
Abstract
Silvopastoral systems with grazing pigs are mainly concentrated in Mediterranean regions, for instance in Iberian dehesas and montados. Conversely, in European deciduous forests, outdoor pig farming has almost disappeared, despite it could provide several environmental and socioeconomic benefits. For instance, pig grazing could be used for habitat restoration and conservation, while providing high quality meat products and promoting local breeds. In this study, we investigated (i) the plant species selection and (ii) the impact on tree resprouts (after a silvicultural cut) by semi-free ranging pigs in a degraded oak temperate forest in Northwestern Italy. We monitored pigs foraging behavior by direct observations and discriminated the plant species into three groups: preferred, indifferently consumed and avoided plants. The impact on the resprouts of six tree species was assessed by modeling the percentage of damaged shoots and their height through time using Generalized Linear and Additive Mixed Models. Swine expressed a selective foraging behavior, which was likely influenced by the forage quality and toxicity of plant species. The leaves and resprouts of Corylus avellana and Castanea sativa were highly consumed, whereas Acer and Quercus species and Fraxinus ornus were avoided. Mature leaves of Robinia pseudoacacia were highly preferred while its young resprouts were only slightly impacted, suggesting that pig grazing may not be effective to control this alien invasive species. Contrarily, pigs consumed Rubus sp. abundantly, suggesting they could be used to clear the understory in degraded forests.
Similar content being viewed by others


Introduction
Silvopastoralism has a long tradition in Europe, being practiced in the past in all the countries and biogeographic regions, from boreal forests to Mediterranean evergreen oak wood-pastures (Rigueiro-Rodríguez et al. 2009). With the intensification of agriculture in the mid-twentieth century, silvopastoral systems declined in many regions and nowadays they are mainly concentrated in the Mediterranean area (Mosquera-Losada et al. 2016). However, during the last 20 years, a novel interest has grown in this multifunctional system, due to the numerous ecosystem services that it provides to society (Moreno et al. 2018). For instance, there is evidence that forest grazing by livestock can enhance plant biodiversity, control shrub encroachment, and counteract the invasion of alien species (Mayle 1999; McEvoy et al. 2006; Öllerer et al. 2019). Moreover, it has also proven to be an efficient and cost-effective fire prevention tool (see for instance ‘RAPCA’, the network of grazed fuel breaks in Andalucia; Varela et al. 2018). As a consequence, the European Union has recognized the importance of silvopastoralism for the sustainability of farming systems, supporting it with the Common Agricultural Policy measures (Mosquera-Losada et al. 2016).
Among present silvopastoral systems, those based on pigs are mainly located in Mediterranean oak forests and wood-pastures. For instance, the farming of Iberian pigs in the Spanish dehesas and Portuguese montados is an excellent example of a multifunctional agroforestry system, whose value is well-recognized by the consumers of the derived animal products (Silva and Nunes 2013). Conversely, in temperate deciduous forests, outdoor pig farming has almost disappeared (Jørgensen 2013; Molnár et al. 2021), despite it could provide multiple environmental and socioeconomic benefits in these habitats as well: pig grazing might be used for habitat conservation and restoration in degraded and abandoned forests (Mayle 1999; Molnár et al. 2021); pig farming systems can support the conservation of local breeds, which are more suitable for forest farming than the commercial ones (Miao et al. 2004); last but not least, meat products could be highly valued on the market due to their quality (Silva and Nunes 2013) and could respond to the increasing demand for higher animal welfare standards (Brownlow et al. 2005).
Despite the potential positive effects of pig grazing in the deciduous forests of Europe, their foraging behavior in these environments has been poorly documented (but see Molnár et al. 2021), while forest sustainable management and habitat conservation actions would profit from this knowledge. Indeed, the selection of plant species by livestock has direct consequences on vegetation composition and structure (e.g., Pittarello et al. 2017; Pauler et al. 2020). Moreover, when silvicultural practices are applied to woodlands (e.g., in coppice systems), the knowledge on how pig browsing affects tree regeneration would be relevant.
In this context, pig grazing was introduced in a degraded oak (Quercus robur L.) stand in Northwestern Italy, where nowadays this type of management is not a common practice, as part of the pilot project ‘Food for Forest’ (Rural Development Programme—RDP, measure 16.2 of Piedmont Region). The aim of the project was to test whether pig grazing could be suitable for the restoration and sustainable management of the forest. The forest was degraded due to the abandonment of silvicultural management, which led to the worsening of the tree composition (reduction of valuable autochthonous tree species, such as oaks) and structure (reduction of large adult trees), the invasion by undesirable and alien plant species, and the reduction of its economic value. Specifically, pig grazing was targeted to reduce the abundance of undesirable shrubs in the understory, such as bramble (Rubus sp.), and of alien plant species, such as Robinia pseudoacacia L. Bramble tends to expand in degraded woodlands, causing a reduction of biodiversity (McEvoy et al. 2006; Harmer et al. 2016). Moreover, it severely limits silvicultural interventions due to the expensive mechanical clearance needed to remove it. Robinia pseudoacacia is a N-fixing legume which alters soil characteristics and affects biodiversity and plant community composition in native forest ecosystems (Vítková et al. 2017). Besides the introduction of pig grazing, the pilot project ‘Food for Forest’ also included silvicultural interventions to improve the composition and structure of the forest tree layer.
In the present study, we aimed at assessing the foraging behavior of semi-free ranging pigs in a degraded temperate forest. Particularly, we investigated (a) the plant species selection of swine grazing in the forest understory and (b) the impact of browsing on the resprouts of six tree species (Acer pseudoplatanus L., Castanea sativa Mill., Corylus avellana L., Fraxinus ornus L., Prunus avium L., and R. pseudoacacia) after silvicultural cuts.
Methods
Study area
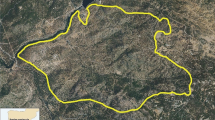
The study was carried out in a hill woodland stand in Piedmont Region, Italy (45° 19′ 59.2″ N 7° 53′ 52.3″ E; Fig. 1a). The area was included in Natura 2000 network (Special Area of Conservation IT1110009 ‘Bosco del Vaj e Bosc Grand’). The elevation was 500 m a.s.l. and the average slope was 20°. Average annual temperature and precipitation were 13 °C and 938 mm, respectively (average of the period 2004–2018, climatic station S3098, Castagneto Po; ARPA Piemonte 2021). Soil texture was sandy loam and the reaction was neutral. The forest had a two-layers vertical structure, with an oak (Q. robur) top layer (about 20 m height) and a second coppice layer (about 10 m height) dominated by C. sativa and by the alien invasive species R. pseudoacacia. Other mesophilous species in the copse were A. pseudoplatanus, P. avium, and F. ornus. Cumulative canopy cover of both tree layers was about 90%. The coppice wood volume was 180 m3/ha. The shrub layer included species such as C. avellana, Ligustrum vulgare L., Hedera helix L., and Rubus sp., and had an average cover of about 40%. Herbaceous species were sparse and their average cover was < 10%.

a Location of the study area in Piedmont region, Northwestern Italy, and b paddocks set up for pig grazing
The traditional management of the forest was coppice with standards, with the coppice layer (C. sativa) managed for firewood and wooden poles production and the top layer (Q. robur) managed by high forest for industrial timber production. The coppice was cut every 10–15 years, while the oaks were cut when they reached a minimum size of about 50 cm of diameter. The management of the forest has been abandoned since about the early 1990s.
Grazing management and silvicultural interventions
Rotational grazing was implemented in the study area with the ‘Nero di Parma’ breed in 2019. Three paddocks (A, B, and C; Fig. 1b) of 2.8, 3.9 and 5.5 ha in size, respectively, were electrically fenced. Paddocks were adjacent one to another and comparable in terms of vegetation composition and structure. A total of 22 barrows exploited the paddocks in rotation from late March to October 2019. Grazing days of rotation periods depended on each paddock size, on the forage availability inside them, and on the animal weight (Table 1). The farmer moved the pigs from one paddock to another ensuring that grazing pressure was uniform among paddocks. During winter 2019–2020, a coppicing treatment extracted 37% of the copse wood volume of the second tree layer (corresponding to 68 m3/ha) in paddock A, while trees of the top layer were not cut. Tree cutting involved the most abundant species in the paddock, i.e. A. pseudoplatanus, C. sativa, C. avellana, F. ornus, P. avium, and R. pseudoacacia. From late May to August 2020, 29 barrows grazed in paddock A after tree cutting for 90 days (Table 1). Grazing days in paddock A were extended in 2020 compared to 2019 because of the additional forage provided by the resprouts after the silvicultural cutting (Table 1). In both years, the swine were 6 months old and weighed around 60 kg at the start of the grazing period, and reached around 160 kg at the end of the season. To meet pig needs, animals were fed daily with a concentrate with 16.5% of dry matter of crude protein, at a dose increasing during the season from 2 to 3 kg pig−1 day−1, according to the barrow weight. Water was supplied ad libitum using a tank with drinkers, which was regularly moved over the paddock to favor a more homogeneous exploitation of the area. Swine were fitted with nose rings to avoid excessive soil rooting.
Plant species selection
In the first year of the study (2019), we used direct observations to assess the plant species selection by pigs exploiting the understory. Direct observations are commonly used to study livestock feeding preferences in woody-dominated habitats (e.g., Rodríguez-Estévez et al. 2009; Elias and Tischew 2016). Pigs were accustomed to humans, so that observers were allowed to stay close to them without affecting their behavior. Each observer monitored the behavior of a randomly chosen animal during 30″ observation sessions followed by 2′:30″ breaks between each observation. For each observation, the activity of the pig was attributed to one of the following categories: grazing, rooting, searching for food, walking, resting, and others (including all other activities such as drinking, defecation, social interactions etc.). For the purpose of this study, the grazing activity referred to the consumption of any green tissue (leaves, stems and inflorescences) belonging to both herbaceous and woody plant species. Identifying the consumption of other resources such as tree fruits, roots, invertebrates, etc. was not possible. For each grazing observation session, we identified and recorded (i) the plant species consumed by the pig, and (ii) the plants that were not consumed within a 50-cm buffer area around the head of the pig and at a maximum above ground height of 1 m. We assumed that all the plants available in this buffer area were detectable by the pig and that a 1-m height corresponded to the layer that animals could exploit (threshold set according to our field observations). The nomenclature of plant species followed Pignatti et al. (2017).
The monitoring was carried out from May to August 2019, i.e. during the vegetation growing season, for a total of 10 monitoring days: 2 days in paddock A, four in paddock B, and four in paddock C. Total monitoring hours were 134: 41 h in paddock A, 49 in paddock B, and 44 in paddock C. The observations were carried out approximately from 12:00 to 17:00, i.e. avoiding the morning hours when the farmer provided the feed supplements.
Browsing on post-cut tree resprouts
In the second year of the study (2020), we assessed the impact of pig browsing on tree regeneration of A. pseudoplatanus, C. sativa, C. avellana, F. ornus, P. avium, and R. pseudoacacia, for a total of 86 stumps randomly distributed over paddock A. Specifically, we monitored (i) the damages to tree shoots sprouted after the silvicultural cuts and (ii) the shoot growth along the growing season. The monitoring of shoot development was carried out at 2-week intervals throughout the growing season, for a total of seven survey dates. On the first date, corresponding to the day before barrows entered the paddock, we checked the presence and development of suckers emitted by the stumps after the cutting. The six subsequent surveys were used to assess the effect of pig browsing on shoot development. We attributed to each stump an identification code and assessed its post-cut diameter. The variables measured at each survey date were: number of shoots per stump, presence/absence of browsing damage to leaves and to apical buds, and individual shoot height. From these data we computed the percentage of damaged shoots and the average shoot height per each stump and survey date.
Statistical analyses
To assess the plant species selection, we computed for each species the number of ‘consumption’ events (i.e., the number of grazing observations when the plant species was consumed) and the number of ‘no consumption’ events (i.e., the number of grazing observations when the plant species was available but not consumed). Then, a Chi-square test on the frequencies of ‘consumption’ vs ‘no consumption’ events was performed for each species to discriminate three groups of plants: preferred, indifferently consumed, and avoided plants. The null hypothesis was that there were no statistical differences between the number of ‘consumption’ and ‘no consumption’ events. In case of acceptance of the null hypothesis, the species was considered ‘indifferently consumed’ by the pigs. In case of rejection of the null hypothesis (at p < 0.05), the species was considered as ‘preferred’ when the number of ‘consumption’ events was higher than that of ‘no consumption’ events, while it was considered as ‘avoided’ when it was lower. According to Chi-square assumptions, only the plant species being recorded in at least ten grazing observation sessions were tested. All grazing observations from the three paddocks were joined in the analysis.
To assess the impact of pig browsing on the post-cut tree resprouts throughout time, we performed a Generalized Linear Mixed Model (GLMM) with a beta distribution to model the percentage of damaged shoots in function of: (i) time by species interaction, to test the hypothesis that the damage to tree shoots changed linearly over time but differed by species; (ii) stump diameter, to test the hypothesis that damage rate decreased with larger stumps; and (iii) shoot density, to test the hypothesis that browsing rate decreased as the number of shoots increased, i.e. a density-dependent economy of scale. Specifically, time by species interaction, shoot density, and stump diameter were set as fixed factors, while stump was set as random factor to account for the repeated measurements over time. Shoot density and stump diameter were standardized (z-score). A second model was built to assess the impact of pig browsing on shoot height throughout time. In this case we used a Generalized Additive Mixed Model (GAMM) since shoot heights might increase and decrease non-linearly through time, according to the initial resprouting vigor and browsing effect. Specifically, shoot height was assumed to follow a Gaussian distribution after log-transformation, and was modeled in function of time by species interaction as fixed factor and stump as random factor to account for the repeated measurements over time.
The R software (R Core Team 2018) was used for statistical analysis. The GLMM and GAMM were run with the ‘glmmTMB’ and ‘mgcv’ packages, respectively. The ‘visreg’ package was used to graphically visualize the relationship between the response and the explanatory variables.
Results
In total, we recorded 2435 observations of animal behavior, among which, rooting (30% of the total) and grazing (24%) were the prevailing ones, followed by resting (17%), walking (13%), searching for food (10%), and others (6%).
According to the Chi-square tests, three plant species in the understory were preferred, nine species were consumed indifferently, and 18 species were avoided (Table 2). Robinia pseudoacacia showed the highest percent consumption (84%), whereas Ruscus aculeatus L., Euonymus europaeus L., Carex sylvatica Huds., Crataegus monogyna Jacq., Asphodelus albus Mill., Lonicera caprifolium L., and Viburnum lantana L. were never eaten by pigs.
The total number of stump shoots monitored after the silvicultural cut was 987, on average, at each survey date. The percentage of shoots damaged by pig browsing significantly increased through time for C. sativa, C. avellana, P. avium, and R. pseudoacacia, with higher slopes for the first three species than for R. pseudoacacia (Table 3, Fig. 2). At the end of the grazing season, damaged shoots were > 75% for C. sativa, C. avellana and P. avium and around 25% for R. pseudoacacia (Fig. 2). Acer pseudoplatanus and F. ornus showed the lowest damage on shoots (< 25%) and no significant changes throughout time. The shoot density (mean = 12, min = 1, max = 74) and stump diameter (mean = 15 cm, min = 2 cm, max = 40 cm) did not affect the percentage of damaged shoots (Table 3). Shoot height (mean = 36 cm, min = 1 cm, max = 3.38 m) significantly changed along the monitoring period for all the plant species (significant smoothers, Table 3). For A. pseudoplatanus, F. ornus and R. pseudoacacia, shoot height increased with time (Fig. 3), with R. pseudoacacia reaching the highest value at the end of the season (159 cm, on average). The height of P. avium shoots increased at the beginning of the season and was more stable in the second half, while that of C. sativa increased and decreased in the first and second half of the season, respectively. The height of C. avellana shoots, instead, decreased during the season (from 11.0 to 6.6 cm, on average).

Relationship between the percentage of damaged shoots and time (survey dates) for each plant species, according to the Generalized Linear Mixed Model. Species codes: ACPS, Acer pseudoplatanus; CASA, Castanea sativa; COAV, Corylus avellana; FROR, Fraxinus ornus; PRAV, Prunus avium; ROPS, Robinia pseudoacacia

Relationship between shoot height (log-transformed) and time (survey dates) for each plant species, according to the Generalized Additive Mixed Model. Species codes: ACPS, Acer pseudoplatanus; CASA, Castanea sativa; COAV, Corylus avellana; FROR, Fraxinus ornus; PRAV, Prunus avium; ROPS, Robinia pseudoacacia
Discussion
The results of our study showed that swine were selective both in the consumption of plant species in the understory and in browsing post-cut tree resprouts. Indeed, only three plant species (R. pseudoacacia, C. avellana, and Rubus sp.) out of 30 were preferred in the understory while more than half (18) were avoided. Concerning the resprouts, C. avellana, C. sativa and P. avium were more impacted than the other three tree species, resulting in poor or no growth throughout time.
The plant species whose resprouts were highly impacted by pigs (i.e., C. avellana, C. sativa, and P. avium) were also eaten with a high rate in the understory (consumption > 45%). Specifically, C. avellana was the second preferred plant species in the understory and its post-cut resprouts were the most affected, suggesting that this species could represent an important forage source for pigs in temperate forests. Corylus avellana foliage is considered of intermediate quality and livestock browse it in many silvopastoral systems (Hejcmanová et al. 2014; Vandermeulen et al. 2018a). The leaves and resprouts of C. sativa were also considerably eaten by pigs, despite the high tannin content (Pistoia et al. 2009). Tannins generally reduce the palatability of forage, but in some circumstances livestock feed on tannin-rich species to wane gastrointestinal parasites (Villalba et al. 2010). The impact on this plant species by swine was also reported by Pistoia et al. (2009), even though at a lower rate (< 30% of leaves and apexes damaged). The selection for P. avium leaves and resprouts was slightly lower than for C. avellana and C. sativa, but still remarkable. Acer pseudoplatanus and F. ornus, instead, were avoided by pigs in the understory and resprouts were only slightly affected. The avoidance for A. pseudoplatanus can be related to its low forage quality (Ravetto Enri et al. 2020) and/or to toxicity issues. The toxicity of A. pseudoplatanus non-woody parts has been linked to atypical myopathy in other monogastric species, like horses (Aboling et al. 2020). Fraxinus ornus is a common fodder plant for goats and sheep in Mediterranean environments and its forage quality is characterized by low crude protein and fiber content (Papachristou 1997; Papachristou et al. 1999). Swine avoided it probably because of the presence of specific chemical compounds, but further research on this topic is needed.
From the point of view of forest management, these results provide useful information about the possible use of free-ranging pigs in coppice systems, particularly in C. sativa and F. ornus stands. Castanea sativa woodlands occur over large areas in Italy, where C. sativa is the third tree per volume (9% of total volume; De Laurentis et al. 2021). They are traditionally managed with short rotation coppice to provide various timber assortments. The high impact on C. sativa resprouts by pigs suggests avoiding pig grazing after a renovation cut until regrowth is above browse height (i.e., about 1 m height). The exploitation of F. ornus coppice stands, typical of the Mediterranean regions, might be instead a feasible option also during the first post-cut years, due to the marked avoidance of this plant by swine.
Unlike other tree species, the preference for R. pseudoacacia was different in the understory and on post-cut resprouts. Indeed, it was the first preferred species in the former, whereas its resprouts were only slightly impacted in the latter. These differences might be determined by the different ages of leaves and shoots, which were older in the understory plants and younger in the post-cut resprouts. Indeed, it is possible that the young tissues contain a higher fraction of toxic compounds, such as robin, robinine, and robitin (Michalska et al. 2019). Poisoning by R. pseudoacacia was documented in other livestock species, such as horses (Vanschandevijl et al. 2010), whereas goats are able to forage on this protein-rich species, likely due to their greater ability to detoxify plant secondary metabolites (Papachristou and Papanastasis 1994; Giger-Reverdin et al. 2020). Interestingly, also Molnár et al. (2021) reported that swine did not consume R. pseudoacacia. Consequently, the resprouts of this fast-growing alien species can be hardly controlled by pigs, at least in the first season after cutting. However, the relationship among consumption, age of the plant tissues, and toxicity of R. pseudoacacia needs further and thorough investigation.
The impact on tree regeneration is a crucial issue in forest grazing, since the young seedlings and plants can be damaged by browsing (López-Sánchez et al. 2016; Öllerer et al. 2019). At the same time, livestock can also facilitate seedling establishment through seed burial and removal of outcompeting vegetation (Kirby et al. 1994; Öllerer et al. 2019). In this study, all oak species (Quercus cerris L., Q. robur, and Quercus pubescens Willd.) were avoided by pigs in the understory (consumption 4–9%), likely due to their low forage quality (Papachristou and Papanastasis 1994; Pistoia et al. 2009; Hejcmanová et al. 2014). These findings agree with Pistoia et al. (2009) and Vandermeulen et al. 2018b, who observed that Q. robur was slightly consumed by pigs and cattle, respectively. Although the impact on oak plants in the understory was very limited, the exclusion of pigs for 2–3 years after renovation cuts would be advisable to avoid acorn consumption and allow undisturbed seedling growth.
Forest grazing is acknowledged for its role in reducing woody vegetation of the understory by both the consumption and trampling (McEvoy et al. 2006; Öllerer et al. 2019). In our study, Rubus sp. was abundant in the understory and it was among the plant species preferred by swine. This result agrees with the preliminary research of Pistoia et al. (2009), suggesting that pig grazing might be a valuable tool to counteract bramble proliferation in temperate abandoned forests, where this species is a major threat to plant diversity for its ability to suppress woodland species (Van Uytvanck and Hoffmann 2009; Harmer et al. 2016). However, specific research is necessary to assess the impact of pig grazing on bramble cover over time in such environments. Hedera helix L. was another abundant woody species in the forest, and despite the presence of toxic saponins (Wink 2010) it was moderately consumed by pigs (consumption 52%). Hedera helix is an important component of roe deer, red deer, and fallow deer diets, especially in autumn and winter (Metcalfe 2005). Being its leaves evergreen, it may thus represent an important forage source for swine from autumn to early spring, when other forages are not available. Shrub species such as L. vulgare and E. europaeus, instead, were avoided by pigs in the understory (consumption ≤ 1%), probably because of the presence of toxic compounds (secoiridoid glycosides in L. vulgare and cardenolides in E. europaeus; Wink 2010). Also C. monogyna was avoided by pigs contrary to the findings of Molnár et al. (2021), who reported, however, that this species was eaten only when leaves were young and spikes were soft.
Herbaceous plant species can benefit from the reduction of shrubs and the increase in light conditions induced by livestock, while on the other hand they can be sensitive to selective consumption, trampling or nutrient addition through urine and faeces (McEvoy et al. 2006; Öllerer et al. 2019). For instance, the palatable Hyacinthoides non-scripta (L.) Chouard ex Rothm. was adversely affected by grazing in Irish temperate forests (McEvoy et al. 2006) and Anemone nemorosa L., although being favored by the reduction of bramble cover, was sensitive to cattle grazing and trampling (Van Uytvanck and Hoffmann 2009). In this study, all the recorded herbaceous species typical of the forest flora, i.e., A. albus, C. sylvatica, Physospermum cornubiense (L.) DC., and Viola riviniana Rchb., were never or only occasionally eaten by pigs, suggesting that they would likely not be threatened by direct consumption. However, long-term studies appear necessary to assess the response of the forest typical flora to the complex integration of grazing, trampling and nutrient addition and to the modification of the shrub-herbs competition induced by pig grazing.
Conclusions
Our research provided novel results concerning the implementation of silvopastoral management practices in degraded deciduous forests, particularly adopting pig grazing. The outcomes of this study revealed a selective foraging behavior of pigs, highlighting contrasting preferences for understory plant species. Indeed, some species were preferred (C. avellana, R. pseudoacacia, and Rubus sp.), others were indifferently consumed (e.g. C. sativa and H. helix), while others were avoided (e.g. Acer spp. and Quercus spp.), likely due to differences in the forage quality and toxicity of plant species. The selection for Rubus sp. suggested that pig grazing might be a valuable tool to control this species in temperate abandoned forests. Conversely, even though R. pseudoacacia leaves from mature plants were highly preferred, its resprouts were not effectively controlled in the first season after the cut.
The outcomes provided by this pilot project could be applied in similar ecological and management conditions and plant species composition all over temperate Europe. Future studies should investigate the effects of pig grazing on bramble cover and on the plant diversity and composition of temperate deciduous forests over time.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Aboling S, Scharmann F, Bunzel D (2020) Equine atypical myopathy: consumption of sycamore maple seedlings (Acer pseudoplatanus) by pastured horses is driven by seedling maturity and might be associated with phenolic compounds. Vet Rec 187(12):e116–e116. https://doi.org/10.1136/vr.105736
ARPA Piemonte (2021) https://www.arpa.piemonte.it. Accessed 26 Oct 2021
Brownlow MJC, Dorward PT, Carruthers SP (2005) Integrating natural woodland with pig production in the United Kingdom: an investigation of potential performance and interactions. Agroforest Syst 64(3):251–263. https://doi.org/10.1007/s10457-004-0250-6
Elias D, Tischew S (2016) Goat pasturing—a biological solution to counteract shrub encroachment on abandoned dry grasslands in Central Europe? Agric Ecosyst Environ 234:98–106. https://doi.org/10.1016/j.agee.2016.02.023
Giger-Reverdin S, Domange C, Broudiscou LP et al (2020) Rumen function in goats, an example of adaptive capacity. J Dairy Res 87(1):45–51. https://doi.org/10.1017/S0022029920000060
Harmer R, Kerr G, Stokes V, Connolly T (2016) The influence of thinning intensity and bramble control on ground flora development in a mixed broadleaved woodland. Forestry 90(2):247–257. https://doi.org/10.1093/forestry/cpw048
Hejcmanová P, Stejskalová M, Hejcman M (2014) Forage quality of leaf-fodder from the main broad-leaved woody species and its possible consequences for the Holocene development of forest vegetation in Central Europe. Veget Hist Archaeobot 23(5):607–613. https://doi.org/10.1007/s00334-013-0414-2
Jørgensen D (2013) Pigs and pollards: medieval insights for UK wood pasture restoration. Sustainability 5(2):387–399. https://doi.org/10.3390/su5020387
Kirby KJ, Mitchell FJ, Hester AJ (1994) A role for large herbivores (deer and domestic stock) in nature conservation management in British semi-natural woods. Arboricultural J 18(4):381–399. https://doi.org/10.1080/03071375.1994.9747043
De Laurentis D, Papitto G, Gasparini P et al (2021) Le Foreste Italiane. Sintesi dei risultati del terzo Inventario Forestale Nazionale INFC2015. Arma dei Carabinieri & CREA - Centro di Ricerca Foreste e Legno
López-Sánchez A, Perea R, Dirzo R, Roig S (2016) Livestock vs. wild ungulate management in the conservation of Mediterranean dehesas: implications for oak regeneration. For Ecol Manag 362:99–106. https://doi.org/10.1016/j.foreco.2015.12.002
Mayle B (1999) Domestic stock grazing to enhance woodland biodiversity. Forestry Commission, Bristol
McEvoy PM, Flexen M, McAdam JH (2006) The effects of livestock grazing on ground flora in broadleaf woodlands in Northern Ireland. For Ecol Manag 225(1–3):39–50. https://doi.org/10.1016/j.foreco.2005.12.026
Metcalfe DJ (2005) Hedera helix L. J Ecol 93(3):632–648. https://doi.org/10.1111/j.1365-2745.2005.01021.x
Miao ZH, Glatz PC, Ru YJ (2004) Review of production, husbandry and sustainability of free-range pig production systems. Asian Australas J Anim Sci 17(11):1615–1634. https://doi.org/10.5713/ajas.2004.1615
Michalska K, Tomczyk A, Łotocka B et al (2019) Oviposition by the vagrant eriophyoid mite Aculops allotrichus on leaves of black locust tree Robinia pseudoacacia. Exp Appl Acarol 79(1):1–19. https://doi.org/10.1007/s10493-019-00412-1
Molnár Z, Szabados K, Kiš A et al (2021) Preserving for the future the—once widespread but now vanishing—knowledge on traditional pig grazing in forests and marshes (Sava-Bosut floodplain, Serbia). J Ethnobiol Ethnomed 17(1):56. https://doi.org/10.1186/s13002-021-00482-9
Moreno G, Aviron S, Berg S et al (2018) Agroforestry systems of high nature and cultural value in Europe: provision of commercial goods and other ecosystem services. Agroforest Syst 92(4):877–891. https://doi.org/10.1007/s10457-017-0126-1
Mosquera-Losada MR, Santiago Freijanes JJ, Pisanelli A et al (2016) Extent and success of current policy measures to promote agroforestry across Europe. Deliverable 823 for EU FP7 Research Project: AGFORWARD 613520 (8 December 2016) 95pp
Öllerer K, Varga A, Kirby K et al (2019) Beyond the obvious impact of domestic livestock grazing on temperate forest vegetation—a global review. Biol Conserv 237:209–219. https://doi.org/10.1016/j.biocon.2019.07.007
Papachristou TG (1997) Intake, digestibility and nutrient utilization of oriental hornbeam and manna ash browse by goats and sheep. Small Rumin Res 23(2–3):91–98. https://doi.org/10.1016/S0921-4488(96)00909-1
Papachristou TG, Papanastasis VP (1994) Forage value of Mediterranean deciduous woody fodder species and its implication to management of silvo-pastoral systems for goats. Agroforest Syst 27(3):269–282. https://doi.org/10.1007/BF00705061
Papachristou TG, Platis PD, Papanastasis VP, Tsiouvaras CN (1999) Use of deciduous woody species as a diet supplement for goats grazing Mediterranean shrublands during the dry season. Anim Feed Sci Technol 80(3–4):267–279. https://doi.org/10.1016/S0377-8401(99)00056-5
Pauler CM, Isselstein J, Suter M et al (2020) Choosy grazers: Influence of plant traits on forage selection by three cattle breeds. Funct Ecol 34(5):980–992. https://doi.org/10.1111/1365-2435.13542
Pignatti S, La Rosa M, Guarino R (2017) Flora d’Italia, 2nd edn. Edagricole di New Business Media, Bologna
Pistoia A, Poli P, Casarosa L et al (2009) Preliminary research on environmental impact of woodland grazing by pigs. Ital J Anim Sci 8:325–327
Pittarello M, Gorlier A, Lombardi G, Lonati M (2017) Plant species selection by sheep in semi-natural dry grasslands extensively grazed in the south-western Italian Alps. Rangel J 39(2):123–131. https://doi.org/10.1071/RJ16068
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ravetto Enri S, Probo M, Renna M et al (2020) Temporal variations in leaf traits, chemical composition and in vitro true digestibility of four temperate fodder tree species. Anim Prod Sci 60(5):643. https://doi.org/10.1071/AN18771
Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (2009) Agroforestry in Europe: current status and future prospects. Springer, Dordrecht
Rodríguez-Estévez V, García A, Peña F, Gómez AG (2009) Foraging of Iberian fattening pigs grazing natural pasture in the dehesa. Livest Sci 120(1–2):135–143. https://doi.org/10.1016/j.livsci.2008.05.006
Silva JS, Nunes JLT (2013) Inventory and characterization of traditional Mediterranean pig production systems. Advantages and constraints towards its development. Acta Agric Slov 4:61–67
Van Uytvanck J, Hoffmann M (2009) Impact of grazing management with large herbivores on forest ground flora and bramble understorey. Acta Oecol 35(4):523–532. https://doi.org/10.1016/j.actao.2009.04.001
Vandermeulen S, Ramírez-Restrepo CA, Beckers Y et al (2018a) Agroforestry for ruminants: a review of trees and shrubs as fodder in silvopastoral temperate and tropical production systems. Anim Prod Sci 58(5):767–777. https://doi.org/10.1071/AN16434
Vandermeulen S, Ramírez-Restrepo CA, Marche C et al (2018b) Behaviour and browse species selectivity of heifers grazing in a temperate silvopastoral system. Agrofor Syst 92(3):705–716. https://doi.org/10.1007/s10457-016-0041-x
Vanschandevijl K, Van Loon G, Lefère L, Deprez P (2010) Black locust (Robinia pseudoacacia) intoxication as a suspected cause of transient hyperammonaemia and enteral encephalopathy in a pony. Equine Vet Educ 22(7):336–339. https://doi.org/10.1111/j.2042-3292.2010.00090.x
Varela E, Górriz-Mifsud E, Ruiz-Mirazo J, López-i-Gelats F (2018) Payment for targeted grazing: integrating local shepherds into wildfire prevention. Forests 9(8):464. https://doi.org/10.3390/f9080464
Villalba JJ, Provenza FD, Hall JO, Lisonbee LD (2010) Selection of tannins by sheep in response to gastrointestinal nematode infection1. J Anim Sci 88(6):2189–2198. https://doi.org/10.2527/jas.2009-2272
Vítková M, Müllerová J, Sádlo J et al (2017) Black locust (Robinia pseudoacacia) beloved and despised: a story of an invasive tree in Central Europe. For Ecol Manag 384:287–302. https://doi.org/10.1016/j.foreco.2016.10.057
Wink M (2010) Mode of action and toxicology of plant toxins and poisonous plants. Julius-Kühn-Archiv 24(421):93
Acknowledgements
The authors thank Davide Lovera (‘Parva Domus’ farm) for the provision of the animals and its constant collaboration in the study and Alessandra Garro, Andrea Mainetti, and Elena Quaglia for helping in the fieldwork.
Funding
Open access funding provided by Università degli Studi di Torino within the CRUI-CARE Agreement. The study was funded by the Rural Development Program of Piedmont Region 2014–2020, project ‘Food For Forest – Selvipastorizia rigenerativa: il cibo nutre la foresta’ (Food For Forest – Restorative silvopastoralism: the food that feeds the forest).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nota, G., Berretti, R., Ascoli, D. et al. Plant species selection and impact on tree resprouts by semi-free ranging pigs in a temperate deciduous forest. Agroforest Syst 97, 121–132 (2023). https://doi.org/10.1007/s10457-022-00792-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-022-00792-1