
Abstract
Precise biostratigraphic dating of the Tojeira Formation (Late Jurassic, Early Kimmeridgian) of the Montejunto section of west-central Portugal, which has yielded important planktonic foraminiferal assemblages, is hindered by poor preservation in the upper part of the section as the lithology shifts from shale to coarser clastics. Assignment was previously made to the Idoceras planula and Sutneria platynota zones based on ammonites. Coccolith and dinoflagellate assemblages described here concur with the Early Kimmeridgian, yet, a finer age constraint is proposed by cyclo- and chemostratigraphical correlation. Peaks in δ 13Corg and TOC, if equivalent to maxima in the envelope of clay/carbonate cycles in SE France, imply that the c. 50 m-thick section spans a 0.8-myr interval of the S. platynota through upper Ataxioceras hypselocyclum ammonite zones, with the approximate base of the A. hypselocyclum Zone at c. 15.4 m (level 13). Such stratigraphy provides new insights into the upper part of the formation by interbasinal correlation with other Tethyan records. An extended first occurrence of the dinoflagellate species Dichadogonyaulax? pannea in the S. platynota Zone is also proposed.
Similar content being viewed by others

Introduction
Several studies have been published on the micropalaeontology of the dominantly clay, marine Tojeira Formation, west-central Portugal. The formation has received attention because of its content with six or more taxa of common to frequent and relatively well-preserved planktonic foraminifera of Kimmeridgian age (Stam 1986; Agterberg et al. 1989; Gradstein 2017; Gradstein et al. 2017). However, the age of the formation as it outcrops in the Montejunto area has received limited attention. Age assignment to the I. planula–S. platynota ammonite zones of early Kimmeridgian age is explained in Atrops and Marques (1986) and Stam (1986). Yet, whilst the lower part of the Tojeira Formation and much of the underlying Montejunto Formation yield an abundance of ammonites, they become rare or absent towards the upper part of the Tojeira Formation. Moreover, macrofossil preservation is poor and many specimens are fragmentary. High-resolution biostratigraphy is hampered by lack of internal biozonation or physical events stratigraphy in the Tojeira Formation. The foraminiferal studies quoted report on few age determinate species which also provide no additional stratigraphic information within the Kimmeridgian.
The Tethyan ammonite zonal scheme (Sub-Mediterranean) has been calibrated to those of the Sub-boreal (Britain) and Boreal (Russian Platform–Arctic) (Gradstein et al. 2012 and references therein). However, due to the provincial distribution of ammonite faunas in the Late Jurassic Boreal seaways and marine shelf margins of the Tethys Ocean, exact interregional correlation is uncertain and an ongoing topic for debate (Wimbledon 2008). The international base of the Kimmeridgian occurs at the base of the Sub-boreal Pictonia baylei Zone and, as now defined, corresponds to the middle of the Epipeltoceras bimammatum Zone in the Tethyan realm or, according to Wierzbowski and Matyja (2014), at the boundary between the Aspidoceras hypselum and E. bimammatum zones. The Tethyan Kimmeridgian base is no longer at the S. platynota Zone, as used by previous studies of the Tojeira-1 section (e.g., Atrops and Marques 1986).
Stratigraphic resolution is increased by calibrating ammonite zones with the coccolith zonation scheme of Casselato (2010) and the dinoflagellate zonation scheme of Riding and Thomas (1988, 1992) and Poulsen and Riding (2003) which have finer subdivisions than either regional ammonite scheme. Considerable literature exists on the ranges of dinoflagellate index species in the Late Jurassic; however, the provincialism of those index species between the north-western Europe faunal realms is less reported on. Ammonite and dinoflagellate assemblages intermediate between the Boreal and Sub-boreal biomes apparently show comparable ranges (Wierzbowski et al. 2002). Poulsen and Riding (2003) also note that, despite marked provincialism, many index species remain of value for local correlation.
Carbon isotope chemostratigraphy is a reliable correlation tool when integrated with biostratigraphy (e.g., Gale et al. 1993; Jarvis et al. 2006; and many others). Reliability, which also may be called stratigraphic fidelity in marking specific levels, is dependent on multiple factors, however. This includes whether there was restricted water mass exchange at the time of deposition which would cause an imprint over the original δ 13C signal. Nevertheless, multiple δ 13Ccarb records exist for the Tethyan Late Jurassic which, when stacked, show regional carbon cycle perturbations over localised records (e.g., Price et al. 2016). Additionally, where relative sedimentation rates are assumed to be steady, cyclostratigraphic correlation with an orbitally tuned coeval section improves age resolution. Comparison of some Kimmeridgian δ 13C records with the Tojeira section δ 13Corg profile is applied as a support in age determination. New geochemical and biostratigraphical data increase stratigraphic resolution of the Tojeira section, as reported on here.
Geological setting
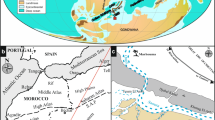
An almost complete, but folded and faulted, sequence of Bathonian through Kimmeridgian marine strata is exposed in the Montejunto area, c. 50 km north of Lisbon. The strata are well exposed on the flanks of Montejunto, a 664-m high diapiric structure, c. 7 km NE of Vila Verde (Fig. 1a). Our stratigraphic account follows that of Stam (1986). From Bathonian through Oxfordian times, a carbonate platform facies persisted, locally of lagoonal facies. An early Kimmeridgian rifting (tectonic subsidence) event, which preceded influx of over 1 km thick marine siliciclastics of the Abadia Formation, caused break-up of the carbonate platform. A transect sketch (Fig. 1b) shows the postulated tectonic-sedimentary setting. Upper Jurassic strata include, from older to younger, the Cabacos, Montejunto, Tojeira, Cabrito, Abadia and Amaral formations, with a total thickness of over 1500 m.

Location of the sections involved (a) and a schematic of the southern slope of Montejunto (b). Numbers 1–7 correspond to the ammonite successions of Atrops and Marques (1986)
Above the disconformity between the Upper Callovian and Lower Oxfordian, the Cabacos Formation comprises about 250 m of platy, thin-bedded grey limestones. The Montejunto Formation comprises thick-bedded, white to grey micritic limestone which alternate with bluish-grey limestones, and grey hard shales, with thicker shales near the top. Several metres wide crinoid-brachiopod biostromes occur in the middle part of the unit. The formation is over 200 m thick with ammonites common to frequent. The Tojeira Formation is over 70 m thick consisting almost entirely of dark grey shales with limonitic concretions and is generally brownish-red towards the middle. Pyritized ammonites are common in the lower part of the unit, and silt content increases considerably near the top; belemnites are rare. The overlying Cabrito, Abadia and Amaral formations together are over 1 km thick with marine sands, silts and conglomerates. Olistolithic limestone blocks and reworked coralline bodies occur, along with sand channel deposits with abundant ripple marks and low-angle cross-bedding.
The Tojeira sections of Stam (1986) and Agterberg et al. (1989) were revisited in 2016 to resample the Tojeira-1 section approximately every metre (Fig. 2). Our 2016 samples M1-1–M1-3 are from a dirt path outcrop of the Montejunto section (Fig. 1), but are not reported on here. The lowest sampling level from the Tojeira-1 section is approximately at the base of the stratigraphic column in Fig. 2—see Gradstein (2017) and Gradstein et al. (2017) for full locality description. Our investigations reveal that the Tojeira-2 section in cross section 2 of Stam (1986), between the village of Tojeira and Pereiro, had largely vanished due to domestic construction and agriculture. A new outcrop, Tojeira-3 section (Fig. 1a), is exposed halfway along the road from Montejunto to Vila Verde dos Francos. Sandy siliclastics of the upper part of the outcrop probably indicate upper Tojeira or lower Cabrito Formation, but is not reported on here (see Gradstein 2017).
Biostratigraphy
The Tojeira Formation was assigned to the I. planula–S. platynota ammonite zones by several authors (Mouterde et al. 1971, 1973, 1979; Atrops and Marques 1986; Stam 1986). The boundary between the two zones can be identified by the first occurrence datum (FAD) of S. platynota (Moliner and Olóriz 2009). However, in general, occurrences of S. platynota reduce towards the upper part of the S. platynota Zone. The base of the following A. hypselocyclum Zone is defined by the FAD of Ataxioceras and last occurrence (LAD) of S. platynota. Age controls for the section are reviewed with consideration of new material presented here.
Ammonites
The boundary between the Tojeira Formation and underlying Montejunto Formation is represented by succession 2 of Atrops and Marques (1986, Fig. 3). Faunal successions of Atrops and Marques (1986) are shown in Fig. 1b. Ammonites are considered rare, but include Epipeltoceras bimammatum suggesting a latest Oxfordian–Early Kimmeridgian E. bimammatum Zone age. The two identifications of Idoceras (Subnebrodites) planula indicating the I. planula Zone come from the overlying succession. Succession 4 is not accessible due to an accumulation of modern deposits within the valley (see Fig. 1). Successions 5–7 are equivalent to our Tojeira-1 section, the base of which overlies a conspicuous limestone band (see Appendix Fig. 5). However, at this horizon Atrops and Marques (1986) instead assigned the top of the Tojeira Formation and the base of the Abadia Formation; the Cabrito Formation was not studied. At the base of succession 5, S. platynota and Orthosphinctes polygyratus are considered fairly abundant. Atrops and Marques (1986) suggest the Oxfordian–Kimmeridgian boundary (which is now equivalent to the I. planula–S. platynota zonal boundary) occurs at this horizon at the top of succession 4. Succession 5 is attributed to the S. platynota Zone, sub-zone O. polygyratus. Sutneria platynota is also present in succession 6, alongside Taramelliceras (Metahaploceras) aff. nodosiusculum in the top part which characterises the younger A. hypselocyclum Zone.
Stam (1986) defined the Tojeira Formation based on ammonites identified by T. Poulton in 1983 [Report No. J-3-TTP-e1983 in Stam (1986), Appendix]. Of the 11 collections of ammonites, 4 were collected from the Tojeira shales (at the same site as the Tojeira-1 section), the remainder from the Montejunto and Cabaco limestones (Fig. 1b). Perisphinctes (Progeronia) (?) or Lithacoceras (?) sp. is reported from sample 6.10/6.11 (of Stam’s 1986 sample numbers) and one unknown horizon, and “Perisphinctes” sp. from samples 6.22 and 6.4. Stam’s (1986) sample numbers described here for the Tojeira Formation equate approximately to the lowermost part of the Tojeira-1 section. The few subgeneric assignments given were noted as tentative at best. This included the Progeronia triplex group of Perisphinctids which constrained the interval to the lowermost Kimmeridgian along with O. polygyratus which is reported from the Oxfordian–Kimmeridgian boundary (Besems and Love 1988; Matyja and Wierzbowski 2003). However, for the most part only juvenile or inner whorls were found, hindering confident differentiation of the dominant group, perisphinctids, which mostly relies on size, apertural modifications and ribbing modification of the adult. Other age constraints are often conflicting. Pertisphinctes in samples 6.10/6.11 have some resemblance to Taramelliceras pseudoflexuosa (now T. compsum). This suggests a younger age as this species has been reported mostly from the Aspidoceras acanthicum Zone (Baudouin et al. 2011) and no older than the A. hypselocyclum Zone (Pszczółkowski et al. 2016, Fig. 14). Additionaly, Perisphinctes danubiensis identified within sample 6.4 may suggest a lower Tithonian age (Schweigert and Scherzinger 2004).
There is also uncertainty regarding the precise age of the underlying Montejunto Formation. Of the five collections of ammonites, the age from two cannot be determined, and of the remaining three there are sample numbers for two collections, only. T. Poulton recognised in Stam’s (1986) sample 5.7 that P. (Orthosphinctes) suggests the assemblage probably belongs to the I. planula Zone; however, some specimens [i.e., Perisphinctes (Arisphinctes) (?) sp.] are similar to older (lower Perisphinctes bifurcatus Zone) species elsewhere. Stam’s (1986) sample 14.2 is likely the Aspidoceras hypselum Zone as defined by T. Poulton; however, the P. (Arisphinctes?) is noted to have similarities to upper Perisphinctes plicatus–Gregoryceras transversarium faunas. Overall age inferences for the Monetejunto ammonite collections range from the Aspidoceras hypselum Zone to as old as the Perisphinctes plicatilis zones.
Foraminifera
The micropaleontology of the Tojeira Formation, studied by Stam (1986), Agterberg et al. (1989), Gradstein (2017) and Gradstein et al. (2017), revealed common to abundant Jurassic planktonic foraminifera. Many samples, particularly in the middle part of the Tojeira unit, are rich in specimens using fractions of 65–125 µm and 125–180 µm of washed residues. Hundreds of well-preserved specimens may be picked from the washed residues in a relatively short time.
In the middle and upper part of the Tojeira Formation, several picked residues of the Stam (1986) and Agterberg et al. (1989) collections were re-studied in detail by Gradstein (2017). Based on the morphological variability observed, six or more Jurassic planktonic foraminiferal types, all microperforate (forms which have <1 μm pore-diameters) were distinguished in the samples. Portuguese Jurassic planktonic foraminifera include Globuligerina oxfordiana, G. balakhmatovae, G. tojeiraensis, G. bathoniana, Conoglobigerina grigelisi and C. helvetojurassica. This assemblage is considered to be typical for Kimmeridgian strata (Stam 1986; Gradstein 2017). The assemblage occurs together with Pseudolamarckina rjasanensis and a rich epistominid (of the genera Epistomina) assemblage, including E. mosquensis and E. uhligi. Mjatliuk (1953) describes P. rjasanensis as rare to common in the Middle and Upper Callovian clays, yet, abundant in the argillaceous Kimmeridgian deposits. E. mosquensis is typical of the Callovian (Kalantari 1969; Pandey and Dave 1993) and its range top is considered characteristic of the Callovian/Oxfordian boundary in India by some workers (Subbotina et al. 1960; Talib et al. 2007). However, in the Grand Banks, Canada, the highest stratigraphical occurrence of E. mosquensis, alongside E. uhligi, extends the Pseudocyclammina jaccardi formaniferal Zone of Late Oxfordian–Early Kimmeridgian age (Gradstein 1979; Stam 1986). The stratigraphic range at the Grand Banks section is a more reliable comparison given the relative close proximity to the Lusitanian basin in the Late Jurassic, whereas India lay in the Southern Hemisphere.
According to Pazdro (1969), epistominids are restricted to silty–clayey–marly sediments in which E. mosquensis and E. uhligi are abundant and P. rjasanensis is frequent. The increased calcareous planktonic sedimentation may have improved the preservation potential of epistominids tests (including E. mosquensis). Agglutinated foraminiferal taxa and micro-gastropods are also abundant in the section. Discorbis paraspis and D. scutiliformis are rare. This benthic foraminiferal assemblage is only known from deeper neritic and bathyal environments (Stam 1986), in agreement with a deeper, basinal setting.
Dinoflagellate cysts and other palynomorphs
Samples were selected at regular intervals throughout the Tojeira-1 section for standard palynological processing. Processing included disaggregation by hydrochloric and hydrofluoric acids of approximately 25 g of sample, sieving and separation of organic material using sodium polytungstate with a specific gravity of c. 2.1.
Index species were recoverable from levels 2, 4, 24 and 26 in low numbers. Few other cysts occur at levels 19 and 48, but for the most part, they are absent within the remaining palynomorph assemblage. Identified dinoflagellate cysts in order of most to least common are as follows, Gonyaulacysta jurassica jurassica, Cribroperidinium spp., Leptodinium subtile, Systematophora areolata, Meiourogonyaulax sp., Dichadogonyaulax? pannea, Rhynchodiniopsis cladophora, Leptodinium eumorphum, Systematophora orbifera, Systematophora spp., Occisucysta sp., Tehamadinium cf. aculeatum. and Endoscrinium luridum. The pollen–spore associations are of low diversity. These include bisaccate pollen, Classopollis classoides, Callialasporites dampieri, Cerebropollenites mesozoicus and Deltoidospora sp., along with frequent foraminiferal linings, terrestrially derived material (i.e., wood) and sphaeromorphs abundant throughout.
The palynological assemblage is typical of the Late Oxfordian–Kimmeridgian dinoflagellate cyst interval zones Scr–Elu of Riding and Thomas (1988, 1992). Despite poor preservation and low abundance of dinoflagellate cysts, a number of index species suggest a Kimmeridgian age. Tehamadinium cf. aculeatum occurs at level 4. This species has a base in the Oxfordian in the Sub-boreal realm. Occurrences of this species in England range from the Late Oxfordian (Thomas and Cox 1988) to the Aulacostephanus mutabilis/Aulacostephanus eudoxus zonal boundary (Riding and Thomas 1988). The Tojeira-1 section is no younger than Kimmeridgian, given the range tops of G. jurassica jurassica and E. luridum which coincide in the Aulacostephanus autissiodorensis Zone (Riding and Thomas 1992; Bailey et al. 1997; Poulsen and Riding 2003). Rhynchodiniopsis cladophora is typical also for the Kimmeridgian with only rare records reported of this species above the A. eudoxus Zone (Riding and Thomas 1992). The spore and pollen associations are typical for the Late Jurassic, but are of little value for precision biostratigraphy.
The presence of Dichadogonyaulax? pannea, however, at level 2 suggests a conflicting age of no older than the species’ first occurrence in the Sub-boreal A. mutabilis Zone (Riding and Thomas 1992). This age is at least two Tethyan ammonite zones younger than the age given to the Tojeira-1 section by Stam (1986), and given the ammonite biostratigraphy, a younger age at this level is improbable. More likely, the species’ first occurrence may have been earlier in the Tethyan Lusitanian basin. The first occurrence of D. pannea as a zonal index species is mostly founded on Sub-boreal records. In England, it appears in the Early A. eudoxus Zone in Lincolnshire (Riding 1987) or younger still, the Pectinatites wheatleyensis Zone in Dorset (Riding and Thomas 1988). In the British–Danish sector of the North Sea it is recorded in the Late Kimmeridgian (Poulsen and Riding 2003) and in Poland, Poulsen (1994) notes its first occurrence in the Early Tithonian. It has become a biostratigraphic marker for Late Kimmeridgian–Tithonian assemblages (Riley 1980; Bailey et al. 1997, Poulsen and Riding 2003). The species’ range is reported to extend no further than the Paracraspedites oppressus Zone in the Sub-boreal realm (Riding and Thomas 1992); however, in the western Barents Sea, it extends to the Ryazanian (Smelror and Dypvik 2005). The range of D. pannea may consequently differ in the Tethyan realm. Hamad and Ibrahim (2005, Fig. 5) illustrate the range base of D. pannea within the Late Oxfordian of Qatar, though the authors do not denote the source clearly. Likewise, elsewhere in Portugal, Taylor et al. (2014) find D. pannea throughout the Lourinhã Formation (A. eudoxus–Hybonoticeras hybonotum Tethyan zones) and reference its base within the A. hypselocyclum Zone, Early Kimmeridgian (see Taylor et al. 2014, Fig. 6). However, those references sourced either do not mention D. pannea or are unobtainable by the author. Ied and Lashin (2016) list D. pannea within an assemblage of Oxfordian–Kimmeridgian species from Egypt, but, a range base is not given. Clearly, discrepancy exists regarding the first occurrence of D. pannea.

Lithostratigraphy of the Montejunto area and stratigraphic log of the Tojeira-1 section alongside δ 13C, TOC, CaCO3 and reworking data. Smoothing, via a 4-point moving average has been applied to the δ 13C, TOC and CaCO3 records (overlain in blue)
Depositional Environment
The low-diversity dinoflagellate cyst assemblage is typical of other Kimmeridgian Portuguese biotas (e.g., Borges et al. 2011). Assemblages from further north in Europe have markedly higher diversities (i.e., Riding and Thomas 1988). This may be due to partial restriction in the Lusitanian basin preventing some mixing of the dinoflagellates with areas outside Portugal. The depositional environment would have likely been within relatively deep, warm, marine waters; the majority of the dinoflagellate taxa have such palaeoecological affinities (Riding and Hubbard 1999) and the epistominid assemblage is indicative of bathyal envrionments. However, a proximal setting is likely given the abundance of sphaeromorphs (Stricanne et al. 2004) and the high abundance of Classopollis, a suggested proxy for regions marginal to bodies of water (Vakhrameyev 1982). Also, Classopollis classoides mostly remains intact as tetrad or quads. This indicates that transportation distances and energy levels were likely low, and the poor preservation of dinoflagellate cysts is not an artefact of environmental stress.
Coccoliths
Calcareous nannofossils occur through the sampled section as moderate to poorly preserved assemblages constituting approximately 2–5% of the sediment by volume. Although the bulk of the specimens are considered to be autochthonous, there is a significant and, in some samples, substantial component of reworked nannofossils (discussed below). The authochthonous assemblage is dominated by Watznaueria barnesiae and Ellipsogelasphaera communis, which together comprise an average of about 80% of the assemblages. The presence of Favoconus multicolumnatus without any Conusphaera indicates Zone NJT14 of Casselato (2010). This placement is corroborated by the occurrences of Calcivascularis cassidyi, Staurolithites lumina and Crepidolithus perforatus. Staurolithites lumina has its first appearance at level 5 and occurs consistently in low numbers throughout the rest of the section. Bergen et al. (2014) place the first appearance of S. lumina in the upper part of the I. planula Zone, implying similar placement of the Tojeira datum. Calcivascularis cassidyi occurs as single specimens in samples below level 38, but is consistently present as multiple specimens in samples from levels 38–48. This horizon (level 38) may correspond to the “First Regular Occurrence” datum of Bergen et al. (2014), which those authors place in the upper I. planula Zone. However, this disagrees with the ammonite biostratigraphy which indicates the I. planula Zone terminates at or below the base of the Tojeira-1 section. Crepidolithus perforatus (=Millbrookia perforata) occurs sporadically through the entire section at Tojeira, including the uppermost sample at level 48. On TS Creator (Version 7.0), the last appearance of this species is placed at or near the base of the A. hypselocyclum Zone (Ogg et al. 2016). This is based on interpretation by Bown and Cooper (1998) that although its final occurrence lies within the Tithonian, in the P. baylei Zone C. perforatus becomes rare/sporadic. If accurate, this datum provides the means to restrict the Tojeira-1 section to the mid-I. planula to upper S. platynota zones of the Early Kimmeridgian. However, Bergen et al. (2014) place this datum significantly higher (H. beckeri Zone) in the Late Kimmeridgian. Consequently, the age of the top of the Tojeira-1 section cannot be definitively placed using calcareous nannofossils.
Reworking
There are 11 coccolith species occurring in the Tojeira-1 section that are clearly reworked, as their ranges end prior to the Kimmeridigan. At least two different source ages are indicated by these taxa: Pliensbachian (Early Jurassic) and Bajocian (Middle Jurassic). The coccolith assemblages in levels 20–21 are characterised by at least 24 and 21.5%, respectively, of specimens that have been clearly reworked. This reworking event and the distribution of demonstrably reworked specimens compared to the total count of autochthonous specimens are shown in Fig. 2. Detrital material would impose a false δ 13C value in the record (e.g., Melchin and Holmden 2006); therefore, the δ 13Corg values between levels 20–21 are considered unreliable. Elsewhere in the sequence, there is a count of no more than ~2% reworked/in situ specimens. Although this is a minimum expectation, the low proportions are considered to have a negligible effect on the C-isotope values.
Chemostratigraphy
Physical properties
To acquire both TOC and δ 13C values for the Tojeira-1 section, fresh sediment chips (<10 g) were ground in a clean lab and treated with a solution of 5% HCl. Between 1 and 3 mg of dried sample was sealed within a Sn (tin) bucket, weighed and analysed by 1020 °C combustion on a Flash 2000 Organic Elemental Analyzer and isotope ratio mass spectrometry was performed with a MAT 253 at the stable isotope geochemistry laboratory at the Open University, Milton Keynes, UK. The results were calibrated using repeated measurements of in house reference solutions and international standards. Rock–Eval data were provided by L R. Snowdon (Geological Survey of Canada) on behalf of B. Stam in 1985. The results presented here are based on 15 samples provided from Tojeira-1 section collected by Stam (1986) and are correlated to δ 13C and TOC values presented in this study.
The δ 13Corg record of the Tojeira-1 section deviates little from an average value of −23.2‰. Heaviest values of −24.4 and 23.2‰ occur as peaks within a distinct couplet at levels 17 and 20 and correspond to a break in the oscillatory pattern in the TOC record. The TOC curve is mostly smooth between levels 13 and 21. After this interval in the δ 13Corg record, there is a discrete shift from −22.6‰ at level 27 to 23.4‰ at level 39. There is a comparably progressive decline in the TOC record for this interval; however, overall there is a very low content of organic carbon (0.4–1.2 wt‰).
Correlation between existing δ 13C records with that of the Tojeira-1 section refines the biostratigraphical age range provided above. Reliability is dependent on multiple factors, however, including the possibility of local palaeoceanographic imprint on the overall isotopic composition of inorganic dissolved carbon in the oceans. The influence on δ 13Corg by local processes is considered unlikely for the Tojeira-1 section. There is no correlation between δ 13Corg and TOC (R 2 = 0.2). Likewise, the relationship between Rock–Eval parameters and δ 13Corg is R 2 = 0.2 (T max) and R 2 = −0.5 (HI). The majority (76%) of T max values are below 435 °C indicating shale immaturity (Hunt 1996). Thus, the δ 13Corg profile is unlikely to have been influenced by thermal degradation. Interpretation of T max values is, however, tentative due to low TOC values and thus difficulty in measuring small S2 peak areas.
Time series analyses
Programs used to carry out time series analyses in the depth domain included PAST v.3.15 (Hammer et al. 2001) and Redfit3.8 (Schulz and Mudelsee 2002), a program specifically designed to analyse time series with uneven sampling rates. The C-isotope and TOC records appear to show regular cycles. The Redfit power spectrums of δ 13Corg and TOC (Fig. 3) both reveal a significant peak above the 95% false-alarm level (FAL) at a frequency of 25.9 m/cycle. A second prominent peak occurs above the 85% FAL at 6.47 and 7.4 m/cycle in the TOC and δ 13Corg records, respectively. Bandpass filters centred on the 26 and 6.5 m/cycle spectral peaks have been applied to the TOC data using PAST v.3.15. The data were regularly interpolated and the mean was subtracted. Although the section is short, the larger frequency (25.9 m/cycle) may represent the 405-kyr-long eccentricity cycle, which is the most reliable Milankovitch cycle for the Mesozoic (Laskar et al. 2004). This is supported by the presence of the 6.47 m/cycle frequency in the TOC record which occurs four times within the larger 25.9 m/cycle frequency. This may represent short-term eccentricity. Statistical reliability is, however, reduced as the larger frequency appears less than three times. Therefore, identification of this frequency as an orbital cycle is proposed, but not verifiable without further study of the lower part of the Tojeira Formation to test if this periodicity continues downwards.
Correlation
We compared the interpreted 405-kyr cycles at Tojeira to those interpreted by Boulila et al. (2008) at La Méouge, southeastern France in Fig. 3. A 25.7-kyr/cycle bandpass shows precession periodicity at La Méouge modulated by long-term eccentricity (Boulila et al. 2008). We identified two maxima in long-term wavelength cyclicity at the Tojeira-1 section and, assuming a steady sedimentation rate, correlated the cycles to Max2–3 of Boulila et al. (2008) using the ammonite zonal boundaries of the Tojeira Formation proposed by Atrops and Marques (1986). Thus, the A. hypselocyclum Zone falls approximately at Max2, and the I. planula–S. platynota boundary below the Tojeira-1 section. This age refinement is in agreement with the ammonite biostratigraphy of Atrops and Marques (1986), where the I. planula–S. platynota boundary is suggested probably at the top of their succession 4 (Fig. 1b), and the A. hypselocyclum Zone within the upper part of succession 6. The Early Kimmeridgian S. platynota–C. divisum zones were orbitally calibrated by Boulila et al. (2008) and provide a reliable age constraint for interbasinal correlation. The Tojeria-1 section is too short to statistically confirm the presence of an orbital periodicity and, therefore, correlation to eccentricity cycles at La Méouge is uncertain. However, the long- and short-term eccentricity cycles suggested at the Tojeira-1 section plausibly correlate to the eccentricity cycles at La Méouge. This proposes precision in the ammonite zonation scheme in the absence of a well-constrained biostratigraphy in the upper part of the Tojeira-1 section. Correlation of the δ 13Corg excursions, synchronous with peaks in TOC, to the minima in oscillations in SE France would not match with the biostratigraphic controls. For example, to instead correlate level 13 to the first minima in the La Méouge section, would reduce the S. platynota Zone beneath horizons where S. platynota were identified. Similarly, if level 13 correlated with the second minima between Max2–3 (level 23), this would extend the S. platynota Zone into Atrops and Marques’ (1986) succession 6, where the younger A. hypselocyclum Zone is represented according to their ammonite identifications.

Cyclostratigraphic correlation based on long-term eccentricity cycles between La Méouge (solid black curve adjacent to the precession bandpass) (from Boulila et al. 2008) and the Tojeira-1 section bandpassess based on Redfit spectral analyses. MS magnetic susceptibility
Correlation of the organic carbon isotope record for the Tojeira-1 section with that taken from other sections provides additional age support. At the onset of the Late Jurassic, there was widespread black shale deposition across north-western Europe (UK, North Sea, Lusitanian Basin; Agterberg et al. 1989); however, organic carbon records in the Tethyan realm are uncommon. This is due to the dominance of carbonate supply in the Tethyan realm over clastic in Boreal-Arctic latitudinal successions. Therefore, isotope data are mostly derived from carbonate carbon in the Tethyan realm and organic carbon in the Boreal realm. Boreal realm organic carbon records from age equivalent sections in the UK (Morgans-Bell et al. 2001), Scotland (Nunn et al. 2009) and Russia (Riboulleau et al. 1998; Price and Rogov, 2009) show a conspicuous positive excursion in the R. cymodoce Zone (Sub-boreal equivalent to the S. platynota/A. hypselocyclum zones) and are correlated here to the Tojeira-1 section C-isotope record (Fig. 4). The lowermost Kimmeridgian ammonite zone (Pictonia baylei) is represented by an interval of stratigraphic incompleteness in the type section of the Kimmeridge Clay Formation (KCF) in Dorset (Morgans-Bell et al. 2001) and, therefore, this zone cannot be correlated to. However, although there is evidence for erosional surfaces in the overlying R. cymodoce–A. mutabilis zones in the KCF type section cores, a minor positive excursion is still apparent (Fig. 4). Likewise, from the Isle of Skye, Scotland, there is a peak in the C-isotope record in the R. cymodoce Zone (Nunn et al. 2009), and in the carbonate carbon record for Makariev village, Russia, is a definitive positive peak (Riboulleau et al. 1998; Price and Rogov, 2009). The cyclostratigraphical age suggestion for the Tojeira-1 section (constrained by biostratigraphy) places a conspicuous peak of similar scale to those Boreal records outlined, in the early A. hypselocyclum Zone. This compares well to the aforementioned records indicating a reliable age tiepoint.

C-isotope correlation of the Tojeira-1 section with a Boreal organic and carbonate carbon record stack (Dorset, Morgans-Bell et al. 2001; Scotland, Nunn et al. 2009; Russia, Riboulleau et al. 1998; Price and Rogov 2009) and a Tethyan δ 13Ccarb stack from Price et al. (2016). See Price et al. (2016) for a full list of references and details on the sections they included. Of the Price et al. (2016) stack, the Gorges du Pichoux, Swiss Jura record was not included due to δ 13C interference from localised factors, and as the Lókút Hill section had very few data points for the Early Kimmeridgian to interpret meaningful shifts, the record was not included in the Tethyan stack here
This minor excursion is present in both the organic and carbonate carbon records from Scotland, as a positive fluctuation in the R. cymodoce Zone (Nunn et al. 2009). However, as shown by an overlay of δ 13Ccarb Tethyan records of southern Europe by Price et al. (2016), a minor positive peak in the A. hypselocyclum Zone is not clearly recognisable. Indeed, at the Oxfordian–Kimmeridgian boundary is a pronounced positive shift that declines across the lowermost Kimmeridgian Tethyan ammonite zones, appearing to culminate over the S. platynota–A. hypselocyclum zonal boundary in some accounts (see Fig. 4). Thereafter, average values appear to steady, apart from in Długa Valley, Poland which shows a positive shift at this point, similar to the Boreal carbonate and organic records (Fig. 4) (Jach et al. 2014).
Conclusions
High-resolution biostratigraphic dating of the Tojeira Formation at Montejunto is limited. Reworking is evident in the coccolith assemblages which suggest two separate source ages, and much of the section sampled for palynology is barren in dinoflagellates. Those that are recoverable in the lower part of the section are infrequent and of reduced diversity. The index species do, however, concur with a Kimmeridgian age, as do the autochthonous coccolith assemblages. The abundance of ammonites is similarly poor, and the preservation significantly declines towards the overlying Cabrito Formation where silt content increases dramatically, as is also apparent in the Tojeira-3 section (Fig. 1a). Although some key stratigraphic taxa are recoverable, indicating ammonite zones S. platynota–A. hypselocyclum (Atrops and Marques 1986), the zonal boundaries are tentative. The application of geochemical data presented here via chemo- and cyclostratigraphic correlation, refines those boundaries more precisely. High and low frequencies are shown in the Tojeira-1 section by spectral analyses, although the section is short and only two long-term wavelength periodicity cycles, possibly in the order of 405 kyr, are present. However, we suggest that the recurrence of two maxima can be fitted to an orbitally tuned coeval section in southeastern France (Boulila et al. 2008). This cyclostratigraphy is not a primary age control, but a supportive tool to increase the age resolution. Whereas previous dating methods for the Tojeira Formation are reliant on preservation of key taxa, the combination of biostratigraphy with geochemical correlation in our integrated approach improves the age determined of the section as a whole.
References
Agterberg, F. P., Gradstein, F. M., & Nazli, K. (1989). Correlation of Jurassic microfossil abundance data from the Tojeira sections, Portugal. Geological Survey Canada, 89–9, 467–482.
Atrops, F., & Marques, B. (1986). Mise en évidence de la zone à Platynota (Kimméridgien inférieur) dans le massif du Montejunto (Portugal); Conséquences stratigraphiques et paléontologiques. Geobios, 19(5), 537–547. (In French).
Bailey, D., Milner, P., & Varney, T. (1997). Some dinoflagellate cysts from the Kimmeridge Clay Formation in North Yorkshire and Dorset, UK. Proceedings of the Yorkshire Geological and Polytechnic Society 51(3). Geological Society of London. pp. 235–243.
Baudouin, C., Boselli, P., & Bert, D. (2011). The Oppeliidae of the Acanthicum zone (Upper Kimmeridgian) from Mount Crussol (Ardèche, France: ontogeny, variability and dimorphism of the genera Taramelliceras and Streblites (Ammonoidea). Revue de Palébiologie, 30(2), 619–684.
Bergen, J. A., Boesiger, T. M., & Pospichal, J. J. (2014). Low-latitude Oxfordian to Early Berriasian Nannofossil biostratigraphy and its application to the subsurface of eastern Texas. In U. Hammes & J. Gale (Eds.), Geology of the Haynesville Gas Shale in East Texas and West Louisiana U.S.A (Vol. 105, pp. 69–102). Tulsa: American Association of Petroleum Geologists.
Besems, R., & Love, C. (1988). A palynological investigation of some Kimmeridgian deposits from Spain. Journal of Micropalaeontology, 7(2), 217–232.
Borges, M. E., Riding, J. B., Fernandes, P., & Pereira, Z. (2011). The Jurassic (Pliensbachian to Kimmeridgian) palynology of the Algarve Basin and the Carrapateira outlier, southern Portugal. Review of Palaeobotany and Palynology, 163(3), 190–204.
Boulila, S., Galbrun, B., Hinnov, L. A., & Collin, P. Y. (2008). Orbital calibration of the Early Kimmeridgian (southeastern France): implications for geochronology and sequence stratigraphy. Terra Nova, 20(6), 455–462.
Bown, P. R., & Cooper, M. K. E. (1998). Jurassic. In P. R. Bown (Ed.), Calcareous nannofossil biostratigraphy (pp. 34–85)., British micropalaeontological society publication series London: Chapman & Hall.
Casselato, C. E. (2010). Calcareous nannofossil biostratigraphy of Upper Callovian-Lower Berriasian successions from the Southern Alps, North Italy. Rivista Italiana di Paleontologia e Stratigrafia, 116, 357–404.
Gale, A. S., Jenkyns, H. C., Kennedy, W. J., & Corfield, R. M. (1993). Chemostratigraphy versus biostratigraphy: data from around the Cenomanian–Turonian boundary. Journal of the Geological Society, 150(1), 29–32.
Gradstein, F. M. (1979). Jurassic micropaleontology of the Grand Banks. Ciências da Terra, Lisboa, 5, 86–96.
Gradstein, F. M. (2017). New and emended species of Jurassic Planktonic Foraminifera. Swiss Journal of Palaeontology, 1–25.
Gradstein, F. M, Gale, A. S, Kopaevich, L., Waskowska, A., Grigelis, A., & Glinskikh, L. (2017). The planktonic foraminifera of the Jurassic. Part I: material and taxonomy. Swiss Journal of Palaeontology, 1–71.
Gradstein, F. M., Ogg, J. G., Schmitz, M., & Ogg, G. (2012). The geologic time scale 2012. Boston: Elsevier.
Hamad, A. L., & Ibrahim, M. I. A. (2005). Facies and palynofacies characteristics of the Upper Jurassic Arab D reservoir in Qatar. Revue de Paléobiologie, Genève, 24(1), 225–241.
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4(1), 9.
Hunt, J. M. (1996). Petroleum geology and geochemistry (p. 743). New York: W.H Freeman & Co.
Ied, I. M., & Lashin, G. M. (2016). Palynostratigraphy and paleobiogeography of the Jurassic-Lower Cretaceous succession in Kabrit-1 well, northeastern Egypt. Cretaceous Research, 58, 69–85.
Jach, R., Djerić, N., Goričan, S., & Reháková, D. (2014). Integrated stratigraphy of the Middle–Upper Jurassic of the Krížna Nappe, Tatra Mountains. Annales Aocietatis Geologorum Poloniae, 84, 1–33.
Jarvis, I. A. N., Gale, A. S., Jenkyns, H. C., & Pearce, M. A. (2006). Secular variation in Late Cretaceous carbon isotopes: a new δ 13C carbonate reference curve for the Cenomanian–Campanian (99.6–70.6 Ma). Geological Magazine, 143(05), 561–608.
Kalantari, A. (1969). Foraminifera from the middle Jurassic-Cretaceous successions of Koppet-Dagh region (NE Iran). Tehran: National Iranian Oil Company, Geological Laboratories.
Laskar, J., Correia, A. C. M., Gastineau, M., Joutel, F., Levrard, B., & Robutel, P. (2004). Long term evolution and chaotic diffusion of the insolation quantities of Mars. Icarus, 170(2), 343–364.
Matyja, A. B., & Wierzbowski, A. (2003). Boreal and Subboreal ammonites in the Submediterranean uppermost Oxfordian in the Bielawy section (northern Poland) and their correlation value. Acta Geologica Polonica, 52(4), 411–421.
Melchin, M. J., & Holmden, C. (2006). Carbon isotope chemostratigraphy in Arctic Canada: sea-level forcing of carbonate platform weathering and implications for Hirnantian global correlation. Palaeogeography, Palaeoclimatology, Palaeoecology, 234(2), 186–200.
Mjatliuk, E. V. (1953). Fossil foraminifera of the USSR. Spirillinidae, Rotalliidae, Epistominidae, and Asterinidae. VNIGRI, 71, 1–271. (In Russian).
Moliner, L., & Olóriz, F. (2009). Correlation potential of the Upper Jurassic (lower Kimmeridgian) Platynota Chronozone deposits in northeastern Spain. Journal of the Geological Society of Sweden, 131(1–2), 205–213.
Morgans-Bell, H. S., Coe, A. L., Hesselbo, S. P., Jenkyns, H. C., Weedon, G. P., Marshall, J. E. A., et al. (2001). Integrated stratigraphy of the Kimmeridge Clay Formation (Upper Jurassic) based on exposures and boreholes in south Dorset, UK. Geological Magazine, 138(05), 511–539.
Mouterde, R., Ramalho, M., Rocha, R. B., Rüget, C., & Tintant, H. (1971). Le Jurassique du Portugal. Esquisse stratigraphique et zonale. Bulletin, Geological Society of Portugal, Lisbon, 18, 73–104. (In French).
Mouterde, R., Rocha, R. B., Ruget, C., & Tintant, H. (1979). Faciès, biostratigraphie et paléogéographie du Jurassique portugais. Ciências da Terra, Lisboa, 5, 29–52. (In French).
Mouterde, R., Ruget, C., & Tintant, H. (1973). Le passage Oxfordien–Kimmeridgien au Portugal (regions de Torres Vedras et du Montejunto). Comptes Rendus. Académie des Sciences (Paris), 277, 2645–2648. (In French).
Nunn, E. V., Price, G. D., Hart, M. B., Page, K. N., & Leng, M. J. (2009). Isotopic signals from Callovian-Kimmeridgian (Middle–Upper Jurassic) belemnites and bulk organic carbon, Staffin Bay, Isle of Skye, Scotland. Journal of the Geological Society, 166(4), 633–641.
Ogg, J. G., Ogg, G., & Gradstein, F. M. (2016). A concise geologic time scale 2016. Boston: Elsevier.
Pandey, J., & Dave, A. (1993). Studies in Mesozoic foraminifera and chronostratigraphy of western Kutch, Gujarat. Paleontographica Indica, Dehradun, 1, 1–221.
Pazdro, O. (1969). Middle Jurassic Epistominidae (Foraminifera) of Poland. Studia Geologica Polonica, 27, 1–92.
Poulsen, N. E. (1994). Dinoflagellate cyst biostratigraphyof the Late Jurassic of Poland. Geobios, 27, 401–407.
Poulsen, N. E., & Riding, J. B. (2003). The Jurassic dinoflagellate cyst zonation of Subboreal Northwest Europe. Geological Survey of Denmark and Greenland Bulletin, 1, 115–144.
Price, G. D., Főzy, I., & Pálfy, J. (2016). Carbon cycle history through the Jurassic–Cretaceous boundary: A new global δ13C stack. Palaeogeography, Palaeoclimatology, Palaeoecology, 451, 46–61.
Price, G. D., & Rogov, M. A. (2009). An isotopic appraisal of the Late Jurassic greenhouse phase in the Russian Platform. Palaeogeography, Palaeoclimatology, Palaeoecology, 273(1), 41–49.
Pszczółkowski, A., Grabowski, J., & Wilamowski, A. (2016). Integrated biostratigraphy and carbon isotope stratigraphy of the Upper Jurassic shallow water carbonates of the High-Tatric Unit (Mały Giewont area, Western Tatra Mountains, Poland). Geological Quarterly, 60(4), 893–918.
Riboulleau, A., Baudin, F., Daux, V., Hantzpergue, P., Renard, M., & Zakharov, V. (1998). Évolution de la paléotempérature de eaux de la plate-forme russe au cours du Jurassique supérieur. Comptes Rendus de l’Académie des Sciences Série II, 326, 239–246.
Riding, J. B. (1987). Dinoflagellate cyst stratigraphy of the Nettleton Bottom Borehole (Jurassic: Hettangian to Kimmeridgian), Lincolnshire, England. Proceedings of the Yorkshire Geological Society, 46(3), 231–266.
Riding, J. B., & Hubbard, R. N. (1999). Jurassic (Toarcian to Kimmeridgian) dinoflagellate cysts and paleoclimates. Palynology, 23(1), 15–30.
Riding, J. B., & Thomas, J. E. (1988). Dinoflagellate cyst stratigraphy of the Kimmeridge Clay (Upper Jurassic) from the Dorset coast, southern England. Palynology, 12(1), 65–88.
Riding, J. B., & Thomas, J. E. (1992). Dinoflagellate cysts of the Jurassic System. In A. J. Powell (Ed.), A stratigraphic index of dinoflagellate cysts (pp. 7–97). London: Chapman & Hall.
Riley, L. A. (1980). Palynological evidence of an early Portlandian age for the uppermost Helmsdale Boulder Beds, Sutherland. Scottish Journal of Geology, 16(1), 29–31.
Schulz, M., & Mudelsee, M. (2002). REDFIT: estimating red-noise spectra directly from unevenly spaced paleoclimatic time series. Computers and Geosciences, 28(3), 421–426.
Schweigert, G., Scherzinger, A. (2004). New efforts for a revision and correlation of the ammonite fauna of the Neuburg Formation (Tithonian, SW Germany). Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy), 110(1), 311–320.
Smelror, M., & Dypvik, H. (2005). Marine microplankton biostratigraphy of the Volgian–Ryazanian boundary strata, western Barents Shelf. NGU Bulletin, 443, 61–69.
Stam, B. (1986). Quantitative analysis of Middle and Late Jurassic Foraminifera from Portugal and its implications for the Grand Banks of Newfoundland. Utrecht Micropaleontology Bulletin, 34, 167.
Stricanne, L., Munnecke, A., Pross, J., & Servais, T. (2004). Acritarch distribution along an inshore–offshore transect in the Gorstian (lower Ludlow) of Gotland. Sweden. Review of Palaeobotany and Palynology, 130(1), 195–216.
Subbotina, N. N., Dutta, A. K., & Srivastava, B. N. (1960). Foraminifera from the Upper Jurassic deposits of Rajasthan (Jaisalmer) and Kutch, India. Bulletin of the Geological Mineralogical and Metallurgical Society of India, 23, 1–48.
Talib, A., Gaur, K. N., & Bhalla, S. N. (2007). Callovian-Oxfordian boundary in Kutch Mainland, India—A foraminiferal approach. Revue de Paléobiologie, 26(2), 625.
Taylor, A. M., Gowland, S., Leary, S., & Martinius, A. W. (2014). Stratigraphical correlation of the Late Jurassic Lourinhã Formation in the Consolação Sub-basin (Lusitanian Basin), Portugal. Geological Journal, 49(2), 143–162.
Thomas, J. E., & Cox, B. M. (1988). The Oxfordian-Kimmeridgian stage boundary (Upper Jurassic): dinoflagellate cyst assemblages from the Harome Borehole, north Yorkshire, England. Review of palaeobotany and palynology, 56(3–4), 313–326.
Vakhrameyev, V. A. (1982). Classopollis pollen as an indicator of Jurassic and Cretaceous climate. International Geology Review, 24(10), 1190–1196.
Wierzbowski, A., & Matyja, B. (2014). Ammonite biostratigraphy in the Polish Jura sections (central Poland) as a clue for recognition of the uniform base of the Kimmeridgian Stage. Volumina Jurassica, 12.
Wierzbowski, A., Smelror, M., & Mork, A. (2002). Ammonites and dinoflagellate cysts in the Upper Oxfordian and Kimmeridgian of the northeastern Norwegian Sea (Nordland VII offshore area): biostratigraphical and biogeographical significance. Neues Jahrbuch fur Geologie und Palaeontologie Abhandlungen, 226(2), 145–164.
Wimbledon, W. A. (2008). The Jurassic–Cretaceous boundary: an age-old correlative enigma. Episodes, 31(4), 423–428.
Acknowledgements
Fieldwork was carried out in receipt of funding by Norwegian Interactive Offshore Stratigraphic Lexicon (NORLEX). The senior author thanks Lundin Petroleum, Norway for providing a travel grant (and M. Charnock for the organization of such), the University of Portsmouth for a Placement Scheme Award, Dr S. Batenburg for Time Series Analysis training and support, G. Bell and N. Walasek for palynological training and support, and M.C. Blanc and others for a productive stay at the Applied Petroleum Technology facilities (APT), Oslo. Gratitude is extended to Dr. S. Nicoara at the Open University, UK for stable isotope and TOC measurements and Dr. D. Loydell for a useful discussion. We thank Profs. J. Ogg and D. Martill and Dr A. Waśkowska whose reviews improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: D. Marty.
Appendix
Appendix
See Figs. 5 and 6, and Table 1.

Limestone band and marlstones at the base of the Tojeira-1 section. The first sampling level is located just above the shrubbery that overhangs this part

Looking downwards on to the lower Tojeira-1 section from a goat path. In the distance, the Montejunto section outcrops along a road
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Turner, H.E., Gradstein, F.M., Gale, A.S. et al. The age of the Tojeira Formation (Late Jurassic, Early Kimmeridgian), of Montejunto, west-central Portugal. Swiss J Palaeontol 136, 287–299 (2017). https://doi.org/10.1007/s13358-017-0137-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-017-0137-6