
Abstract
The microfossil assemblages of subsurface Carboniferous rocks from Faghur-1x were examined and identified. Their biostratigraphical and paleoenvironmental importance were investigated. The assemblage included well-preserved foraminifera like Omphalotis omphalota, Omphalotis sp. 2, Omphalotis sp. 3, Paraarchaediscus stilus, Paraarchaediscus koktjubensis, Archaediscus krestovnikovi, Archaediscus complanatus, Archaediscus inflatus, Archaediscus karreri, Diplosphearina inequalis, Eotubertina sp., Tetrataxis conica, Cribrostomum lecomptei, Palaeotextularia angulata, and Palaeotextularia longiseptata. This foraminiferal association indicates the late Viséan-early Serpukhovian. The other microfossils are gastropods, brachiopods, ostracods, crinoidal ossicles, frond-like fenestrate bryozoan types and stick-like colonies, echinoderms, microproblematica like Draffania biloba and algal Calcisphaera and the dasyclad Koninckopora. This microfossils assemblage points to the deposition in a restricted to open platform in a lagoonal framework environment. The Tehenu Basin is the eastern segment of northern African Sahara basins that provided refuge for the foraminiferal genera through the mass extinction events during the glacial Viséan-Serpukhovian times. However, its foraminiferal associations have lower diversities than the western basins, which indicate that it was more readily affected by the mass extinction event.
Similar content being viewed by others

Introduction
The Paleozoic rocks in Egypt are becoming more important in the strategic planes of exploration for petroleum in the Western Desert of Egypt by the date. They are the least studied rocks among the subsurface successions in the Western Desert, where the Cretaceous and Jurassic intervals remained the central target for petroleum exploration. The Carboniferous surface rocks in Sinai and Eastern Desert were the first Paleozoic strata that had been discovered in the Egypt (Wanas 2011). In Sinai, the presence of the Paleozoic (Carboniferous) rocks has been recorded by Bauerman (1869). The knowledge about the subsurface Paleozoic rocks in Egypt generally comes from the scattered studies that outlined their geographical occurrences, lithostratigraphy and biostratigraphic content (e.g., Amin 1961; Said and Andrawis 1961; Said 1962; Soliman and El Fetouh 1970; Andrawis 1972; Barakat 1982; Abdel Sattar 1983; El-Dakkak 1988; Andrawis 1990; El Shamma et al. 1996; El Shamma et al. 1998; El Shamma et al. 2012; Wanas 2011). They were recorded in 11 wells in the Western Desert to the date of Hantar (1990), which are mostly located to the west of longitude 27° E. Mostly, these records of marine Carboniferous rocks used the micropaleontological criteria of Said and Andrawis 1961, who recovered a well-preserved Lower Carboniferous (Viséan) microfossils assemblages from the subsurface Western Desert in Faghur-1x (this study) and Mamura-1 wells, Fig. 1. These microfossil assemblages delineated the biozonation of Andrawis (1990) for the Carboniferous strata in northern Western Desert. These works used the Carboniferous foraminiferal scheme for the Paleotethyan provincialism, which are found to be correlatable with Russian and Turkish Platforms. Additionally, almost the same core intervals that were used in the study of Said and Andrawis (1961) were used by El Shamma et al. (1996), who identified significant Devonian–Carboniferous microflora from Faghur-1x and NWD 302-1 wells. Accordingly, Faghur-1x well is considered as one of the most important wells for the Paleozoic lithological and biological studies. Through these extensive studies in Faghur-1x (FRX-1) well (Said and Andrawis 1961; Keeley 1989, 1994; El Dakkak 1988; El Shamma et al. 1996; Wanas 2011), the Devonian-Carboniferous boundary was placed at the depth of 6840 ft.

On the other hand, there is a prominent evidence for the active glaciations that were especially well developed in the southern hemisphere at the Gowndwana land. The continental ice caps extended from the late Viséan to Sakmarian and reached to the latitude 35° S from the South Pole (Frakes et al. 1992; Stanley and Powell 2003). An extensive mass extinction of marine fauna accompanied the development of such ice caps especially at the late Serpukhovian. Being controlled by the glacioeustacy, the basinal migration was partially allowed during the interglacial periods (sea level rise) and the taxonomical diversity was maintained. The northwestern African Sahara basins such as Tindouf, Béchar, and Reggan represented an ecological refuge for at least foraminiferal assemblages (Cózar et al. 2014). These refuges are formed as a result of the active tectonism created partially isolated basins with stable conditions for species survive longer. The north to north western oceanic currents from the opening Palaeotethyan parallel to the coast is responsible for that longer survival of species are more common in the western basins that these are the eastern (Cózar et al. 2014). The Tehenu Basin, where the Faghur-1x well is located, is near to these basins in the eastern African Sahara.
In the present work, foraminiferal assemblages from some cores of Faghur-1x are revisited and revised. The foraminiferal biostratigraphy is used to discuss the correct Devonian-Carboniferous boundary that has been published in previously mentioned studies. The studied foraminifera were identified and microphotographed to update the records of Said and Andrawis (1961). The paleoenvironmental conditions were investigated based on these foraminiferal assemblages and other associated microfossils. These associations were compared with their coeval association from the Palaeotethyan regions in biostratigraphical and paleoenvironmental aspects. This comparison is the first opportunity to evaluate the Tehenu Basin as a segment of the ecological refuge in northwestern African basin.
Geological settings
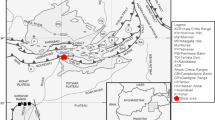
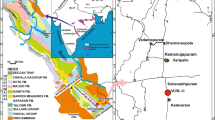
Kostandi 1959 pointed out that a fast rise of the Carboniferous Sea covered almost all of the northern Egypt and a purely marine deposition must have taken place in an area not far north of latitude 30° N in Sinai and the Eastern Desert. An expansion of the Carboniferous sea developed as a gulf along the Clysmic trend and covered the area now occupied by the Gulf of Suez and its borderland. In the north, the sea mainly deposited limestone, while in the Carboniferous gulf to the south clastics of black shales and sands were deposited, Figs. 2 and 3.

The paleogeographic map of Egypt during Carboniferous–Permian. It is adopted from Keeley 1994

In the Western Desert, the Carboniferous sediments were recorded from 11 wells west of the longitude 27° E, Figs. 2 and 3. The presence of marine Carboniferous carbonate deposits in some of the wells like Faghur-1x suggested that an open link occurred between the Carboniferous gulf and the shallow Carboniferous sea of the Libyan Desert. The carboniferous sediments exhibit lateral variation of facies from north to south, where fully marine and carbonate sediments are thicker and more common to the north and west (Hantar 1990; Klitzsch 1990). In these western areas, the strata are more or less of fully marine origin; consisting of sandstone, siltstone and shale with thin limestone intercalations, mainly at the top. This description is identical with and typical for the lithological content of Lower Carboniferous rocks of Faghur-1 well, which extend from the depth 6030 to 7780 ft (1750 ft), Fig. 4. This lithology is distinctive for the Desouqy and Dhiffah formations, which are assembled with the lower Zeiton and the upper Safi formations in the “Faghur Group” to describe the Devonian through Permo-Carboniferous subsurface rock successions in the Western Desert by Paleoservice 1986. This classification is widely applicable in the Paleozoic studies (e.g., Keeley 1989; Keeley 1994; El Shamma et al. 1996; Wanas 2011). The Desouqy Formation is laying on the Zeitoun Formation, which is only completely preserved in the Faghur-1x well. Towards the south-east, progressively earlier stages of first the Late, and then the entire Devonian are missing under Lower Carboniferous strata (Keeley 1989), Figs. 2 and 3. The lithological composition of this Desouqy Formation in Faghur-1x incorporates alluvial shoreface and delta-top sandstones thick beds of sandstone and thin layers of shales. Dhiffah Formation is overlying the Desouqy Formation conformably. In the Dhiffah Formation, there are four beds of limestone. The lower group consists of one bed and this is where the studied core materials (Core#10) were collected. The upper group consists of three beds. They are separated by few shoreface sandstones and open marine shale intercalations, Fig. 4. These two groups referred to the “Lower Limestone and Upper Limestone” by Keeley 1989, 1994. The Permo-Carboniferous Safi Formation is overlying the Dhiffah Formation.

The lithological and lithostratigraphical section of Faghur-1x well. The microfossil content is after the composite log of Pan-American (1968). 1* is the Devonian–Carboniferous boundary that used in El Dakkak (1998), El Shamma et al. (1996) and Wanas (2011). 2* is the Devonian–Carboniferous boundary that is used in Keeley 1989
The deposition of Faghur Group comprised a part of sedimentation phase II that extended through Early Tournaisian–Middle Permian (Keeley 1994). The deposition was affected primarily by eustatic changes and dominantly by the tectonics of the Calanshiyu ‘Awaynat Arch. This tectonic arc shaped the Tehenu Basin almost parallel to the Libyan-Egyptian Border (Klitzsch 1971), Figs. 2 and 3.
Material and methods
The collected materials are from the top parts of the core #10 in the Faghur-1x well, Fig. 4. Two versions of composite logs were available for Faghur-1x well (Pan-American 1959, unpublished data, 1968). The core was cut at depth interval of 6887–6928 ft and studied materials were prepared from the depths of 6887 and 6890 ft. The material were cut and polished by Carborandum and a couple of slides were prepared from each depth. The slides were then examined and microphotographed by polarized light Olympus Microscopic System. The identified taxa were arranged taxonomically according to Boudagher-Fadel (2008) in Plates 1 and 2.

1–8 Omphalotis omphalota; 1 diameter 211 μm; 2 diameter 201 μm; 3 diameter 184 μm; 4 diameter 203 μm; 5 diameter 195 μm; 6 diameter 100 μm; 7 diameter 95 μm; 8 diameter 187 μm; 9 diameter 175 μm;10 diameter 181 μm; 11, 12 Omphalotis sp. 2; 11 diameter 340 μm; 12 diameter 260 μm; 13–15 Omphalotis sp. 3; 13–17, 13 diameter 213 μm; 14 diameter 191 μm; 15 diameter 183 μm; 16 diameter 221 μm; 17 diameter 172 μm; 18 Paraarchaediscus stilus, diameter 112 μm; 19–22 Araarchaediscus koktjubensis; 19 diameter 204 μm; 20 diameter 213 μm; 21 diameter 204 μm; 22 diameter 221 μm; 23–25 Archaediscus krestovnikovi; 23 diameter 196 μm; 24 diameter 187 μm; 25 diameter 175 μm; 26 Archaediscus complanatus, diameter 217 μm; 27 Archaediscus inflatus, diameter 215 μm; 28, 29 Archaediscus karreri; 28 diameter 184 μm; 29 diameter 177 μm; 30, 31, Eotubertina sp.; 30 diameter 125 μm; 31 diameter 155 μm

1 Tetrataxis conica, diameter 185 μm; 2–6 Cribrostomum lecomptei; 2 diameter 487 μm; 3 diameter 387 μm; 4 diameter 251 μm; 5 diameter 224 μm; 6 diameter 186 μm; 7 Palaeotextularia angulata, diameter 176 μm; 8–10 Palaeotextularia longiseptata; 8 diameter 367 μm; 9 diameter 586 μm; 10 diameter 240 μm; 11 Auroria, diameter 142 μm; 12,13 Draffania biloba; 12 diameter 168 μm; 13 diameter 174 μm; 14 Corals, scale bar 500 μm; 15 Alga incerta, scale bar 500 μm; 16 Brayozoa, scale bar 500 μm; 17 A, ostracod; B, C Koninckopora sp., scale bar 1000 μm
Results and discussions
Biostratigraphy
Despite of their small size, the studied materials are unique because of two reasons. Firstly, most of the Carboniferous calcareous sediments exist only in the subsurface which is being reported by unpublished data in petroleum companies. In addition, only a little information can be obtained from surface the mainly clastic carboniferous exposures. Secondly, the samples are cores, which allowed the examination polished thin section from precise depths that cannot be obtained from the cutting samples. The microscopic examination of core materials revealed a low diversity and medium abundant foraminiferal assemblage. The individuals are well preserved, which allowed fine identification of some significant Carboniferous foraminifera. Twenty species were identified from 15 genera. However, many other forms appeared in the assemblage in inadequate positions that did not allow fine identification. The identified assemblage consists of Omphalotis omphalota, Omphalotis sp. 2, Omphalotis sp. 3, Paraarchaediscus stilus, Paraarchaediscus koktjubensis, Archaediscus krestovnikovi, Archaediscus complanatus, Archaediscus inflatus, Archaediscus karreri, Diplosphearina inequalis, Eotubertina sp., Tetrataxis conica, Cribrostomum lecomptei, Palaeotextularia angulata, Palaeotextularia longiseptata and the microproblematicum Draffania biloba, Plates 1 and 2.
The Carboniferous foraminiferal biozonation were discussed previously in Said and Andrawis 1961; Andrawis 1990. This biozonation consisted mainly of seven biozones, which are Tetrataxis conica, Endothyramopsis crassa and Archaediscus krestovnikovi for Viséan, Eostaffella postmosquensis, Ozawainella umbonata and Hemigordius simplex for Namurian and Hyperammina earlandia assemblage for Westphalian. This biozonation is correlatable with biozonation scheme from the Western Europe and Russian Platforms. The Western Europe Namurian Stage is replaced with the Russian Platform Serpukhovian in the new Geological Time Scale (Davydov et al. 2012).
The species of the genus Omphalotis (O. Omphalota, O. sp. 1 and O. sp. 2) are common in the Palaeotethyan regions of the Serpukhovian, especially in the northern basins. They are well recognized through late Viséan (Asbian-Brigantian) in Ireland, North England, Spain, Iran and Russia (Herbig and Mamet 1985; Rukina 1996; Gallagher 1998; Kalvoda 2002; Cózar and Somerville 2004; Somerville and Cózar 2005; Cózar et al. 2011, Zandkarimi et al. 2016). In the Western Africa, they remarkably flourish in the southern Palaeotethys basins like Tindouf, Béchar and Reggan through the late Viséan to Brigantian (Cózar et al. 2014). The present study is an extension of the Omphalotis occurrences in such basins to the Eastern Africa basins that were completely separated from them by African Shield in northern Saharan regions.
Cribrostomum lecomptei is most remarkable for the late Viséan–early Serpukhovian in many of the Paleotethyan basins (Pille 2008). It extends geographically from the northern regions in Ireland and Spain (Asbian-Brigantian; Gallagher 1998; Cózar 2000; Cózar and Somerville 2004; Cózar and Somerville 2005). In Western African Sahara at Tindouf Basin, it is recorded in the late Serpukhovian (Cózar et al. 2014).
Tetrataxis conica occurs for the first time in the latest Tournaisian. It is abundant in the Carboniferous Paleotethyan basins (Viséan of Ireland, Gallagher et al. 2006; Viséan-Serpukhovian of Spain, Cózar 2000; Herbig and Mamet 1985; Serpukhovian-Bashkirian of Iran, Leven et al. 2006; Zandkarimi et al. 2016; Western Arica Sahara-Cózar et al. 2014). Level of extinction undermined, probably Permian (Armstrong and Mamet 1977). Palaeotextularia longiseptata is recorded in the Viséan of the Guadiato Area in Spain (Cózar 2000).
Archaediscus krestovnikovi is essential marker for Viséan and originally described from the Late Viséan of the Russian Platform (Armstrong and Mamet 1977; Davydov et al. 2012). Nevertheless, it is recorded from Serpukhovian Palaeotethyan basins (Serpukhovian-Bashkirian of Iran, Leven et al. 2006; Serpukhovian of Turkey, Atakul-Özdemir et al. 2011).
The last occurrence Archaediscus karreri marked the Viséan/Serpukhovian boundary in the Palaeotethys basins and in the Sahara Platform (Mamet et al. 1966; Mamet 1972, Cózar and Somerville 2004). However, this species is recorded in the Tindouf Basin in Western Africa through the early Serpukhovian (Cózar et al. 2014). In addition, it is documented in the early Serpukhovian of the Rhadames Basin in western Libya (Massa and Vachard 1979). It is also traced through the late Serpukhovian and Bashkirian in the Béchar Basin (Sebbar and Lys 1989; Cózar et al. 2014).
In addition to the identified foraminifera, the microscopic examination revealed a diverse and abundant microfossil assemblage. This assemblage included a well preserved gastropods, brachiopods, ostracods, crinoidal ossicles, bryozoans frond-like fenestrate types and stick-like colonies, echinoderms, algal Calcisphaera and the dasyclad Koninckopora and the microproblematicum Draffania biloba. These contents are distinctive for the Paleozoic in general and the Carboniferous in particular. The bryozoan frond-like fenestrate types and stick-like colonies are common at the Carboniferous of Alaska (Armstrong and Mamet 1977). Calcisphaera and the dasyclad Koninckopora are markers for Viséan in Eurasia, North America, Australia, and North Africa. They are more abundant in the Paleotethyan realm (Armstrong and Mamet 1977; Cózar et al. 2008; Gallagher and Somerville 2003; Cózar et al. 2014).
The microproblematicum Draffania biloba is among recorded in the assemblages from late Viséan to early Serpukhovian in many of the Palaeotethyan region in UK and Western African Sahara in Tindouf Basin (Hallett 1971; Cózar and Somerville 2004; Cózar et al. 2014).
In the present contribution, the coexistence of the, Omphalotis spp., Cribrostomum lecomptei, Tetrataxis conica, Archaediscus karreri, Archaediscus krestovnikovi and Draffania biloba suggests a late Viséan-early Serpukhovian age for the studied core #10 in Faghur-1x well.
Paleoenvironment of deposition
The composition of the studied limestone in Core#10 in Faghur-1x well is medium-sorted, medium-grained, essentially bryozoan-crinoid-echinodermal-foraminifer packstone-grainstone with mostly sparry calcite cement. Some fragments show signs of reworking and rounding and various degrees of coating. Others show in situ accumulation. The high abundance of algal mats, calcispheres and the well-segregated bedding of clastics and carbonate sediments point to deposition in the restricted platform to open platform in a lagoonal framework environment (Armstrong and Mamet 1977). The bryozoan thickets indicate deposition in deep waters (Gallagher 1998). Tetrataxis are interpreted to have thrived at water depths below 20 m in algal-poor facies (Gallagher 1998). In addition, Draffania is a microproblematicum that characterize the low energy, deeper water, subtidal, bryozoan-rich facies that mostly lacks or with few calcareous algae (Gallagher and Somerville 2003).
These paleoenvironmental conditions in addition to the occurrence of these shallow marine beds on two levels in Dhiffah Formation above the mainly clastic Desouqy Formation indicate a full marine transgression by the end of Viséan and beginning of Serpukhovian. This marine transgression is connected to the tectonic activity in Calanshiyu’Awaynat Arch, which resulted in cyclic sea level changes (Keeley 1989; Keeley 1994) or high glacioeustacy (sea level rise) during the interglacial periods.
Tehenu Basin as segment of the western African Sahara basins
There are prominent similarities between the identified microfossil association in general and foraminiferal assemblages in particular in Faghur-1x well at the Tehenu Basin and assemblages that are identified in the western African Sahara basins (e.g., basins of Tindouf, Béchar and Reggan; Cózar et al. 2014). These similarities suggest a possible mixing of waters between these basins during the deposition. This mixing allowed better distribution of the some genera of foraminifera through the migration and permitted better surviving conditions for them to pass the mass extinction events during the extensive glaciations through the northern African Sahara basins in general. It is not possible to determine whether the foraminiferal assemblages that thrived locally in Tehenu Basin managed to pass the extinction events because of the limited sampling and difficulty to follow up the stratigraphic ranges of the species.
The diversity of the foraminiferal assemblage in the Tehenu Basin (n genera = 8) is lower than diversity of the assemblages from the western African Sahara (n genera = 10–57). This may be interpreted on the basis of that the Palaeotethyan north-northwestern warm currents from the palaeoequator did not reach effectively to the eastern basins like Tehenu Basin. Accordingly, the foraminiferal association affected more readily with glaciations and mass extinction. This assumption requires more investigations in the future on a larger sample collection.
Devonian-Carboniferous boundary in Faghour-1x well
There are two editions of the composite logs for the stratigraphy of Faghur-1x well. The older edition printed in 1958–1959 and recorded only the lithological description of the retrieved ditch cuttings and cores and the rate of drilling. In this edition, the Devonian–Lower Carboniferous boundary was placed above core# 10 at depth of 6840 ft. This edition was adopted by El Dakkak 1998; El Shamma et al. 1996; Wanas 2011. This boundary was readjusted by Mustafa 1997 and lowered to be placed above core# 11 at depth 7320 ft, depending on the palynological investigation.
The younger edition was revised by stratigraphic and biostratigraphic (using mainly foraminifera and palynology) information and was printed by Pan American United Arab Oil Company under the supervision of J.P Beckmann in November 1968. In this version, the Devonian–Lower Carboniferous boundary was placed above core #15 at depth of 7780 ft. This edition was accepted in Keeley 1989.
The findings of Carboniferous foraminifera in core #10 in the present contributions keep up the placing of the Devonian-Carboniferous beneath this core. Nevertheless, they do not support any of the reports and studies mentioned above; though, it coincides with results of Mustafa (1997).
Conclusions
The study of subsurface sample materials from the limestone beds of Core#10 in Faghur-1x well lead to identification of a well preserved microfossil assemblage. The foraminifera association is composed of Omphalotis omphalota, Omphalotis sp. 2, Omphalotis sp. 3, Paraarchaediscus stilus, Paraarchaediscus koktjubensis, Archaediscus krestovnikovi, Archaediscus complanatus, Archaediscus inflatus, Archaediscus karreri, Diplosphearina inequalis, Eotubertina sp., Tetrataxis conica, Cribrostomum sp., Palaeotextularia angulata, Palaeotextularia longiseptata and microproblematicum Draffania biloba. This assemblage is of late Viséan-early Serpukhovian age. Accordingly, this division of the Faghur-1x well sediments should be Carboniferous. The other microfossil assemblage consisted mainly of gastropods, brachiopods, ostracods, crinoidal ossicles, bryozoans frond-like fenestrate types and stick-like colonies, echinoderms and algal Calcisphaera and the dasyclad Koninckopora. This microfossil assemblage indicates deposition in the deep (> 20 m) restricted to open platform in a lagoonal framework environment.
The foraminiferal associations the Tehenu Basin indicate that it is a segment of the northern African Sahara Palaeotethyan basins like Tindouf, Béchar and Reggan. Although it has a lower foraminiferal diversity in comparison of the western basin that indicates that the warm north-northwestern Palaeotethyan currents were not effective in the Tehenu Basin.
The Devonian-Carboniferous boundary is placed beneath the core #10 according to the present investigations. This is coinciding with results from the microfloral biozonation of Mustafa (1997).
References
Abdel Sattar G (1983) Biostratigraphy of Paleozoic rocks in north Egypt. PhD. Thesis, Ain Shams Univ
Amin MS (1961) Subsurface features and oil prospects of the Western Desert. Egypt. 3rd Arab petroleum congress. Alexandria 8pp
Andrawis SF (1972) New biostratigraphic contribution for the Paleozoic rocks of Gibb Aria Well No. 2, Western Desert, Egypt. 8th Arab Petroleum Congress, Algiers
Andrawis SF (1990) Tables of foraminiferal biozones. In: Said R (ed.) The Geology of Egypt Balkema, Rotterdam: 639–648
Armstrong AK, Mamet BL (1977) Carboniferous microfacies, microfossils, and corals, Lisburne Group, Arctic Alaska. Geological Survey Professional Paper 849, 238pp
Atakul-Özdemir A, Altıner D, Özkan-Altıner S, Yılmaz İÖ (2011) Foraminiferal biostratigraphy and sequence stratigraphy across the mid-Carboniferous boundary in the Central Taurides, Turkey. Facies 57:705–730
Barakat MG (1982) General review of the petroliferous provinces of Egypt. Petroleum and natural gas project, Cairo Univ./M.I.T. Technology Planning Program
Bauerman H (1869) Note on geological reconnaissance made in Arabia Petraea in Spring of 1869. Quart J Geol Soc London 25:17–38
Boudagher-Fadel Mk (2008) Evolution and geological significance of larger benthic foraminifera. Dev Palaeontol Stratigraphy 21:570
Cózar P (2000) Mississippian Tetrataxidae, Palaeotextulariidae and Biseriamminidae (Foraminiferida) from Guadiato area (Córdoba, Spain). Coloquios de Paleontología 51:35–68
Cózar P, Somerville ID (2004) New algal and foraminiferal assemblages and evidence for recognition of the Asbian-Brigantian boundary in northern England. Proc Yorks Geol Soc 55(1):43–65
Cózar P, Somerville ID (2005) Stratigraphy of upper Visean carbonate platform rocks in the Carlow area, southeast Ireland. Geol J 40(1):35–64
Cózar P, Vachard D, Somerville ID, Berkhli M, Medina-Varea P, Rodríguez S, Said I (2008) Late Viséan–Serpukhovian foraminiferans and calcareous algae from the Adarouch region (central Morocco), North Africa. Geol J 43:463–485
Cózar P, Said I, Somerville ID, Vachard D, Medina-Varea P, Rodríguez S, Berkhli M (2011) Potential foraminiferal markers for the Visean–Serpukhovian and Serpukhovian–Bashkirian boundaries—a case-study from Central Morocco. J Paleontol 85:1105–1127
Cózar P, Vachard D, Somerville ID, Medina-Varea P, Rodríguez S, Said I (2014) The Tindouf Basin, a marine refuge during the Serpukhovian (Carboniferous) mass extinction in the northwestern Gondwana platform. Palaeogeogr Palaeoclimatol Palaeoecol 394:12–28
Davydov VI, Korn D, Schmitz MD, (2012) The carboniferous period. In Gradstein FM, Ogg JG, Schmitz MD, Ogg G (eds.). Geological Time Scale 2012. Elsevier BV 2:603–651
El Shamma AA, Abdel Malik WM, Baioumi AA, Moustafa TF (1996) Microfloral characteristics of some Carboniferous rocks in the Western Desert, Egypt. Bull Fac Sci Assiut Univ 25(2–F):107–124
El Shamma AA, Mostafa TF, Abdel Malik WM (1998) Devonian spores from subsurface rocks in the Western Desert, Egypt. Proceedings of the 14th Petroleum Conference, EGPC, Cairo 1:451–465
El Shamma AA, Moustafa TF, Hosny AM (2012) A new study of acritarchs and chitinozoa of established subsurface Devonian rocks at north western Desert, Egypt. J Appl Sci Res 8(4):1901–1917
El-Dakkak MW (1988) Geological studies of subsurface Paleozoic strata of northern Western Desert, Egypt. J Afr Earth Sci 7:103–111
Frakes LA, Francis JE, Syktu JI (1992) Climate modes of the Phanerozoic. University Press, Cambridge, p 274
Gallagher SJ (1998) Controls on the distribution of calcareous Foraminifera in the Lower Carboniferous of Ireland. Mar Micropaleontol 34(3):187–211
Gallagher SJ, Somerville ID (2003) Lower carboniferous (late Vise’an) platform development and cyclicity in southern Ireland: foraminiferal biofacies and lithofacies evidence. Riv Ital Paleontol Stratigr 109(2):159–171
Gallagher SJ, MacDermot CV, Somerville ID, Pracht M, Sleeman AG (2006) Biostratigraphy, microfacies and depositional environments of upper Viséan limestones from the Burren region, County Clare, Ireland. Geological Journal 41(1):61–91
Hallett D (1971) Foraminifera and algae from the Yoredale “series” (Viséan–Namurian) of northern England. Compte Rendu 6 Congrès International de Stratigraphie du Carbonifère, Sheffield 1967, 3: 873–901
Hantar G (1990) Geology of selected areas: north Western Desert. In: Said R (ed) The geology of Egypt. Balkema, Rotterdam, pp 293–320
Herbig HG, Mamet B (1985) Stratigraphy of the limestone boulders, Marbella Formation (Betic Cordillera, Southern Spain). CR 10eme Congr. Intern. Strat. Géol. Carbonifere, 1: 199–212
Kalvoda J (2002) Late Devonian–Early Carboniferous foraminiferal fauna: zonations, evolutionary events, paleobiogeography and tectonic implications. Folia Facultatis Scientiarium Naturalium Universitatis Masarykianae Brunensis, Geologia 30:1–213
Keeley ML (1989) The Palaeozoic hisory of the Western Desert of Egypt. Basin Res 2:35–48
Keeley ML (1994) Phanerozoic evolution of the basins of northern Egypt and adjacent areas. Geol Rundsch 83:728–742
Klitzsch E (1971) The structural development of parts of North Africa since Cambrian time. In: Gray C (ed) The geology of Libya. Faculty of Science, Tripoli, pp 255–261
Klitzsch E (1990) The Paleozoic. In: Said R (ed) The geology of Egypt. Balkema, Rotterdam, pp 393–406
Kostandi AB (1959) Facies maps for the study of the Paleozoic and Mesozoic sedimentary region. In: 1st Arab Petroleum Congress, Cairo 2: 54–62
Leven EJ, Davydov VI, Gorgij MN (2006) Pennsylvanian Stratigraphy and Fusulinids of Central and Eastern Iran. Palaeontologia Electronica 9(1):1A 36p
Mamet BL (1972) Considérations paléogéographiques déduites de l'étude des Foraminifères des couches de passage du Viséen au Namurien (bassins de Reggane et Fort-Polignac, Sahara central). Bull Inst R Sci Nat Belg 48:1–13
Mamet BL, Choubert G, Hottinger G (1966) Notes sur le Carbonifère du Jebel Ouarkziz. Étude du passage du Viséen au Namurien d'après les foraminifères. Notes Serv Géol Maroc 27(198):7–21
Massa D, Vachard D (1979) Le Carbonifère de Libye Occidentale: biostratigraphie et micropaléontologie; position dans le domaine téthysien d'Afrique du Nord. Rev Inst Fr Pétrol 34:3–65
Mustafa, T.F. 1997. Palynostratigraphic study of the Devonian rocks of the Western Desert Egypt. Unpublished PhD thesis Ain Shams University., p. 160
Paleoservices Ltd (1986) Hydrocarbon potential of the Paleozoic rocks of the Western Desert, Egypt. Unpublished report
Pan-American (1968) Revised stratigraphic data. Unpublished Composite log
Pille L (2008) Foraminifères et algues calcaires du Missississippien supérieur (Viséen supérieur-Serpukhovien): rôles biostratigraphique, paléoécologique et paléogéographique aux échelles locale, régionale et mondiale. Ph. D.Diss
Rukina GA (1996) Sequence biostratigraphy of the Tournaisian-Lower Viséan rocks of the Russian platform. Geol Soc Lond, Spec Publ 107(1):365–369
Said R (1962) Geology of Egypt. Elsevier, Amsterdam, p 377
Said R, Andrawis SF (1961) Lower carboniferous microfossils from the subsurface rock of the Western Desert of Egypt. Contrib Cushman Found Foraminifer Res 12(1):22–25
Sebbar A, Lys M (1989) Biostratigraphie du Carbonifère inférieur: Serpukhovien du Djebel Arlal, Bassin de Béchar, Algérie. Rev Micropaleontol 32:53–62
Soliman SM, El Fetouh MA (1970) Carboniferous of Egypt, isopach and lithifacies maps. Bull AAPG 54:1918–1930
Somerville ID, Cózar P (2005) Late Asbian to Brigantian (Mississippian) foraminifera from southeast Ireland: comparison with northern England assemblages. J Micropalaeontol 24(2):131–144
Stanley SM, Powell MG (2003) Depressed rates of origination and extinction during the late Paleozoic ice age: a new state for globalmarine ecosystem. Geology 31:877–880
Wanas HA (2011) The Lower Paleozoic rock units in Egypt: an overview. Geosci Front 2(4):491–507
Zandkarimi K, Najafian B, Vachard D, Bahrammanesh M, Vaziri SH (2016) Latest Tournaisian–late Viséan foraminiferal biozonation (MFZ8–MFZ14) of the Valiabad area, northwestern Alborz (Iran): geological implications. Geol J 51(1):125–142
Acknowledgements
The authors express their gratitude to the Egyptian General Petroleum Corporation EGPC, for providing the samples and well logs for this study. The authors are grateful for the Egyptian Petroleum Research Institute (EPRI) for supporting all the facilities used in this study. We appreciate the help of Mr. Moaaz Adham in reviewing the English language. Thanks are also due to Prof. Dr. Mohamed El Boukhary and an anonymous reviewer for their scientific comments and valuable recommendations on the initial version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Makled, W.A., Abdo, A.A. The biostratigraphic and paleoecological importance of the subsurface Carboniferous microfossil assemblages from Faghur-1x well, Western Desert of Egypt. Arab J Geosci 10, 389 (2017). https://doi.org/10.1007/s12517-017-3181-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-017-3181-z