has been studied in 20 Kazakhstan populations of three Nitraria species (N. schoberi L., N. sibirica Pall., and N. komarovii Iljin & Lava ex Bobrov). According to the results, N. sibirica differs from N. schoberi and N. komarovii in small leaves, fruits, and seeds, as well as in the structural features of inflorescences and flowers. N. sibirica is characterized by a greater number of flowers per inflorescence (25–66 on average) than N. schoberi and N. komarovii (14–28 on average). Additionally, N. sibirica differs from N. schoberi by narrower (1.3×) petals and smaller anthers (1.46× in length and 1.2× in width) and pistils (1.25× in length and 1.44× in width). N. komarovii differs from N. schoberi by narrower leaves (1.7×). N. komarovii is characterized by relatively large (especially in width) size of anthers that, together with smaller petals, makes it possible to distinguish this species during flowering. Flowers of N. komarovii are bright yellow, while those in N. schoberi and N. sibirica are white and light purple, respectively. N. komarovii fruits are orange, pale red, or bright red. Fruits of N. schoberi and N. sibirica are dark burgundy and black, respectively. At the intraspecific level, most of the studied metric traits (except for the habitus of N. sibirica plants) are stable and do not depend on the 16 climatic parameters of plant habitats. At the same time, a number of morphological features have been revealed in N. sibirica plants from the Ili Depression. In these populations, N. sibirica plants form shrubs up to 1.8 m in height with a large (up to 90) number of flowers per inflorescence, large petals (3.5 mm in length and 2.2 mm in width on average), small (~0.6 mm) anthers, small (5–5.5 mm in length) fruits, and small (~4 mm in length) narrow ovate stones. These features indicate the ecological–geographical differentiation of N. sibirica under the extra-arid conditions of the stony desert of the depression and evidence a separate taxonomic rank of these populations.
Similar content being viewed by others

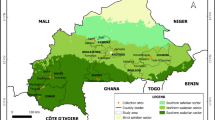

INTRODUCTION
The genus Nitraria L. consists of ~10 species of halophytic shrubs from steppe, semidesert and desert regions of Asia, North Africa, Southeastern Europe (Romania), and Australia. Despite the smallness of the genus, scientists have no clear ideas about the number, chorology, or genesis of Nitraria species. Insignificant differences in the metrical traits of vegetative and generative organs serve as a reason for some authors to recognize extreme variants as independent species, while others consider them intraspecific forms or ecological races (Ilyin, 1944; Bobrov, 1946, 1965; Lukanenkova, 1964; Petrov, 1972; Vasilyeva, 1974).
Only two Nitraria species, N. schoberi L. and N. sibirica Pall, are mentioned in the special literature for the studied region (Pavlov, 1963; Abdulina, 1998; Baitenov, 1999, 2001). Petrov (1972) describes Nitraria roborowskii Kom. from Central and Middle Asia. According to the opinion of this author, this species is rather widespread and grows together with N. schoberi. In our earlier study of the Balkhash-Alakol depression, we first discovered N. komarovii Iljin & Lava ex Bobrov on the territory of Kazakhstan (Tomoshevich et al., 2019).
N. komarovii is considered a relatively young modification of N. schoberi under halophilic conditions of the Caspian littoral (Ilyin, 1958; Bobrov, 1965). However, M.P. Petrov (1972) states that similar processes occur in other ecologically similar habitats located on the coasts of large saline Asiatic lakes. In connection with this, in his opinion, narrow-leaved forms identified as N. komarovii should be considered as a form of N. schoberi f. komarovii (Iljin & Lava) Petrov instead.
According to E.G. Bobrov (1965), N. roborowskii Kom. described by V.L. Komarov (1908) and differing from N. schoberi by greater leaves, inflorescences, and drupes, is an ecological form of N. sibirica (N. sibirica f. majus). He suggested considering the increased size of vegetative and generative organs of this plant as a gigantism under favorable ecological conditions (moist clay-alkaline soil of oases, lake shores, and dry sandy reedbeds).
To understand the processes occurring in natural populations and to analyze taxonomic diversity, one should study the variability of organisms, because the number of species can be increased via their geographic variability (Gall and Popov, 1998). Morphological variability can result from unheritable changes of traits caused by the variability of environmental conditions or from the process of evolutionary transformation of living things (Lukin, 1940; Mayr, 1974).
The purpose of the study was to investigate the variability of morphological traits in plants from the genus Nitraria to identify the environmental component of this variability and clarify their taxonomic rank.
MATERIALS AND METHODS
Specimens of N. sibirica (nine habitats), N. schoberi (10 habitats), and N. komarovii were collected in 2012–2017 during field trips arranged on the territory of the Republic of Kazakhstan. For each population of plants, 25–30 herbarium sheets were collected, as well as specimens of flowers, fruits, and seeds, which were put into paper bags, marked, and delivered to the Novosibirsk laboratory for the morphometric analysis.
All samples collected during field trips were deposited in the NSC collection of the Central Siberian botanical garden of the Siberian branch of the Russian Academy of Sciences (Novosibirsk, Russia); they are also available in the digital herbarium of this botanical garden
A comparative morphological analysis was carried out by the method used for studying the intraspecific variability of woody plants and developed by S.A. Mamaev (1972, 1975). Leaves, flowers, and fruits were measured using a Carl Zeiss Stereo Discovery V12 microscope equipped with a high-resolution Axio Cam digital camera. The images were obtained, treated, and analyzed using AxioVision v. 4.8 software. Morphological measurements of vegetative and generative plant parts were performed in 30 replications using a SIAMS Photolab program and a SIAMS MesoPlant module.
The analysis of climatic characteristics of Nitraria habitats included 16 parameters: average monthly temperatures in May, July, and January; annual air temperature amplitude; Conrad’s index; frost-free period; average maximum and minimum of annual temperatures; number of days per year with T > 5°C; number of days per year with T > 10°C; sum of air temperatures for the period with T > 5°C; sum of air temperatures for the period with T > 10°C; annual precipitation, mm; precipitation–evaporation ratio; precipitations during April–October period; and air saturation insufficiency.
The similarity or difference between the objects (habitats or populations) was determined by a cluster analysis (Ward’s method) using Euclidean distance as a measure of similarity.
RESULTS
Morphological Variability of N. schoberi
N. schoberi typically represents loose sprawling shrubs with arcuate stems and sparse large thorns. In most populations, plant height varied within 60–80 cm. Specimens with a greater height (up to 1–1.2 m) were reported for the Koktal and Basshi populations. The length of N. schoberi leaf in the studied populations varied from 2 to 4 cm, with an average value of 2.5–3 cm (Table 2). The smallest leaves were observed in N. schoberi plants belonging to the Aidarly, Basshi, and Koktal populations; their average length was ~2 cm. The leaf coefficient values varied from 0.14 to 0.26; the widest leaf part was localized above its middle. Thus, N. schoberi is characterized by oblong–spatulate and oblanceolate leaves (Fig. 1). Leaves of plants from the Aidarly population were linear–spatulate.

Leaf shape in N. schoberi: (a) Aidarly and (b) Balkhash.
The number of flowers in N. schoberi inflorescences varied from 10 to 32; an increased value of this parameter (25 on average) was observed in southern populations, such as Saryozek, Basshi, Koktal, and Charyn (Figs. 2, 3). The internode length in N. schoberi inflorecences varied within 3.5–4.5 mm. The longest internodes (up to 10.7 mm) were observed in the Basshi population.

Features of inflorescence structure in N. schoberi: (a) Balkhash, (b) Basshi, and (c) Lepsy.

Grouping of N. schoberi populations according to the number of flowers per inflorescence.
Petals in N. schoberi were ovate or (rarely) diamond-shaped, with short claws (Ak-Lama et al., 2017) (Fig. 4). The petal length varied from 3 to 4.5 mm (Table 3).

Variability of petals in N. schoberi: (a) Charyn, (b) Basshi, and (c) Karatal.
Fruits were dark-red or (rarely) black (Alakol, Lepsy, and Raz’ezd_47), but could also be bright burgundy (Saryozek). The juice had a pale burgundy or pale pink color. The juice color significantly correlated with the peel color. Fruits were widely oval and usually did not exceed 1 cm in length and 7–8 mm in diameter. The weight of 100 stone varied from 220 to 440 g. The stone length in N. schoberi varied between 7 and 10 mm, being 8–9.5 mm on average. The stone width was 4.5–5.5 mm. In all studied N. schoberi populations, stones were ovate (Fig. 5).

Variability of N. schoberi seeds: (a) Balkhash, (b) Lepsy, and (c) Saryozek.
Morphological Variability of N. komarovii
The height and diameter of N. komarovii plants were 0.5–1 and 1.5–3 m, respectively. Shrubs were strongly branched from the base, shoots with relatively sparse thorns were slightly bent and elevated in the central part. The length and width of leaves were 2.5–3 and 0.3 cm, respectively. The leaf coefficient was 0.08–0.13.
The number of flowers per inflorescence varied from 17 to 30 (25 on average). The internode length in inflorecences varied within 4–7 mm (5.5 mm on average). Petals were ovate; their average length and width were 3 and 2 mm, respectively.
Fruits were orange, pale red, or bright red, with a light pink juice. The weight of 100 stone varied between 370 and 530 g. Stones had a prolate–conical form with a typical tapered top; their length and width were 9–11 and 5–7 mm, respectively.
Morphological Variability of N. sibirica
N. sibirica had a variable habitus and was characterized by very thorny semibent shoots elevated in the central part of a shrub.
The leaf length varied slightly, being 1.0–1.5 cm on average. The smallest leaves were observed in the northern Kainar and Kurgan populations (Table 2). The leaf coefficient varied within 0.2–0.33; the widest part of the leaf was located above its middle, so N. sibirica leaves can be considered oblanceolate.
The maximum number of flowers per inflorescence (up to 90) was observed in N. sibirica populations localized in the Ili depression (Koktal and Basshi; Fig. 6), whereas the typical average value of this parameter in this species was 25–30.

Structure of the N. sibirica inflorescence: (a, b) Koktal, (c) Kurti, and (d) Matai.
Petals were acuminate–elliptic; diamond-shaped and ovate petals were observed in some plants of the Basshi, Karatal, and Koktal populations (Fig. 7). The petal length varied from 2.5 to 4 mm. In three abovementioned populations, petals had a greater size (3.4–3.5 mm on average; Table 3), while in the Balkhash and Taskarasu populations, the average value of this parameter was ~3.1 mm.

Variability of petals in N. sibirica: (a) Balkhash, (b) Kurti, (c) Taskarasu, and (d) Karatal.
Ripe fruits of N. sibirica were always black; in the ripening period, they had a bright red color. The juice of ripe fruits was dark blue; the only exclusions were the Karatal and Kurti populations, in which we observed blackish green and light black juice of fruits, respectively. N. sibirica fruits were rounded (~5 mm in diameter). Small stone (4–6 mm) were observed in the Basshi and Karatal populations, while the largest ones (9 mm) were registered in the Matai population. The weight of 100 stone in the Matai population was 200–270 g; in the Basshi population, it was 50–80 g. In other populations, the value of this parameter varied within 100–200 g.
Small and narrow ovate stones with a narrow pointed tip were typical for the Basshi, Koktal, Kurti, and Taskarasu populations of N. sibirica. Greater ovate stones were observed in the Balkhash, Karatal, and Matai populations (Fig. 8).

Individual variability of N. sibirica seeds: (a) Balkhash, (b) Karatal, (c) Matai, (d) Basshi, (e) Koktal, and (f) Kurti.
DISCUSSION
An analysis of the data confirmed morphological differences between N. sibirica, N. schoberi, and N. komarovii (Banaev et al., 2017). N. sibirica was put into a separate cluster (Fig. 9) as a small-leaved and small-fruited species. In addition, N. sibirica differs from N. schoberi and N. komarovii in some details of the inflorescence structure. N. sibirica is characterized by a greater number of flowers per inflorescence (25–66 on average) compared to N. schoberi and N. komarovii (14–28 on average). Additionally, N. sibirica differs from N. schoberi by narrower (1.3-fold) petals, as well as smaller anthers (1.46-fold in length and 1.2-fold in width) and pistils (1.25-fold in length and 1.44-fold in width).

Population similarity dendrogram for Nitraria species.
N. komarovii differs from N. schoberi by narrower leaves (1.7-fold); the leaf width in N. schoberi is ~5 mm on average, while that in N. komarovii is equal to 3 mm (Fig. 10). Leaves of both N. schoberi and N. sibirica are oblanceolate or oblong–spatulate with a blunt or slightly pointed apex. N. komarovii leaves are characterized by linear or sometimes linear–spatulate leaves with a sharply pointed, sometimes weakly pronounced, apex.

Leaf variability in Nitraria species: (a) N. komarovii, (b) N. shoberi, and (c) N. sibirica.
N. komarovii has a smaller length of the stamen (<3.5 mm) and stamen filament (<2 mm); at the same time, the length of the stamen, stamen filament, and anther in N. schoberi and N. sibirica reaches 4, 3, and almost 2 mm, respectively. At the same time, the anthers of N. komarovii are relatively large (especially in width). This phenomenon, together with smaller petals, makes it possible to distinguish this species during flowering. Flowers of N. komarovii look bright yellow, while those of N. schoberi and N. sibirica are white and light purple, respectively. N. schoberi is characterized by the largest pistils (2.5–4.7 mm in length and 1.3–2.3 mm in width). The pistils of N. sibirica and N. komarovii flowers rarely exceed 3.5 mm in length and 1.8 mm in width.
Analyzing the connection between metric traits of plants and ecological–geographical parameters of their habitats, one should mention a high stability of such traits that agrees with our earlier observations of N. sibirica populations from Siberia (Banaev et al., 2017). In the case of N. shoberi, a weak correlation with climatic factors was observed only for leaf parameters. Smaller leaves were observed in N. schoberi plants from the Aidarly, Basshi, and Koktal populations, the habitats of which are characterized by a low precipitation–evaporation ratio (0.15–0.17), a high sum of positive air temperatures (for T > 5°C it exceeds 3800, for T > 10°C it exceeds 3500), and highly insufficient air saturation (8.4–9.7). Metric traits of N. schoberi vegetative organs are also influenced by microconditions of their habitats, mainly by the groundwater availability. The Alakol population is located in the close vicinity of Lake Alakol. In Karatal and Charyn populations, N. schoberi plants grow on river terraces. Saryozek and Raz’ezd_47 populations are located on territories characterized by a high groundwater level or waterway presence. At the same time, the Aydarly population is located on the Bozoi plateau, at the roadside, where only temporary spring underflooding with meltwater occurs. Basshi and Koktal populations are located at an altitude of >1000 m above sea level, in a stony desert with over-arid soils similar to the soils of Mongolian hamadas (Evstifeev and Rachkovskaya, 1976; Nasyrov and Sokolov, 1998).
However, the effect of habitat conditions on the metric traits of vegetative and generative organs of N. schoberi is insignificant. These traits are also stable over the species range. For example, in Western Siberian populations of N. schoberi, the leaf length is comparable or sometimes slightly smaller (~2 cm) than those in Kazakh populations (Banaev et al., 2017). At the same time, sums of positive air temperatures (T > 5°C and T > 10°C) at the eastern border of the species range are significantly (by 1400°C) lower than those in Kazakhstan.
Since the Aral–Caspian basin is considered the center of origin of N. schoberi, from which the species spread to the south and east into Central Asia and Western Siberia (Bobrov, 1946), one could expect an appearance of extreme morphotypes in the island populations located at the border of the species range (Kulunda steppe). However, according to Komarov (1908), “the variability of N. schoberi is great, but poorly oriented” (p. 165) in the geographic sense. This is explained by the fact that Nitraria species are confined to local intrazonal habitats and can survive only under a combination of necessary soil and ground parameters, such as soil moisture and salinity (Khudyaev and Banaev, 2012). Both temperature and air humidity are not the main limiting factors for N. schoberi and N. sibirica. In connection with this, the clinal variability, which is inherent in many species with continuous ranges (Mayr, 1974), such as boreal woody plant species, is not typical for Nitraria species (Banaev, 1997a, 1997b; 2009a, 2009b; Banaev and Shemberg, 2000; Banaev and Adelshin, 2009).
Climatic parameters of habitats influence only on the habitus of N. sibirica plants (Figs. 11, 12), as we have shown previously for Siberian populations of this species (Banaev et al., 2017). In Altai and Tyva republics, the height of N. sibirica plants does not exceed 20 cm, while the plant height in the Khakassia and Kulunda steppe populations is 50 cm on average. Similar bushes (about 50 cm in height) were observed in the N. sibirica populations of northern Kazakhstan (Kainar and Kurgan). Generative individuals from the Balkhash, Matai, and Karatal populations have a height of 70 cm; plant height in Taskarasu and Kurti populations reached 1.0–1.4 m. The highest N. sibirica plants were observed in the Basshi and Koktal districts (their height reached 1.8 m (Fig. 13)).

Dendrogram of similarity of N. sibirica habitats in 16 climatic parameters of temperature and humidity.

Clustering of N. sibirica populations by the shrub habitus.

Habitus of N. sibirica: (a) Kainar (up to 50 cm), (b) Matai (up to 70 cm), (c) Kurti (up to 1.4 m), and (d) Koktal (up to 1.8 m).
A number of morphological features was revealed in N. sibirica plants from the Ili depression (Basshi, Koktal, and Taskarasu populations). Plants from these populations have a greater number of flowers per inflorescence (up to 90), large petals (3.5 mm in length and 2.2 mm in width on average), small anthers (~0.6 mm), small fruits (5–5.5 mm in length), and small narrow ovate stones (~4 mm in length). In addition, N. sibirica plants from the Koktal and Taskarasu populations contain fewer phenolic components compared to those from the northern (Kurgan and Kainar) populations (Ak-Lama, 2020). The revealed features in the composition and content of phenolic compounds in N. sibirica plants from the Almaty district of Kazakhstan are confirmed by the results of a comparison of phenolic profiles of N. sibirica plants from other regions, including Siberia (Voronkova et al., 2017; 2020). The intraspecific heterogeneity of N. sibirica was also revealed by an analysis of the ITS2 region of nuclear ribosomal DNA (Polyakova et al., 2020; Banaev et al., 2020). All these facts indicate the ecological and geographic differentiation of N. sibirica populations.
CONCLUSIONS
Three species of the genus Nitraria (N. schoberi, N. sibirica, and N. komarovii) have been revealed in the Republic of Kazakhstan. N. komarovii grows on the Caspian Sea coast, as well as on coastal sands in the southeastern part of Lake Balkhash. N. komarovii reliably differs from N. schoberi by narrow linear or sometimes linear–spatulate leaves with a sharply pointed or sometimes weakly manifested apex. Additionally, N. komarovii is characterized by anthers of a greater size (especially in width), which, together with small petals, allow one to distinguish this species during flowering. Flowers of N. komarovii are bright yellow, while those in N. schoberi and N. sibirica are white and light purple, respectively. Fruits of N. komarovii are orange, pale or bright red; fruits of N. schoberi are dark burgundy, while in N. sibirica they are black.
Most morphometric traits of N. schoberi and N. sibirica are stable in different parts of their range, which indicates the low dependence of these parameters on the air temperature and humidity at their habitats. Climatic parameters influence only on the habitus of plants, especially in N. sibirica.
The intraspecific differentiation of N. sibirica under conditions of the extra arid Ili depression evidences a separate taxonomic rank of these populations.
Change history
20 October 2021
An Erratum to this paper has been published: https://doi.org/10.1134/S1995425521050188
REFERENCES
Abdulina, S.A., Spisok sosudistykh rastenii Kazakhstana (The List of Vascular Plants of Kazakhstan), Almaty, 1998.
Ak-Lama, T.A., Banaev, E.V., and Tomoshevich, M.A., Features of the flower structure of some species of the genus Nitraria L., Izuchenie, sokhranenie i ratsional’noe ispol’zovanie rastitel’nosti mira Evrazii: Materialy mezhdunar. nauch. konf. posvyashch. 85-letiyu Instituta botaniki i fitointroduktsii KN MON RK (17–19 avgusta 2017 g.) (Research, Conservation, and Rational Use of the Vegetation of the World of Eurasia: Proc. Int. Sci. Conference Dedicated to the 85th Anniversary of Institute of Botany and Phytointroduction of Science Committee of Ministry of Education and Science of Republic of Kazakhstan (August 17–19, 2017)), Almaty, 2017, pp. 98–103.
Baitenov, M.S., Flora Kazakhstana (Flora of Kazakhstan), Almaty: Gylym, 1999, vol. 1.
Baitenov, M.S., Flora Kazakhstana (Flora of Kazakhstan), Almaty: Gylym, 2001, vol. 2.
Banaev, E.V., Features of variation of some metric traits in Alnus hirsuta (Spach) Turcz. Ex Rupr. with regard to the problem of biodiversity, Sib. Ekol. Zh., 1997a, no. 1, pp. 51–58.
Banaev, E.V., Variability of quantitative traits of leaves and female catkins of Alnus hirsuta (Betulaceae), Bot. Zh., 1997b, vol. 82, no. 6, pp. 86–91.
Banaev, E.V., On the effect of climate on the morphological structure of Alnus hirsuta (Betulaceae), Russ. J. Ecol., 2009a, vol. 40, no. 1, pp. 18–23.
Banaev, E.V., Phenotypic variability of Alnus fruticosa Rupr. s. l. (Betulaceae) in Asian Russia, Rast. Mir Aziat. Ross., 2009b, no. 1, pp. 44–52.
Banaev, E.V. and Adel’shin, R.V., Structure of Alnus fruticosa Rupr. s. l. and its relationships with other taxa of subgenus Alnobetula (Ehrhart) Peterman, Contemp. Probl. Ecol., 2009, vol. 2, no. 6, pp. 601–610.
Banaev, E.V. and Shemberg, M.A., Ol’kha v Sibiri i na Dal’nem Vostoke Rossii (izmenchivost', taksonomiya, gibridizatsiya) (Alder in Siberia and the Russian Far East (Variability, Taxonomy, Hybridization)), Novosibirsk: Sib. Otd. Ross. Akad. Nauk, 2000.
Banaev, E.V., Tomoshevich, M.A., and Yamtyrov, M.B., On variation of metric and qualitative characters of Nitraria L. species in the context of ecological-climatic conditions of habitat in Siberia, Contemp. Probl. Ecol., 2017, vol. 10, no. 6, pp. 664–673.
Banaev, E.V., Poliakova, T.A., Tomoshevich, M.A., and Ak-Lama, T.A., Using the ITS2 transcribed spacer of nuclear ribosomal DNA to identify species of the genus Nitraria, BIO Web of Conferences “Plant Diversity: Status, Trends, Conservation Concept,” 2020, vol. 24, artic. no. 00009.
Bobrov, E.G., About Asian species of the genus Nitraria L., Sov. Bot., 1946, vol. 14, no. 1, pp. 19–30.
Bobrov, E.G., On the origin of the desert flora of the Old World in connection with the review of the genus Nitraria L., Bot. Zh., 1965, vol. 50, no. 8, pp. 1053–1067.
Evstifeev, Yu.G. and Rachkovskaya, E.I., On the issue of zoning factors in the southern part of the Mongolian People’s Republic, in Struktura i dinamika osnovnykh ekosistem Mongol’skoi narodnoi respubliki (Structure and Dynamics of Main Ecosystems of Mongolian People’s Republic), Leningrad: Nauka, 1976, pp. 125–144.
Gall, Ya.M. and Popov, I.Yu., Geographic variability and “evolutionary synthesis,” Zh. Obshch. Biol., 1998, vol. 59, no. 3, pp. 302–317.
Il’in, M.M., Nitraria and the origin of desert flora, Priroda, 1944, nos. 5–6, pp. 116–118.
Il’in, M.M., Flora pustyn' Tsentral’noi Azii, ee proiskhozhdenie i etapy razvitiya. Materialy po istorii flory i rastitel’nosti SSSR (Flora of Central Asian Deserts, Its Origin, and Stages of Development. Materials on the History of Flora and Vegetation of the USSR), Moscow: 1958, no. 3, pp. 129–229.
Khudyaev, S.A. and Banaev, E.V., The soil and galogeochemical conditions of locations of the species of the Nitraria (nitrariaceae) genus in the southern part of the Siberian region, Contemp. Probl. Ecol., 2012, vol. 5, no. 6, pp. 597–601.
Komarov, V.L., Introduction to the floras of China and Mongolia, Tr. S.-Peterb. Bot. Sada, 1908, vol. 19, pp. 1–179.
Lukanenkova, V.K., Southeastern Pamir as a botanical refugium, Bot. Zh., 1964, vol. 49, no. 1, pp. 21–30.
Lukin, E.I., Darvinizm i geograficheskie zakonomernosti v izmenenii organizmov (Darwinism and Geographical Patterns in the Change of Organisms), Moscow: Akad. Nauk SSSR, 1940.
Mamaev, S.A., Formy vnutrividovoi izmenchivosti drevesnykh rastenii (Forms of Intraspecific Variability in Woody Plants), Moscow: Nauka, 1972.
Mamaev, S.A., Basic principles of research methods of intraspecific variability of plants, in Individual’naya i ekologo-geograficheskaya izmenchivost' rastenii (Individual and Ecological-Geographical Variability of Plants), Sverdlovsk: Ural. Nauchn. Tsentr Akad. Nauk SSSR, 1975, pp. 3–14.
Mayr, E., Populations, Species, and Evolution: An Abridgment of Animal Species and Evolution, Harvard Univ. Press, 1970.
Nasyrov, R.M. and Sokolov, A.A., Soil cover and soils of foothill stony deserts of the Ili depression, Vestn. Kazan. Gos. Univ., Ser. Geogr., 1998, no. 6, pp. 23–32.
Pavlov, N.V., Flora Kazakhstana (Flora of Kazakhstan), Alma-Ata: Akad. Nauk Kaz. SSR, 1963, vol. 6, pp. 52–53.
Petrov, M.P., On the taxonomy and geography of Nitraria L. in Asia, in Istoriya flory i rastitel’nosti Evrazii (History of Flora and Vegetation of Eurasia), Leningrad, 1972, pp. 156–181.
Polyakova, T.A., Banaev, E.V., and Tomoshevich, M.A., Identification of species in the genus Nitraria L. (Nitrariaceae) based on nucleotide variability of nuclear ribosomal DNA, Vavilovskii Zh. Genet. Sel., 2020, vol. 24, no. 5, pp. 481–488.
Tomoshevich, M.A., Banaev, E.V., and Ak-Lama, T.A., Nitraria komarovii Iljin & Lava ex Bobrov (Nitrariaceae), a new record for the flora of Kazakhstan, Check List, 2019, vol. 15, no. 5, pp. 891–897.
Vasil’eva, L.I., A new species of the genus Nitraria L. from the Pamirs, Nov. Sist. Vyssh. Rast., 1974, vol. 11, pp. 341–344.
Voronkova, M.S., Banaev, E.V., and Tomoshevich, M.A., Comparative study of the composition and content of phenolic compounds of leaves of plants of the genus Nitraria (Nitrariaceae), Khim. Rast. syr’ya, 2017, no. 4, pp. 107–116.
Voronkova, M.S., Banaev, E.V., Tomoshevich, M.A., and Ak-Lama, T.A., Possibilities of using the HPLC method in the taxonomy of the genus Nitraria (Nitrariaceae), BIO Web of Conferences “Plant Diversity: Status, Trends, Conservation Concept,” 2020, vol. 24, artic. n.000.0o. 00096.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest. This article does not contain any studies involving animals performed by any of the authors.
Additional information
Translated by N. Statsyuk
The original online version of this article was revised: the article was mistakenly published as an Open Access article.
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Banaev, E.V., Tomoshevich, M.A. & Ak-Lama, T.A. Morphological Variability of Nitraria Species in Central and Southern Kazakhstan. Contemp. Probl. Ecol. 14, 255–268 (2021). https://doi.org/10.1134/S1995425521030021
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1995425521030021