
Abstract
Stable isotope analysis is an essential investigative technique, complementary to more traditional zooarchaeological approaches to elucidating animal keeping practices. Carbon (δ13C) and nitrogen (δ15N) stable isotope values of 132 domesticates (cattle, caprines and pigs) were evaluated to investigate one aspect of animal keeping, animal forage, at the Late Chalcolithic (mid-fourth millennium BC) site of Çamlıbel Tarlası, which is located in north-central Anatolia. The analyses indicated that all of the domesticates had diets based predominantly on C3 plants. Pig and caprine δ13C and δ15N values were found to be statistically indistinguishable. However, cattle exhibited distinctive stable isotope values and, therefore, differences in diet from both pigs and caprines at Çamlıbel Tarlası. This difference may relate to the distinct patterns of foraging behaviour exhibited by the domesticates. Alternatively, this diversity may result from the use of different grazing areas or from the foddering practices of the Çamlıbel Tarlası inhabitants.
Similar content being viewed by others


Introduction
Archaeological research in north-central Anatolia has focused predominantly on Late Bronze Age and Iron Age urban sites. By comparison, prehistoric settlement in the region has received relatively little attention (e.g. Parzinger 1993; Steadman 1995; Özdoğan 1996; Schoop 2005; Düring 2008). As a result, the animal keeping practices and human use of meat and secondary products of this region and time period are poorly understood.
Dietary stable isotope studies of animal bones offer a direct means of reconstructing at least one aspect of animal production: the type of forage domesticates had access to or were given. While such information will not be a panacea in understanding herding practices per se, it elucidates a new dimension directly relevant to animal keeping in the Anatolian region. Here carbon (δ13C) and nitrogen (δ15N) stable isotope analyses of remains from domesticated animals and humans (Pickard et al. 2016) from the Late Chalcolithic settlement site of Çamlıbel Tarlası are used to investigate animal diets. The implications of the results for stock-keeping and the consumption of meat and secondary products by a rural farming community in north-central Anatolia are explored.
Çamlıbel Tarlası—archaeological background
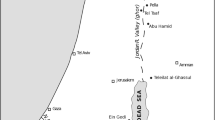
Çamlıbel Tarlası is one of only a very small number of prehistoric sites to have been investigated in north-central Anatolia. Located 2.5 km east of the modern village of Boğazkale (the location of the Late Bronze Age Hittite capital Ḫattuša), the site was excavated, as a cooperative project between the German Archaeological Institute and Edinburgh University, over three seasons from 2007 to 2009 (Schoop 2010, 2011, 2015 for summary reports). A small, short-lived settlement and mortuary site, Çamlıbel Tarlası was occupied in the mid-fourth millennium BC (Schoop et al. 2009). The site is located on a small plateau (c. 1040 m asl) in a mountainous and previously wooded region (Dörfler et al. 2000; Marsh 2010), a short distance from the main Budaközü Plain (Fig. 1).

Map of Anatolia indicating the location of Çamlıbel Tarlası and other major Neolithic and Chalcolithic settlements
Seven phases of activity (summarised in Table 1) have been identified at Çamlıbel Tarlası, all of which date to the Late Chalcolithic between 3650 and 3375 cal BC. Extractive metallurgy and small-scale agriculture are the principal activities evident at the site. The site’s occupants may have been attracted to this location because, in addition to the close proximity of an outcrop of copper ore ∼2 km to the east of the site, the surrounding plateaus would have been ideally suited to small-scale agriculture (Marsh 2010).
Çamlıbel Tarlası—the animal remains
An assemblage of 2752 identifiable animal bones (67.7 kg) was recovered at Çamlıbel Tarlası (Bartosiewicz and Gillis 2011; Bartosiewicz et al. 2013). Wild species constituted less than 1 % of the assemblage by NISP (Fig. 2). The remains were predominantly waste from food consumption. Cattle dominated the domesticate remains with pig and caprines also common. Tooth wear and epiphyseal fusion data indicate that the vast majority of the cattle were mature animals. Longevity is attributable to several factors in cattle including the slow reproduction rate and intrinsically high value of individuals as well as secondary product exploitation. Single purpose pigs were slaughtered at or before adulthood for meat. The proportion of bones from young to mature caprines was low. In addition to cattle potentially having been used for traction and sheep for wool, all bovids (including goats) were likely exploited for dairy produce. Distinction between the bones of young lambs and kids, however, tends to be unreliable, further distorting speculations concerning the ‘management’ of caprine herds (Bartosiewicz et al. 2013).

Animal taxa representation at Çamlıbel Tarlası by NISP
It has been inferred from remains of churns found at Çamlıbel Tarlası (Fig. 3) and nearby sites such as Yarıkkaya (Boğazköy) that dairy produce was important at least in the local economy (Sauter et al. 2003, Figs. 1, 2 and 3). This is congruent with the identification of animal fats, in one case likely milk fat or a derivative, on two pottery sherds from Yarıkkaya (Sauter et al. 2003).

Example of a reconstructed pottery ‘churn’ from Çamlıbel Tarlası II
Problems with proxies
Animal remains recovered at settlements such as Çamlıbel Tarlası typically represent food refuse or manufacturing waste unless interpreted as ‘ritual’ deposits of intact or articulated skeletons. However, there is a tendency to contextualize them in broader terms, inseparably mixing evidence of consumption with far less tangible aspects of exploitation, especially for secondary products. Evidence for the latter is related to animal longevity as secondary products are renewable resources from live animals (e.g. Gerritsen et al. 2010). However, interpretations regarding the bones of calves or lambs/kids alternate between showing emphasis on milk or meat ‘production’ (cf. Gourichon and Helmer 2008 and Gerritsen et al. 2010). Moreover, age profiles treated as key evidence are often based on small or unknown numbers of ageable elements encountered among the food remains.
Stable isotope analysis
Stable carbon and nitrogen isotope analysis of bone collagen has been widely demonstrated to be a useful tool in the investigation of animal diets (e.g. Pearson et al. 2007; Towers et al. 2011; Fuller et al. 2012; Gillis et al. 2013).
The technique involves the measurement of carbon (δ13C=12C/13C) and nitrogen (δ15N 14N/15N) isotope ratios in samples of bone collagen (Sealy et al. 1995). 12C/13C isotopes are incorporated into plant tissues during photosynthesis. Carbon isotope ratios (δ13C) vary between plants depending on the mechanism used to fix atmospheric carbon. Most plants fix carbon through one of two routes, either the C3 or C4 pathways (Edwards and Walker 1983). C3 plants comprise cereals such as wheat and barley, and most fruits and vegetables, while C4 plants include some tropical grasses and cereals such as millet and sorghum. Variation in plant δ13C is passed on to the tissues of animal and human consumers. Plant δ13C values may also exhibit inter-species variation within a C3 plant environment (e.g. Feranec 2007). The δ13C value of animal bone collagen can therefore be used to determine not only the relative importance of C3 vs C4 plants to diet but may also distinguish between different patterns of foraging behaviour in regions with an exclusively C3 biome (DeNiro and Epstein 1978; Feranec 2007).
Nitrogen isotope ratios (δ15N) exhibit a ‘trophic level’ effect, becoming more ‘enriched’ with each step in the food chain, i.e. they indicate an organism’s position in the food chain. The increase in δ15N values of animals over that of their diets has been observed to be in the order of 3–5 ‰ (e.g. Bocherens and Drucker 2003)—although the diet-consumer offset may vary between species and has been demonstrated to be higher than 5 ‰ in humans (Hedges and Reynard 2007; O’Connell et al. 2012). 14N/15N isotopes may be incorporated into plants from soils and/or drawn from atmospheric N2. Plants that fix nitrogen from the atmosphere (e.g. legumes) generally have lower δ15N values than those plants that fix nitrogen from soil nitrate or ammonium (DeNiro and Epstein 1981). Plant δ15N values can also vary significantly depending on water stress (e.g. Ambrose 1991), salinity (e.g. Britton et al. 2008) and increased nitrogen cycling in forested environments (e.g. van der Merwe and Medina 1991). The variability in δ15N values is passed on to the consumers of plants and may therefore indicate distinctive foraging behaviours among domesticates.
Although the fundamental principles of stable isotope analysis were established in the 1970s and have remained largely unchallenged, it is important to bear in mind that there are a number of additional issues (both dietary and non-dietary) that have implications for the reconstruction of animal forage from stable isotope values, for example the potential effects of physiological stresses, such as gestation and lactation on intra- and inter-individual isotope signatures, as well as the impact of practices such as manuring on isotopic signatures in plants and their animal consumers (e.g. Jenkins et al. 2001; Fuller et al. 2005; Bogaard et al. 2007; Nitsch et al. 2010; Olsen et al. 2014).
Materials and method
A total of 163 domesticated animal (cattle, caprine and pig) bone specimens from Çamlıbel Tarlası were measured for δ13C and δ15N to investigate animal forage and human diets (see Pickard et al. 2016) in Late Chalcolithic north-central Anatolia—see Table 2 for animal data.
A sample of approximately 1 g of bone was taken from each specimen. Pre-treatment included cleaning to remove 1–2 mm of the outer surface of the bone. Collagen was extracted using a modified version of the Longin (1971) method (Brown et al. 1988). Each sample was demineralized in 1 N HCl at 20 °C for a minimum of 24 h, rinsed three times in Milli-Q™ purified water and gelatinized in 0.03 N HCl at 80 °C for approximately 16 h. The resulting solution was then lyophilised. Samples with well-preserved collagen, i.e. collagen wt% yield of >1.00 % (Brock et al. 2010; van Klinken 1999), were measured for δ13C and δ15N by the SUERC Radiocarbon Laboratory in East Kilbride, UK, using a Thermo Scientific Delta V Advantage continuous-flow isotope ratio mass spectrometer (CF-IRMS) coupled via a Thermo Scientific ConfloIV to a Costech ECS 4010 elemental analyser (EA) fitted with a pneumatic autosampler. In-house gelatine standards, which are calibrated to the International Atomic Energy Agency (IAEA) reference materials USGS40 (l-glutamic acid, δ13CV-PDB = −26.39 ‰), USGS41 (l-glutamic acid, δ13CV-PDB = +37.63 ‰), IAEA-CH-6 (sucrose, δ13CV-PDB = −10.45 ‰), USGS25 (ammonium sulphate, δ15NAIR = −30.41 ‰), IAEA-N-1 (ammonium sulphate, δ15NAIR = +0.43 ‰) and IAEA-N-2 (ammonium sulphate, δ15NAIR = +20.41 ‰), are run in duplicate for every ten unknown samples. Results are corrected for linearity and instrumental drift and are reported as per mil (‰) relative to the internationally accepted standards V-PDB and AIR, with 1σ precisions of ±0.2 and ±0.3 ‰ for δ13C and δ15N, respectively. Collagen integrity was assessed according to the following criteria: (i) C/N ratio in the range 2.9 to 3.6 (DeNiro 1985) and (ii) minimum %C and %N as outlined by Ambrose (1990).
Less than 1 % collagen yield was obtained from 22 samples (seven caprines, 11 cattle and four pig). A further nine samples (one cattle, seven pig and one sheep) had a collagen yield of wt% >1.0 % but when measured failed to meet the criteria cited for well-preserved collagen. These samples were excluded from the discussion below.
Results
The average carbon isotope values of the domesticates (cattle, caprines and pigs) are typical of animals foraging predominantly on C3 resources.
The domestic caprine δ13C and δ15N values were found to be relatively homogeneous, with average δ13C = −19.2 ± 0.5 ‰ and δ15N = 6.4 ± 0.7 ‰ (n = 38), and are typical of herbivores consuming exclusively C3 resources (cf. Pearson et al. 2007). Little difference in the average isotope values of goat and sheep is evident (sheep average δ13C = −19.1 ± 0.5 ‰ and δ15N = 6.4 ± 0.6 ‰ (n = 23); goat average δ13C = −19.3 ± 0.8 ‰ and δ15N = 5.8 ± 0.8 ‰ (n = 4)).
The pigs sampled were identified as domestic based on their small phenotypic size in comparison to the wild progenitor and on the fact that other hunted animals were a very small component of the faunal assemblage (Bartosiewicz and Gillis 2011; Bartosiewicz et al. 2013, 111, Figs. 7 and 8). One of the pigs sampled, GUsi-2663, is a sub-adult specimen. However, the δ13C and δ15N values of this specimen overlap those obtained for the adults, i.e. no suckling enrichment effect is evident by this age. This specimen is therefore considered along with the adults. Pig mean δ13C and δ15N values suggest a relatively uniform diet, with average δ13C and δ15N of −19.3 ± 0.4 and 6.8 ± 0.6 ‰ (n = 54), respectively.
By comparison, cattle δ13C and δ15N values are more variable, with a relatively large range in both isotope values evident (see Table 2 and Fig. 4). The average values with standard deviation are δ13C = −18.7 ± 0.9 ‰ and δ15N = 7.1 ± 1.1 ‰ (n = 40). The range of cattle δ15N values is wide, spanning 4.3 ‰ with a minimum value of 5.5 ‰ and a maximum value of 9.8 ‰. The range of cattle δ13C values is also broad, from −20.1 to −17.0 ‰. The variation in cattle stable isotope values does not correspond to diachronic change (there are no statistically significant differences in the δ13C and δ15N values between bones associated with various phases of activity at Çamlıbel Tarlası).

Scatterplot of average δ13C and δ15N values of Çamlıbel Tarlası domesticate samples by species with standard deviation indicated by error bars. The average δ13C and δ15N values of the adult humans from Çamlıbel Tarlası have been included for comparison
Statistical evaluation of domesticate herbivore stable isotope values
Bartlett’s test was used to assess the null hypothesis that the variance of the δ13C values of the domesticates (cattle, caprine and pig) is homogenous. The variance of the domesticate δ13C values was found to be significantly different (Bartlett χ 2 = 25.582 with probability 0.000003, χ 2 value for significance at 5 % and d.f. 2 is >5.991). Figure 5 shows the distribution of δ13C values by taxon. The non-parametric Kruskal–Wallis test was used to assess the null hypothesis that the δ13C values of the domesticates were uniform. The δ13C values were found to be statistically different (K = 12.466179, p = 0.001963). The same statistical tests were undertaken for the domesticates with the exclusion of goats (owing to small sample size) and the caprines that could not be identified to species level. Bartlett’s test indicated that the variance of the domesticate δ13C values (i.e. cattle, pig and sheep) remained significantly different (Bartlett χ 2 = 24.763 with probability 0.000004, χ 2 value for significance at 5 % and d.f. 2 is >5.991). The Kruskal–Wallis test also indicated that there is a statistically significant difference in the δ13C values of the different domesticates (K = 12.938133, p = 0.001551, for cattle, caprine and pig; K = 12.466179, p = 0.0001963, for cattle, sheep and pig).

Boxplot showing the distribution of δ13C values of the domesticates from Çamlıbel Tarlası by taxon. The red line highlights the δ13C value −18.0 ‰, which has been used as a ‘threshold’ value for the consumption of C4 resources (e.g. Pearson et al. 2007)
The same sets of statistical analyses were repeated to compare the δ15N values of the domestic herbivores. The variances of the δ15N values were found to be unequal (Bartlett χ 2 = 9.957 with probability 0.002069, χ 2 value for significance at 5 % and d.f. 2 is >5.991, for cattle, caprines and pig; Bartlett χ 2 = 12.362 with probability 0.006885, for cattle, sheep and pig)—see Fig. 6 for distribution of δ15N values. The δ15N values of the domesticates are significantly different (Kruskal–Wallis test—K = 10.135896, p = 0.006295, for cattle, caprines and pig; K = 9.423774, p = 0.008988, for cattle, sheep and pig).

Boxplot showing the distribution of δ15N values of the domesticates from Çamlıbel Tarlası by taxon. Two cattle have statistically outlying δ15N values
A post hoc Mann-Whitney U test was used to examine pairwise differences in stable isotope values. Increased stringency was employed to compensate for multiple testing (p has significance at ≤0.05/3, i.e. 0.0167). The p values of the Mann-Whitney U test and a summary of statistical significance are presented in Tables 3, 4 and 5. Figure 7 provides a graphical representation of the isotopic relationships of the cattle, caprines and pigs for both carbon and nitrogen values based on the Mann-Whitney results.

Graphical representation of statistical significance based on Mann-Whitney U test with increase stringency. Lines between the groups represent statistically indistinguishable isotope values. Statistical equivalence of δ15N values is indicated by the green line, while that of δ13C is indicated by the red line
Discussion
Domesticate forage and animal keeping at Çamlıbel Tarlası
Statistical analysis of the carbon and nitrogen stable isotope values of the domesticates has demonstrated that the cattle have distinctive stable isotope values from both the pigs and the caprines at Çamlıbel Tarlası. There are several possible diet-related explanations that may account for these differences including (i) differences in foddering practices, (ii) differences in foraging behaviour and (iii) differences in areas grazed.
Domesticate animals often have multiple uses and may be fed distinctive diets depending on function (Bayer et al. 2003). For example, in traditional small-scale farming economies, cattle kept principally for milk may be non-grazed, fed exclusively on fodder crops to ensure high quality diet to maximise productivity (van Shaik et al. 1996). At Late Neolithic/Early Chalcolithic Ilıpınar, barley kernels and chaff were recovered in the dung of stalled cattle, interpreted by Cappers (2008, p. 120) as a diet specific to ‘beasts that are used for ploughing’ (Cappers 2008, p. 120). Although the energy requirements of non-female draught cattle have been demonstrated to be similar to that of non-draught bulls (Goe and McDowell 1980), cows used for traction show reduced fertility and milk production partially mitigated if nutritionally high quality feed is provided (Jabbar 1993). The majority of the identified cattle remains were those of mature animals, a mortality profile consistent with use for traction or dairy (Bartosiewicz and Gillis 2011). There is no archaeological evidence to support confined stalling or selective foddering of cattle at Çamlıbel Tarlası. However, the most recent settlement at Çamlıbel Tarlası was surrounded by a boundary wall, a relatively weak construction which was clearly not defensive in character. Such a wall would have been extremely useful for the protection of livestock in an environment which was potentially home to a range of large predators such as leopards, lions, bears and wolves—the latter still present today.
Domesticates have distinct ingestive and digestive capabilities (Prache et al. 1998) resulting in different behaviours in free-foraging animals. Pigs, as facultative omnivores, might be anticipated to have a distinctive diet from the domesticated ruminants. The diets of domestic pigs vary depending on husbandry practices. Pig farmers traditionally practice one of three feeding strategies, (i) free roaming and foraging; (ii) grazing in fields often with other livestock; and (iii) penning, i.e. confined to spaces such as pig sties. Free roaming pigs tend to forage over large areas and while they consume a wide range of animal foods including insects, eggs, small mammals and carrion, they are predominantly herbivorous (Studnitz et al. 2007). Penned pigs will generally be fed on plants and on domestic/household waste. If the waste products include animal proteins (e.g. milk and meat or even faeces) resulting pig δ15N values will be higher than those of free roaming or field grazed animals. However, as indicated above, the δ13C and δ15N values of the pigs at Çamlıbel Tarlası are statistically equivalent to those of the sheep as well as the extended group that includes all caprines (i.e. obligate herbivores), suggesting that the pigs were unlikely to have been fed on domestic refuse often rich in animal proteins.
Cattle and sheep, despite both being grazing ruminants, may also have distinctive foraging behaviours. Sheep tend to target higher quality forage owing to reduced gut size and shorter digestion times as well as being able to be more selective in forage owing to the shape and reduced size of the ovine dental arcade (Prache et al. 1998). Cattle tolerate lower quality and rougher forage more readily than sheep (Prache et al. 1998).
Local vegetation heterogeneity accompanied by different foraging behaviours could potentially account for the differences evident in the isotope values of cattle and sheep at Çamlıbel Tarlası. Given the range of the cattle δ13C values at Çamlıbel Tarlası, from −20.1 up to −17.0 ‰, it could be argued (cf. Pearson et al. 2007; Budd et al. 2013) that the source of this heterogeneity is the inclusion (or greater inclusion) of C4 plants in cattle diet. However, all of the crop and wild grass species identified at Çamlıbel Tarlası use the C3 photosynthetic pathway (Papadopoulou and Bogaard 2012, Table 4). Two plants recovered were not identifiable to species level, one from the borage (Boraginacaea) or sedge (Cyperaceae) families and the other from the saltbush genus (Atriplex sp.). These families include both C3 and C4 species (Edwards and Walker 1983). However, the presence of these broad taxa does not convincingly indicate the presence of C4 plant species in the Çamlıbel Tarlası locale. Furthermore, it is unlikely that C4 cereal crops such as millet (Panicum miliaceum) were available to the inhabitants of Çamlıbel Tarlası. Millet was not cultivated in central Anatolia in large quantities before at least the Bronze Age and possibly later (e.g. Nesbitt and Summers 1988; Riehl and Nesbitt 2003).
An alternative scenario to heterogeneity in local plant forage is the movement of domestic livestock between different pastures, potentially some distance from settlements and possibly with access to isotopically distinct plants (cf. Peters et al. 2013). Although C3 plants have dominated Anatolian vegetation throughout the Holocene (Rao et al. 2012), C4 plants were certainly present at least regionally. Consumption of C4 plants is clearly indicated by the range of δ13C values of wild cattle (Bos spp.) and the plant remains at the Aceramic Neolithic site of Aşıklı Höyük in the Cappadocian region of Turkey (Pearson et al. 2007). C4 plants have also been recovered from a number of sites in Anatolia and in some cases were evidently consumed by livestock. For example, at Çatalhöyük, located in south-central Anatolia, C4 grasses Aeluropus, Crypsis and Sporobolus, as well as Eleocharis (generally a C4 plant although Bruhl and Wilson (2007) describe C3/C4 intermediate and variant species), were recovered from animal dung (Richards et al. 2003; Bogaard et al. 2013a). The C4 ruderal, sun spurge (Euphorbia helioscopia), is present at Late Neolithic/Early Chalcolithic Ilıpınar, in North West Anatolia, but is not abundant (Cappers 2008).
A further possibility cannot be ruled out, that is the external, and possibly long-distance, provisioning of smaller settlements such as Çamlıbel Tarlası with at least some of their livestock or meat. Arbuckle (2012) noted increasing complexity in the management of sheep herds throughout the Chalcolithic in central Anatolia, which were at least in part distributed to settlements by specialist mobile pastoralists. Concrete evidence for far-reaching trade contacts at Çamlıbel Tarlası include finds of cockle shells (Cardiidae), which must have originated from either the Black Sea or the Mediterranean, obsidian from Cappadocia, flint blades from unknown exotic sources and a casting mould for ring-shaped figurines indicating an ideological link to the Southern Balkans (Schoop 2011; Schoop and Lehner 2013). Livestock may thus have been imported in the form of exchange or at social occasions when groups living in the broader area congregated. Meanwhile, locals may have kept small stocks. That cattle meat may have been consumed only occasionally, possibly during social gatherings or ‘feasts’, is supported indirectly by the average nitrogen stable isotope value of the inhabitants of Çamlıbel Tarlası (Pickard et al. 2016), which is similar to that of the cattle, suggesting that cattle meat was unlikely to be a significant or consistent source of dietary protein.
Domesticate forage and animal keeping across prehistoric Anatolia
The results from Çamlıbel Tarlası add to a small but growing body of isotope data from domesticates at Neolithic and Chalcolithic sites across prehistoric Anatolia.
At Çamlıbel Tarlası, the relatively low δ15N values of the pigs in comparison to ruminants suggest that they were likely free roaming and foraging. Pearson et al. (2015) also noted that the relatively low average δ15N value of pig in comparison to the human inhabitants and domesticates of Neolithic Çatalhöyük suggested that these animals were not fed on domestic waste (although three outliers with comparatively high δ15N values may be an exception). By contrast, at the PPNB site of Nevalı Çori in southeast Turkey, domestic pig and humans have similar δ15N mean values (5.5 ± 1.9 and 6.1 ± 1.0 ‰, respectively) and ranges (3.9 to 8.2 ‰ and 4.4 to 8.8 ‰, respectively) implying similar diets, i.e. at Nevalı Çori, the pigs were likely fed on household waste and thus possibly penned (Lösch et al. 2006). However, only four pig specimens were analysed at this site.
The wide ranges of δ13C and δ15N values of sheep and cattle from Çatalhöyük were attributed by Pearson et al. (2007, 2015) to the grazing of herds in different areas—in pastures with only C3 species, as well as those with both C3 and C4 plants. Extra-local grazing of at least some cattle is paralleled at Çamlıbel Tarlası. By contrast, the sheep from Çamlıbel Tarlası have relatively homogeneous δ13C and δ15N values. The differences in sheep forage and by implication stock-keeping practices between Çatalhöyük and Çamlıbel Tarlası may reflect the distinctive nature of the two sites. The scale of Çatalhöyük, a large agglomerated village or proto-urban site spanning c. 130,000 m2 (Hodder and Cessford 2004), likely necessitated extra-household herding strategies and grazing in various pastures potentially at some distance from the site (Pearson et al. 2015). In contrast, Çamlıbel Tarlası was a small-scale rural settlement covering an area of no more than 250 m2—small herds of sheep were likely managed locally.
A trend of 15N enrichment is evident in the sheep from later levels at Çatalhöyük. Budd et al. (2013) also noted an increase in δ13C and δ15N values of animal remains (n = 9) from Neolithic to Early Chalcolithic contexts at Aktopraklık, Northwest Anatolia. As both Pearson et al. (2015) and Budd et al. (2013) indicated, one possible explanation for 15N enrichment is the grazing of animals on manured fields (e.g. Fraser et al. 2011; Bogaard et al. 2013b; Styring et al. 2014). Crop manuring has been demonstrated at early Neolithic sites in southeastern Europe from c. 5900 BC (Bogaard et al. 2013b). At Çamlıbel Tarlası, the average δ15N value of all the domesticates is 6.7 ‰. Assuming a diet-herbivore offset of 4 ‰, the average δ15N value of the domesticate forage at Çamlıbel Tarlası was 2.7 ‰; this relatively low forage δ15N value suggests that these animals were not fed on manured crops/stubble (cf. Bogaard et al. 2013b). However, identifying manuring practices and/or animal penning from the stable isotope values of domesticates is non-trivial. In part, this is a consequence of the temporal and spatial variability in soil δ15N values (e.g. Ambrose 1991). Also problematic is the inclusion of 15N-depleted pulse crops in domesticate diet, which may mask the effects of consuming manured cereals or stubble. Moreover, 15N enrichment of pulse crops is evident only with intensive and protracted manuring (Fraser et al. 2011; Bogaard et al. 2013b). Pulses including bitter vetch and lentil were identified at Çamlıbel Tarlası and may have comprised a significant part of human (see Pickard et al. 2016) and/or domesticate diet.
The comparative analysis of the domesticate stable isotope datasets presented above is offered cautiously because of (i) spatial and temporal variations in environmental δ13C and δ15N values (e.g. Schwarcz et al. 1999; van Klinken et al. 1994; van Klinken et al. 2000) and the broad geographical range and chronological span of sites with stable isotope datasets in prehistoric Anatolia, (ii) the difficulty of distinguishing early domesticates from wild progenitors from morphological characteristics at Early Neolithic sites (see Zeder et al. (2006) for summary), (iii) the small datasets available from some sites and (iv) the complexities of identifying the consumption of pulses.
Conclusion
Carbon and nitrogen stable isotope analyses indicate that the domesticates from Çamlıbel Tarlası had diets based mainly on C3 resources. Of all the domesticates at Çamlıbel Tarlası, cattle exhibit the widest range of δ13C and δ15N values. Moreover, cattle have statistically distinct isotope values from both the sheep/caprines, and the pigs recovered from the site. This indicates that the cattle had distinctive foraging behaviour not only from that of the pigs (which is perhaps not surprising) but also from that of the sheep. The δ13C values of the cattle hint at the inclusion of C4 plants in diet. However, no C4 plants were identified in the plant macro assemblage recovered at the site and this suggests (if indeed C4 plants were included in the diet of cattle) that they were grazing in non-local pastures. The inter-species variability in isotope values points to distinctive and potentially complex foraging and/or feeding behaviours at Late Chalcolithic Çamlıbel Tarlası.
References
Ambrose SH (1990) Preparation and characterisation of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451. doi:10.1016/0305-4403(90)90007-R
Ambrose SH (1991) Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. J Archaeol Sci 18:293–317
Arbuckle BS (2012) Animals and inequality in Chalcolithic central Anatolia. J Anthropol Archaeol 31:302–313. doi:10.1016/j.jaa.2012.01.008
Bartosiewicz L, Gillis R (2011) Preliminary report on the animal remains from Çamlıbel Tarlası. Archeol Anz 1:76–79
Bartosiewicz L, Gillis R, Girdland-Flink L, Evin A, Cucchi T, Hoelzel R, Vidarsdottir U, Dobney K, Larson G, Schoop U-D (2013) Chalcolithic pig remains from Çamlıbel Tarlası, Central Anatolia. In: De Cupere B, Linseele V, Hamilton-Dyer S (eds) Archaeozoology of the Near East X. Proceedings of the Tenth International Symposium on the Archaeozoology of South-Western Asia and adjacent areas. Peeters, Leuven, pp. 101–120
Bayer W, von Lossau A, Feldmann A, (2003) Smallholders and community-based management of farm animal genetic resources. In Community-based management of animal genetic resources. Proceedings of the workshop held in Mbabane, Swaziland May 2001. Food and Agriculture Organization of the United Nations, Rome, 7–11
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53. doi:10.1002/oa.662
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343
Bogaard A, Charles M, Livarda A, Ergun M, Filipovic D, Jones G (2013a) The archaeobotany of mid-later occupation levels at Neolithic Çatalhöyük. In: Hodder I (ed) Humans and landscapes of Çatalhöyük: reports from the 2000–2008 seasons, monographs of the Cotsen Institute of Archaeology. University of California at Los Angeles, Los Angeles, pp. 93–128
Bogaard A, Fraser R, Heaton THE, Wallace M, Vaiglova P, Charles M, Jones G, Evershed RP, Styring AK, Andersen NH, Arbogast R-M, Bartosiewicz L, Gardeisen A, Kanstrup M, Maier U, Marinova E, Ninov L, Schäfer M, Stephan E (2013b) Crop manuring and intensive land management by Europe’s first farmers. Proc Natl Acad Sci U S A 110:12589–12594. doi:10.1073/pnas.1305918110
Britton K, Müldner G, Bell M (2008) Stable isotope evidence for salt-marsh grazing in the Bronze Age Severn Estuary, UK: implications for palaeodietary analysis at coastal sites. J Archaeol Sci 35:2111–2118. doi:10.1016/j.jas.2008.01.012
Brock F, Higham T, Bronk Ramsey C (2010) Pre-screening techniques for identification of samples suitable for radiocarbon dating of poorly preserved bones. J Archaeol Sci 37:855–865. doi:10.1016/j.jas.2009.11.015
Brown TA, Nelson DE, Southon JR (1988) Improved collagen extraction by modified Longin method. Radiocarbon 30:171–177
Bruhl JJ, Wilson KL (2007) Towards a comprehensive survey of C3 and C4 photosynthetic pathways in Cyperacea. Aliso 23:99–148
Budd C, Lillie MC, Alpaslan Roodenberg S, Karul N, Pinhasi R (2013) Stable isotope analysis of Neolithic and Chalcolithic populations from Aktopraklık, northern Anatolia. J Archaeol Sci 40:860–867. doi:10.1016/j.jas.2012.09.011
Cappers R (2008) Plant remains from the Late Neolithic and Early Chalcolithic levels. In: Roodenberg J, Alpaslan Roodenberg S (eds) Life and death in a prehistoric settlement in northwest Anatolia: the Ilıpınar excavations, Volume III. Leiden, Nederlands Instituut voor het Nabije Oosten, pp. 117–148
DeNiro MJ (1985) Postmortem preservation and alteration of in-vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317:806–809. doi:10.1038/317806a0
DeNiro MJ, Epstein S (1978) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42:495–506. doi:10.1016/0016-7037(78)90199-0
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341–351. doi:10.1016/0016-7037(81)90244-1
Dörfler W, Neef R, Pasternak R (2000) Untersuchungen zur Umweltgeschichte und Agrarökonomie im Einzugsbereich hethitischer Städte. Mitteilungen der Deutschen Orientgesellschaft 132:367–380
Düring BS (2008) The early Holocene occupation of north-central Anatolia between 10,000 and 6,000 BC cal: investigating an archaeological terra incognita. Anatol Stud 58:15–46
Edwards G, Walker DA (1983) C3, C4: mechanisms, and cellular and environmental regulation, of photosynthesis. University of California Press, Berkley
Feranec RS (2007) Stable carbon isotope values reveal evidence of resource partitioning among ungulates from modem C-3-dominated ecosystems in North America. Palaeogeogr Palaeoclimatol Palaeoecol 252:575–585
Fraser R, Bogaard A, Heaton T, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton PR, Sparkes D, Styring AK (2011) Manure and stable isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2790–2804. doi:10.1016/j.jas.2011.06.024
Fuller BT, Fuller JL, Sage NE, Harris DA, O’Connell TC, Hedges REM (2005) Nitrogen balance and δ15N: why you’re not what you eat during pregnancy. Rapid Commun Mass Spectrom 18:2889–2896. doi:10.1002/rcm.170
Fuller BT, De Cupere B, Marinova E, Van Neer W, Waelkens M, Richards MP (2012) Isotopic reconstruction of human diet and animal husbandry practices during the Classical-Hellenistic, Imperial, and Byzantine periods at Sagalassos, Turkey. Am J Phys Anthropol 149:157–171. doi:10.1002/ajpa.22100
Gerritsen F, Özbal R, Thissen L, Özbal H, Galik A (2010) The Late Chalcolithic settlement of Barcın Höyük. Anatolica 36:197–225
Gillis R, Bréhard S, Balasescu A, Vigne J-D, Popovici D, Balasse M (2013) Sophisticated cattle dairy husbandry at Borduşani-Popină (Romania, 5th mill BC): the evidence from complementary analysis of mortality profiles and stable isotopes. World Archaeology 45:447–472. doi:10.1080/00438243.2013.820652
Goe MR, McDowell RE (1980) Animal traction: guidelines for utilization. Department of Animal Science Cornell University, Ithaca, New York
Gourichon L, Helmer D (2008) Etude de la faune néolithique de Menteşe (Turquie). In: Roodenberg JJ, Alpaslan Roodenberg S (eds) Life and death in a prehistoric settlement in northwest Anatolia. The Ilıpınar excavations, volume III. Leiden, Nederlands Institut voor het Nabije Oosten, pp. 435–448
Hedges REM, Reynard LM (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251. doi:10.1016/j.jas.2006.10.015
Hodder I, Cessford C (2004) Daily practice and social memory at Çatalhöyük. Am Antiq 69:17–40. doi:10.2307/4128346
Jabbar MA (1993) Research on cow traction in Africa: some lessons to be learned. In: Lawrence PR, Lawrence K, Dijkman JT, Starkey PH (eds) Recherche pour Le Développement de la traction Animale en Afrique de L’Ouest. Proceedings of the Fourth Workshop of the West Africa Animal Traction Network held in Kano, Nigeria, 9–13 July 1990. The International Livestock Centre for Africa, Adis Ababa, Ethiopia, pp. 263–268
Jenkins SG, Partridge ST, Stephenson TR, Farley SD, Robbins CT (2001) Nitrogen and carbon isotope fractionation between mothers, neonates and nursing offspring. Oecologia 129:336–341. doi:10.1007/s004420100755
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230:241–242. doi:10.1038/230241a0
Lösch S, Grupe G, Peters J (2006) Stable isotopes and dietary adaptations in human and animals at pre-pottery Neolithic Nevalı Çori, Southeast Anatolia. Am J Phys Anthropol 131:181–193
Marsh B (2010) Geoarchaeology of the landscape at Boğazköy-Ḫattuša. Archaeol Anz 2010:201–207
Nesbitt M, Summers GD (1988) Some recent discoveries of millet (Panicum miliaceum L. and Setaria italica (L.) P. Beauv.) at excavations in Turkey and Iran. Anatol Stud 38:85–97
Nitsch EK, Humphrey LT, Hedges REM (2010) The effect of parity status on δ15N: looking for the “pregnancy effect” in 18th and nineteenth century London. J Archaeol Sci 37:3191–3199. doi:10.1016/j.jas.2010.07.019
O’Connell TC, Kneale CJ, Tasevska N, Kuhnle GGC (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149:426–434. doi:10.1002/ajpa.22140
Olsen KC, White CD, Longstaffe FJ, von Heyking K, McGlynn G, Grupe G, Rühli FJ (2014) Intraskeletal isotopic compositions (δ13C, δ15N) of bone collagen: nonpathological and pathological variation. Am J Phys Anthropol 153:598–604. doi:10.1002/ajpa.22459
Özdoğan M (1996) Pre-Bronze Age sequence of Central Anatolia: an alternative approach. In: Magen U, Rashad M (eds) Vom Halys zum Euphrat: Thomas Beran zu Ehren. Ugarit-Verlag, Münster, pp. 185–202
Papadopoulou I, Bogaard A (2012) A preliminary study of the charred macrobotanical assemblage from Çamlıbel Tarlası, North-Central Anatolia. Archaeol Anz 2011:22–27
Parzinger H (1993) Zur Zeitstellung der Büyükkaya-Ware: Bemerkungen zur vorbronzezeitlichen Kulturfolge Zentralanatoliens. Anatolica 19:211–229
Pearson JA, Buitenhuis H, Hedges REM, Martin L, Russell N, Twiss KC (2007) New light on early caprine herding strategies from isotope analysis: a case study from Neolithic Anatolia. J Archaeol Sci 34:2170–2179. doi:10.1038/230241a0
Pearson JA, Bogaard A, Charles M, Hillson SW, Larsen CS, Russell N, Twiss K (2015) Stable carbon and nitrogen isotope analysis at Neolithic Çatalhöyük: evidence for human and animal diet and their relationship to households. J Archaeol Sci 57:69–79. doi:10.1016/j.jas.2015.01.007
Peters J, Buitenhuis H, Grupe G, Schmidt K, Pöllath N (2013) The long and winding road: ungulate exploitation and domestication in Early Neolithic Anatolia (10000–7000 cal BC). In: Colledge S, Conolly J, Dobney K, Manning K, Shennan S (eds) Origins and spread of domestic animals in Southwest Asia and Europe. Left Coast Press, Walnut Creek CA, pp. 83–114
Pickard C, Schoop U-D, Dalton A, Sayle KL, Channell I, Calvey K, Thomas J-L, Bartosiewicz L, Bonsall C (2016) Diet at Late Chalcolithic Çamlıbel Tarlası, north-central Anatolia: an isotopic perspective. J Archaeol Sci: Reports 5:296–306. doi:10.1016/j.jasrep.2015.11.034
Prache S, Gordon IJ, Rook AJ (1998) Foraging behaviour and diet selection in domestic herbivores. Annales de zootechnie, INRA/EDP. Sciences 47:335–345 <hal-00889735>
Rao ZG, Chen FH, Zhang X, Xu YB, Xue Q, Zhang PY (2012) Spatial and temporal variations of C3/C4 relative abundance in global terrestrial ecosystem since the Last Glacial and its possible driving mechanism. Chin Sci Bull 57:4024–4035. doi:10.1007/s11434-012-5233-9
Richards MP, Pearson J, Molleson TI, Russell N, Martin L (2003) Stable isotope evidence of diet at Neolithic Çatalhöyük, Turkey. J Archaeol Sci 30:67–76. doi:10.1006/jasc.2001.0825
Riehl S, Nesbitt M (2003) Crops and cultivation in the Iron Age Near East: change or continuity? In Fischer B, Genz H, Köroğlu JE (eds) Identifying changes: the transition from Bronze to Iron Ages in Anatolia and its neighbouring regions. Proceedings of the International Workshop, 8–9 November Istanbul 2002. Istanbul, Türk Eskiçağ Bilimleri Enstitüsü, pp 301–312.
Sauter F, Puchinger L, Schoop U-D (2003) Fat analysis sheds light on everyday life in prehistoric Anatolia: traces of lipids identified in chalcolithic potsherds excavated near Boğazkale, Central Turkey. ARKIVOC XV:15–21
Schoop U-D (2005) Das anatolische Chalkolithikum. Eine chronologische Untersuchung zur vorbronzezeitlichen Kultursequenz im nördlichen Zentralanatolien und den angrenzenden Gebieten. Verlag Bernhard Albert Greiner, Remshalden
Schoop U-D (2010) Ausgrabungen in Çamlıbel Tarlası 2009. Archaeol Anz 2010:191–201
Schoop U-D (2011) Çamlıbel Tarlası, ein metallverarbeitender Fundplatz des vierten Jahrtausends v. Chr. im nördlichen Zentralanatolien. In: Yalçın Ü (ed) Anatolian Metal V. Deutsches Bergbaumuseum, Bochum, pp. 53–68
Schoop U-D (2015) Çamlıbel Tarlası: Late Chalcolithic settlement and economy in the Budaközü Valley (north-central Anatolia). In: Steadman SR, McMahon G (eds) The archaeology of Anatolia I. Recent discoveries (2011–2014). Cambridge Scholars Publishing, Newcastle upon Tyne, pp. 46–68
Schoop U-D, Lehner J (2013) Long-distance trade and communication networks in Late Chalcolithic Anatolia. Heritage Turkey 3:27
Schoop U-D, Grave P, Kealhofer L, Jacobsen G (2009) Radiocarbon dates from Chalcolithic Çamlıbel Tarlası. Archeol Anz 2009:66–67
Schwarcz HP, Dupras T, Fairgrieve SI (1999) 15N enrichment in the Sahara: in search of a global relationship. J Archaeol Sci 26:629–636
Sealy J, Armstrong R, Schrire C (1995) Beyond lifetime averages: tracing life histories through isotopic analysis of different calcified tissues from archaeological human skeletons. Antiquity 69:290–300
Steadman S (1995) Prehistoric interregional interaction in Anatolia and the Balkans: an overview. Bull Am Sch Oriental Res 299(300):13–32
Studnitz M, Jensen MB, Pedersen LJ (2007) Why do pigs root and in what will they root? A review on the exploratory behaviour of pigs in relation to environmental enrichment. Appl Anim Behav Sci 107:183–197. doi:10.1016/j.applanim.2006.11.013
Styring AK, Fraser RA, Bogaard A, Evershed RP (2014) The effect of manuring on cereal and pulse amino acid δ15N values. Phytochemistry 102:40–44. doi:10.1016/j.applanim.2006.11.013
Towers J, Jay M, Mainland I, Nehlich O, Montgomery J (2011) A calf for all seasons? The potential of stable isotope analysis to investigate prehistoric husbandry practices. J Archaeol Sci 38:1858–1868. doi:10.1016/j.jas.2011.03.030
van der Merwe NJ, Medina E (1991) The canopy effect, carbon isotope ratios and foodwebs in Amazonia. J Archaeol Sci 18:249–259. doi:10.1016/j.jas.2011.03.030
van Klinken GJ (1999) Bone collagen quality indicator for palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695. doi:10.1006/jasc.1998.0385
van Klinken GJ, van der Plicht H, Hedges REM (1994) Bond 13C/12C ratios reflect (palaeo-) climatic variations. Geophys Res Lett 21:445–448
van Klinken GJ, Richards MP, Hedges REM (2000) An overview of causes for stable isotopic variations in past European human populations: environmental, ecophysiological, and cultural effects. In: Ambrose SH, Katzenberg MA (eds) Biogeochemical approaches to paleodietary analysis. Springer, New York, pp. 39–63
van Shaik G, Perry BD, Mukhebi AW, Gitau GK, Djikhuizen AA (1996) An economic study of smallholder dairy farms in Murang’a District, Kenya. Prev Vet Med 29:21–36
Zeder MA, Bradley DG, Emshwiller E, Smith BD (2006) Documenting domestication. In: Zeder MA, Emshwiller E, Smith BD, Bradley DG (eds) Archaeological approaches to documenting animal domestication. University of California Press, Berkeley, pp. 1–12
Acknowledgments
This research was supported by the British Academy/Leverhulme Small Research Grants (SG121910) and the School of History, Classics and Archaeology, University of Edinburgh. The excavations at Çamlıbel Tarlası were conducted as a collaborative project by the University of Edinburgh and the German Archaeological Institute. Thanks are extended to Professor Gordon Cook (SUERC). We are also grateful to Juliet Voon, Samantha Williamson, Lisa Bird and Caroline Hall-Eastman who helped select and prepare the samples for analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Pickard, C., Schoop, UD., Bartosiewicz, L. et al. Animal keeping in Chalcolithic north-central Anatolia: what can stable isotope analysis add?. Archaeol Anthropol Sci 9, 1349–1362 (2017). https://doi.org/10.1007/s12520-016-0386-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-016-0386-0