
Abstract
The ability of human embryonic stem cells (hESCs) to proliferate indefinitely is attributed to its high telomerase activity and associated long telomere. However, factors regulating telomere length in hESCs remain largely uncharacterized. The aims of this study were, to identify factors which modulate telomere length of hESCs, and to determine if the telomere length of hESCs influences cellular senescence of its differentiated progeny cells. Telomerase reverse transcriptase (TERT) gene expression, telomerase activity and telomere length of hESCs cultured in different culture systems were compared. Genetically identical hESCs of different telomere lengths were differentiated into fibroblasts simultaneously, and the population doubling and cellular senescence levels were determined. We found that telomere lengths were significantly different in different culture systems and Fibroblast growth factor-2 (FGF-2) upregulated TERT expression, telomerase activity and telomere length via Wnt/β-catenin signaling pathway in hESCs in a significant manner. We also provide evidence that fibroblast differentiated from hESCs with longer telomere exhibited significant more population doublings and longer life span than those derived from hESCs with shorter telomeres. Thus, FGF-2 levels in hESCs culture systems can be manipulated to generate cells with longer telomere which would be advantageous in the applications of hESCs in regenerative medicine.






Similar content being viewed by others
References
Allsopp RC et al (1992) Telomere length predicts replicative capacity of human fibroblasts. Proc Natl Acad Sci USA 89:10114–10118
Amit M et al (2000) Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Dev Biol 227:271–278. doi:10.1006/dbio.2000.9912
Armstrong L et al (2005) Overexpression of telomerase confers growth advantage, stress resistance, and enhanced differentiation of ESCs toward the hematopoietic lineage. Stem Cells 23:516–529. doi:10.1634/stemcells.2004-0269
Chen S, Choo A, Chin A, Oh SK (2006) TGF-beta2 allows pluripotent human embryonic stem cell proliferation on E6/E7 immortalized mouse embryonic fibroblasts. J Biotechnol 122:341–361. doi:10.1016/j.jbiotec.2005.11.022
Choo A, Padmanabhan J, Chin A, Fong WJ, Oh SK (2006) Immortalized feeders for the scale-up of human embryonic stem cells in feeder and feeder-free conditions. J Biotechnol 122:130–141. doi:10.1016/j.jbiotec.2005.09.008
Counter CM, Avilion AA, LeFeuvre CE, Stewart NG, Greider CW, Harley CB, Bacchetti S (1992) Telomere shortening associated with chromosome instability is arrested in immortal cells which express telomerase activity. EMBO J 11:1921–1929
Debacq-Chainiaux F, Erusalimsky JD, Campisi J, Toussaint O (2009) Protocols to detect senescence-associated beta-galactosidase (SA-betagal) activity, a biomarker of senescent cells in culture and in vivo. Nat Protoc 4:1798–1806. doi:10.1038/nprot.2009.191
Dimri GP et al (1995) A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci USA 92:9363–9367
Ding V, Choo AB, Oh SK (2006) Deciphering the importance of three key media components in human embryonic stem cell cultures. Biotechnol Lett 28:491–495. doi:10.1007/s10529-006-0005-8
Ding VM, Ling L, Natarajan S, Yap MG, Cool SM, Choo AB (2010) FGF-2 modulates Wnt signaling in undifferentiated hESC and iPS cells through activated PI3-K/GSK3beta signaling. J Cell Physiol 225:417–428. doi:10.1002/jcp.22214
Eiselleova L et al (2009) A complex role for FGF-2 in self-renewal, survival, and adhesion of human embryonic stem cells. Stem Cells 27:1847–1857. doi:10.1002/stem.128
Feng J et al (1995) The RNA component of human telomerase. Science 269:1236–1241
Forsyth NR, McWhir J (2008) Human embryonic stem cell telomere length impacts directly on clonal progenitor isolation frequency. Rejuvenation Res 11:5–17. doi:10.1089/rej.2007.0567
Fu X, Toh WS, Liu H, Lu K, Li M, Cao T (2011) Establishment of clinically compliant human embryonic stem cells in an autologous feeder-free system. Tissue Eng Part C 17:927–937. doi:10.1089/ten.TEC.2010.0735
Greider CW, Blackburn EH (1987) The telomere terminal transferase of Tetrahymena is a ribonucleoprotein enzyme with two kinds of primer specificity. Cell 51:887–898
Hande MP, Samper E, Lansdorp P, Blasco MA (1999) Telomere length dynamics and chromosomal instability in cells derived from telomerase null mice. J Cell Biol 144:589–601
Hoffmeyer K et al (2012) Wnt/beta-catenin signaling regulates telomerase in stem cells and cancer cells. Science 336:1549–1554. doi:10.1126/science.1218370
James D, Levine AJ, Besser D, Hemmati-Brivanlou A (2005) TGFbeta/activin/nodal signaling is necessary for the maintenance of pluripotency in human embryonic stem cells. Development 132:1273–1282. doi:10.1242/dev.01706
Katoh M, Katoh M (2006) Cross-talk of WNT and FGF signaling pathways at GSK3beta to regulate beta-catenin and SNAIL signaling cascades. Cancer Biol Ther 5:1059–1064
Katoh M, Katoh M (2007) WNT signaling pathway and stem cell signaling network. Clin Cancer Res 13:4042–4045. doi:10.1158/1078-0432.CCR-06-2316
Kurz DJ et al (2003) Fibroblast growth factor-2, but not vascular endothelial growth factor, upregulates telomerase activity in human endothelial cells. Arterioscler Thromb Vasc Biol 23:748–754. doi:10.1161/01.ATV.0000069624.55424.61
Levenstein ME, Ludwig TE, Xu RH, Llanas RA, VanDenHeuvel-Kramer K, Manning D, Thomson JA (2006) Basic fibroblast growth factor support of human embryonic stem cell self-renewal. Stem Cells 24:568–574. doi:10.1634/stemcells.2005-0247
Levy MZ, Allsopp RC, Futcher AB, Greider CW, Harley CB (1992) Telomere end-replication problem and cell aging. J Mol Biol 225:951–960
Lin J et al (2016) Systematic and cell type-specific telomere length changes in subsets of lymphocytes. J Immunol Res 2016:5371050. doi:10.1155/2016/5371050
Liu X, Zheng SX, Zhou LJ, Zhu XZ (2000) Basic fibroblast growth factor up-regulates the expression of vascular endothelial growth factor in primary cultured rat astrocytes. Acta Pharmacol Sin 21:19–22
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25:402–408. doi:10.1006/meth.2001.1262
Ludwig TE, Bergendahl V, Levenstein ME, Yu J, Probasco MD, Thomson JA (2006) Feeder-independent culture of human embryonic stem cells. Nat Methods 3:637–646. doi:10.1038/nmeth902
Rochette PJ, Brash DE (2010) Human telomeres are hypersensitive to UV-induced DNA Damage and refractory to repair. PLoS Genet 6:e1000926. doi:10.1371/journal.pgen.1000926
Sakaki-Yumoto M, Liu J, Ramalho-Santos M, Yoshida N, Derynck R (2013) Smad2 is essential for maintenance of the human and mouse primed pluripotent stem cell state. J Biol Chem 288:18546–18560. doi:10.1074/jbc.M112.446591
Sato N, Meijer L, Skaltsounis L, Greengard P, Brivanlou AH (2004) Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat Med 10:55–63. doi:10.1038/nm979
Stambolic V, Ruel L, Woodgett JR (1996) Lithium inhibits glycogen synthase kinase-3 activity and mimics wingless signalling in intact cells. Curr Biol 6:1664–1668
Takao Y, Yokota T, Koide H (2007) Beta-catenin up-regulates Nanog expression through interaction with Oct-3/4 in embryonic stem cells. Biochem Biophys Res Commun 353:699–705. doi:10.1016/j.bbrc.2006.12.072
Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM (1998) Embryonic stem cell lines derived from human blastocysts. Science 282:1145–1147
Turner S, Wong HP, Rai J, Hartshorne GM (2010) Telomere lengths in human oocytes, cleavage stage embryos and blastocysts. Mol Hum Reprod 16:685–694. doi:10.1093/molehr/gaq048
Vallier L, Alexander M, Pedersen RA (2005) Activin/Nodal and FGF pathways cooperate to maintain pluripotency of human embryonic stem cells. J Cell Sci 118:4495–4509
van Noort M, Meeldijk J, van der Zee R, Destree O, Clevers H (2002) Wnt signaling controls the phosphorylation status of beta-catenin. J Biol Chem 277:17901–17905. doi:10.1074/jbc.M111635200
von Zglinicki T (2000) Role of oxidative stress in telomere length regulation and replicative senescence. Ann N Y Acad Sci 908:99–110
Wright WE, Pereira-Smith OM, Shay JW (1989) Reversible cellular senescence: implications for immortalization of normal human diploid fibroblasts. Mol Cell Biol 9:3088–3092
Xu C, Inokuma MS, Denham J, Golds K, Kundu P, Gold JD, Carpenter MK (2001) Feeder-free growth of undifferentiated human embryonic stem cells. Nat Biotechnol 19:971–974. doi:10.1038/nbt1001-971
Xu RH, Peck RM, Li DS, Feng X, Ludwig T, Thomson JA (2005) Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nat Methods 2:185–190. doi:10.1038/nmeth744
Yamamoto H, Sakane H, Yamamoto H, Michiue T, Kikuchi A (2008) Wnt3a and Dkk1 regulate distinct internalization pathways of LRP6 to tune the activation of beta-catenin signaling. Dev Cell 15:37–48. doi:10.1016/j.devcel.2008.04.015
Zeng X (2007) Human embryonic stem cells: mechanisms to escape replicative senescence? Stem Cell Rev 3:270–279. doi:10.1007/s12015-007-9005-x
Zeng S et al (2014) Telomerase-mediated telomere elongation from human blastocysts to embryonic stem cells. J Cell Sci 127:752–762. doi:10.1242/jcs.131433
Zhang Y, Toh L, Lau P, Wang X (2012) Human telomerase reverse transcriptase (hTERT) is a novel target of the Wnt/beta-catenin pathway in human cancer. J Biol Chem 287:32494–32511. doi:10.1074/jbc.M112.368282
Acknowledgments
This research was supported by grant from National University Health System (R221000085733) and National University of Singapore (R221000050133).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no financial or commercial conflict of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
10522_2016_9662_MOESM1_ESM.tif
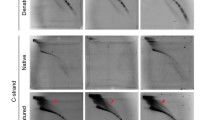
Fig. S1a Southern blot image of TRF assay of H1 hESCs cultured in MEF systems at different passages. The band on top of the second and third lane is contaminated telomere of MEF feeder. b Southern blot image of TRF assay of H1 hESCs cultured in mTeSR system at different passages. Yellow lines indicate average telomere length of respective samples. c Southern blot image of TRF assay of MEF feeder cells only (Lane 5) and hESCs cultured in mTeSR system (Lane 6) and hESCs cultured in MEF feeder system (Lane 2, 3 and 4)
10522_2016_9662_MOESM2_ESM.tif
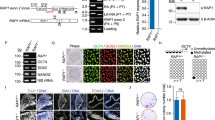
Fig. S2a Relative gene expression of TERT after H1 hESCs were treated 24 h with and without TGFβ (0.6 ng/mL) in CM system. b Relative gene expression of TERT after H1 hESCs were treated 24 h with or without LiCl (1 and 5 mM) in CM system. Data were presented as mean ± SEM and results were generated from three independent experiments, n.s. not significant. Compared with non-treated H1 hESCs. c Western blot repeats of active β-catenin and total β-catenin in H1 hESCs were treated with either FGF-2 (100 ng/mL), DKK1 (50 ng/mL) + FGF-2 (100 ng/mL), DKK1 (50 ng/mL), DKK1 (50 ng/mL) + Wnt3a (100 ng/mL) or Wnt3a (100 ng/mL) for 24 h in CM system. d Western blot of active β-catenin and total β-catenin in H1 hESCs treated with different level of FGF-2. e Relative quantification of above western blot. f Relative TA of H1 hESCs treated with FGF-2 of different concentration. Data were presented as mean ± SEM and results were generated from three independent experiments, n.s. not significant. Compared with non-treated H1 hESCs
10522_2016_9662_MOESM3_ESM.tif
Fig. S3a Relative gene expression of TERT of long-ebFs and short-ebFs at the indicated passage numbers. b Relative gene expression of the fibroblast marker P4Hβ of ebFs with long- and short telomeres at the indicated passage numbers. Data were presented as mean ± SEM and results were generated from three independent experiments, Asterisk p < 0.05, compared with undifferentiated H1 hESCs. n.s. not significant. c Relative gene expression of p21 of ebFs with long- and short-telomere at the indicated passage number. Data were presented as mean ± SEM and results were generated from three independent experiments, Asterisk p < 0.05, compared with undifferentiated H1 hESCs. n.s. not significant
Rights and permissions
About this article

Cite this article
Zou, Y., Tong, H.J., Li, M. et al. Telomere length is regulated by FGF-2 in human embryonic stem cells and affects the life span of its differentiated progenies. Biogerontology 18, 69–84 (2017). https://doi.org/10.1007/s10522-016-9662-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10522-016-9662-8