
Abstract
Background
The role of mesenchymal stem/stromal cells (MSCs) in tumorigenesis remains controversial. This study aimed to determine whether heterotypic interactions between MSCs and colon cancer cells can supply contextual signals towards tumor progression.
Methods
Xenografts consisting of co-implanted human colorectal cancer cells with rat MSCs in immunodeficient mice were evaluated by tumor progression, angiogenic profiles, and MSC fate. Furthermore, we investigated how MSCs function as a cancer cell niche by co-culture experiments in vitro.
Results
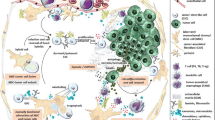
Tumor growth progressed in two ways, either independent of or dependent on MSCs. Such cell line-specific dependency could not be explained by host immune competency. COLO 320 xenograft angiogenesis was MSC-dependent, but less dependent on vascular endothelial growth factor (VEGF), whereas HT-29 angiogenesis was not MSC-dependent, but was VEGF-dependent. MSCs and COLO 320 cells established a functional positive feedback loop that triggered formation of a cancer cell niche, leading to AKT activation. Subsequently, MSCs differentiated into pericytes that enhanced angiogenesis as a perivascular niche. In contrast, the MSC niche conferred an anti-proliferative property to HT-29 cells, through mesenchymal–epithelial transition resulting in p38 activation.
Conclusions
In conclusion, MSCs demonstrate pleiotropic capabilities as a cancer cell or perivascular niche to modulate colorectal cancer cell fate in a cell line-dependent manner in a xenogeneic context.






Similar content being viewed by others


Abbreviations
- MSCs:
-
Mesenchymal stem/stromal cells
- TME:
-
Tumor microenvironment
- CAFs:
-
Cancer-associated fibroblastic cells
- CCL5:
-
C–C motif chemokine ligand 5
- EMT:
-
Epithelial–mesenchymal transition
- IL-6:
-
Interleukin-6
- VEGF:
-
Vascular endothelial growth factor
- eGFP:
-
Enhanced green fluorescence protein
- αMEM:
-
α-Modified Eagle’s medium
- FBS:
-
Fetal bovine serum
- qRT-PCR:
-
Quantitative real-time reverse transcription PCR
- MVD:
-
Tumor microvessel density
- Thy-1:
-
Thymus cell antigen-1
- NG2:
-
Neural/glial antigen 2
- αSMA:
-
α-Smooth muscle actin
- OE:
-
Overexpression
- CXCL12:
-
Chemokine C-X-C motif ligand 12
- KD:
-
Knock down
- MSC-CM:
-
MSC-conditioned medium
- CXCR4:
-
C-X-C chemokine receptor type 4
- MAPKs:
-
Mitogen-activated protein kinases
- FACS:
-
Fluorescence-activated cell sorting
- ANOVA:
-
Analysis of variance
- PECAM-1:
-
Platelet endothelial cell adhesion molecule-1
- Vegfr1 (Flt1):
-
Vascular endothelial growth factor receptor 1
- PDGF-BB:
-
Platelet-derived growth factor BB
- Pdgfr-β:
-
Platelet-derived growth factor receptor-β
- MET:
-
Mesenchymal–epithelial transition
- Vcam1:
-
Vascular cell adhesion molecule-1
- CCR5:
-
Chemokine (C–C motif) receptor 5
- VLA-4:
-
Very late antigen-4
References
Bissell MJ, Radisky D. Putting tumours in context. Nat Rev Cancer. 2001;1:46–54.
Hall B, Andreeff M, Marini F. The participation of mesenchymal stem cells in tumor stroma formation and their application as targeted-gene delivery vehicles. Handb Exp Pharmacol. 2007;263–83.
Park CC, Bissell MJ, Barcellos-Hoff MH. The influence of the microenvironment on the malignant phenotype. Mol Med Today. 2000;6:324–9.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74.
Khakoo AY, Pati S, Anderson SA, et al. Human mesenchymal stem cells exert potent antitumorigenic effects in a model of Kaposi’s sarcoma. J Exp Med. 2006;203:1235–47.
Karnoub AE, Dash AB, Vo AP, et al. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature. 2007;449:557–63.
Tanaka H, Arimura Y, Yabana T, et al. Myogenic lineage differentiated mesenchymal stem cells enhance recovery from dextran sulfate sodium-induced colitis in the rat. J Gastroenterol. 2011;46:143–52.
Yabana T, Arimura Y, Tanaka H, et al. Enhancing epithelial engraftment of rat mesenchymal stem cells restores epithelial barrier integrity. J Pathol. 2009;218:350–9.
Hogan NM, Dwyer RM, Joyce MR, Kerin MJ. Mesenchymal stem cells in the colorectal tumor microenvironment: recent progress and implications. Int J Cancer. 2012;131:1–7.
Tsai K-S, Yang S-H, Lei Y-P, et al. Mesenchymal stem cells promote formation of colorectal tumors in mice. Gastroenterology. 2011;141:1046–56.
Shinagawa K, Kitadai Y, Tanaka M, et al. Mesenchymal stem cells enhance growth and metastasis of colon cancer. Int J Cancer. 2010;127:2323–33.
De Boeck A, Pauwels P, Hensen K, et al. Bone marrow-derived mesenchymal stem cells promote colorectal cancer progression through paracrine neuregulin 1/HER3 signalling. Gut. 2013;62:550–60.
Lin J-T, Wang J-Y, Chen M-K, et al. Colon cancer mesenchymal stem cells modulate the tumorigenicity of colon cancer through interleukin 6. Exp Cell Res. 2013;319:2216–29.
Liu Y, Han Z, Zhang S, et al. Effects of inflammatory factors on mesenchymal stem cells and their role in the promotion of tumor angiogenesis in colon cancer. J Biol Chem. 2011;286:25007–15.
Li H-J, Reinhardt F, Herschman HR, Weinberg RA. Cancer-stimulated mesenchymal stem cells create a carcinoma stem cell niche via prostaglandin E2 signaling. Cancer Discov. 2012;2:840–55.
Mele V, Muraro MG, Calabrese D, et al. Mesenchymal stromal cells induce epithelial-to-mesenchymal transition in human colorectal cancer cells through the expression of surface-bound TGF-β. Int J Cancer. 2014;134:2583–94.
Hakamata Y, Tahara K, Uchida H, et al. Green fluorescent protein-transgenic rat: a tool for organ transplantation research. Biochem Biophys Res Commun. 2001;286:779–85.
Ito M, Hiramatsu H, Kobayashi K, et al. NOD/SCID/gamma(c)(null) mouse: an excellent recipient mouse model for engraftment of human cells. Blood. 2002;100:3175–82.
Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–7.
Javazon EH, Colter DC, Schwarz EJ, Prockop DJ. Rat marrow stromal cells are more sensitive to plating density and expand more rapidly from single-cell-derived colonies than human marrow stromal cells. Stem Cells. 2001;19:219–25.
Kuan SF, Byrd JC, Basbaum CB, Kim YS. Characterization of quantitative mucin variants from a human colon cancer cell line. Cancer Res. 1987;47:5715–24.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods. 2001;25:402–8.
Weidner N, Semple JP, Welch WR, Folkman J. Tumor angiogenesis and metastasis—correlation in invasive breast carcinoma. N Engl J Med. 1991;324:1–8.
Iyonaga K, Takeya M, Yamamoto T, et al. A novel monoclonal antibody, RM-4, specifically recognizes rat macrophages and dendritic cells in formalin-fixed, paraffin-embedded tissues. Histochem J. 1997;29:105–16.
Tamaki T, Akatsuka A, Okada Y, et al. Cardiomyocyte formation by skeletal muscle-derived multi-myogenic stem cells after transplantation into infarcted myocardium. PLoS ONE. 2008;3:e1789.
Grunewald M, Avraham I, Dor Y, et al. VEGF-induced adult neovascularization: recruitment, retention, and role of accessory cells. Cell. 2006;124:175–89.
Wynn RF, Hart CA, Corradi-Perini C, et al. A small proportion of mesenchymal stem cells strongly expresses functionally active CXCR4 receptor capable of promoting migration to bone marrow. Blood. 2004;104:2643–5.
Karp JM, Leng Teo GS. Mesenchymal stem cell homing: the devil is in the details. Cell Stem Cell. 2009;4:206–16.
Rattigan Y, Hsu J-M, Mishra PJ, et al. Interleukin 6 mediated recruitment of mesenchymal stem cells to the hypoxic tumor milieu. Exp Cell Res. 2010;316:3417–24.
Nasuno M, Arimura Y, Nagaishi K, et al. Mesenchymal stem cells cancel azoxymethane-induced tumor initiation. Stem Cells. 2014;32:913–25.
Vertelov G, Kharazi L, Muralidhar MG, et al. High targeted migration of human mesenchymal stem cells grown in hypoxia is associated with enhanced activation of RhoA. Stem Cell Res Ther. 2013;4:5.
Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delaunay T, Naeem R, et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell. 2005;121:335–48.
Acknowledgments
We are very grateful to Ms. K. Fujii of First Department of Internal Medicine, for technical assistance, and Dr. Y. Sasaki of Medical Genome Sciences, Research Institute for Frontier Medicine, Sapporo Medical University, for critical comments. We are also thankful to Dr. M. Tsuji of Chromosome Science Labo Inc., for providing FISH probes and technical advice. This work was supported in part by Health and Labor Sciences Research Grants for research on intractable diseases from the Ministry of Health, Labour, and Welfare of Japan (K.I. and Y.A.).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
S. Nakagaki and Y. Arimura contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
535_2015_1049_MOESM2_ESM.tif
Supplementary Fig. 1 Xenografts analysis and co-culture experiments. To examine whether promotion of tumor growth by MSCs was dependent on the ratio of tumor cells that were co-implanted, xenografts were formed at various ratios of MSCs to COLO 320 cells (2:1 to 0:1) in SCID mice [3]. The 1:1 ratio was used for subsequent analyses because of maximal ability to promote tumor growth. M represents MSCs and C indicates COLO-320 cells. Average data from multiple experiments are shown (a total of six tumors from three mice for each cell line) (a). COLO 320 xenografts without co-implanted MSCs failed to engraft even over a longer period (day 32) observation period in SCID mice (b). Angiogenic factors in cell lines and culture supernatants were quantified by qRT-PCR (c) and quantitative real-time immuno-PCR (iPCR) (d), respectively. iPCR [4] was performed as sandwich assays using an angioplex detection kit in microtiter plates pre-coated with anti-fibroblast growth factor-2 (FGF-2), anti-hepatocyte growth factor (HGF), or anti-VEGF antibodies (Synthera Technologies, Tokyo, Japan) according to the manufacturer’s instructions. Briefly, the analyte (i.e. culture supernatant) as well as the standard and negative controls were diluted, and for subsequent detection of immunocomplexes (capture antibody–analyte antigen-detection antibody), MUSTag (multiple simultaneous tag) mix was added, followed by incubation for 1 h at room temperature (RT). After EcoRI digestion and the retrieved reaction mixture was added, real-time PCR was conducted in a PRISM 7500 Sequence Detection System (Applied Biosystems) for 50 cycles of a two-step PCR amplification protocol (95 °C for 15 s and 60 °C for 1 min). Oligo-tags of FGF-2, HGF, and VEGF antibodies were labeled by FAM, HEX, and Cy5 fluorochromes, respectively. For flow cytometry, a single cell suspension of colon cancer cells co-cultured with MSCs was fixed in 90 % cold ethanol, treated with RNase A, and stained with propidium iodide. (d) Histograms of cell cycle analysis of co-cultured colon cancer cells using FACS after eliminating MSCs by gating on GFP-negative cells. (e) The percentages of each population gated in panel d are shown. The G0/G1 peak was decreased and the G2/M was increased in co-cultured COLO 320 cells. In contrast, the G0/G1 peak was increased whereas the G2/M was decreased in co-cultured HT-29 cells compared with that in mono-cultured cells. (f) The 11 cytokines, Egf, Tgfα, Tgfβ, Vegf, Igf1, Ptgs2, Hgf, Fgf2, Tnfα, Il1β, and Il10, was analyzed by qRT-PCR in MSCs separated by FACS. Relative expression of Il1β compared with that of the steady state of a single culture of MSCs was 5.7-fold and 744-fold in MSCs co-cultured with COLO320 and HT-29 cells, respectively. Whether abundant IL-1β production induced by co-cultured MSCs (approximately 130-fold more abundant in co-cultured MSCs with HT-29 than that in COLO 320 cells) could activate p38 was not likely according to a report by Liu et al. [5]. Recently, Waterman et al. [6] classified MSCs into two distinct phenotypes: MSC1 has pro-inflammatory and anti-tumor effects, while MSC2 has an immunosuppressive role and promotes tumor growth and metastases. Although these classifications appeared to be similar to the phenotypes of MSCs co-cultured with HT-29 cells and COLO 320 cells, respectively, there were some inconsistencies concerning IL-1β and IL-10 expression in our observations (TIFF 1180 kb)
535_2015_1049_MOESM3_ESM.tif
Supplementary Fig. 2. Candidate panel of potential cancer niche signals and CXCL12 CpG island methylation. The candidate panel for MSC niche signaling molecules included wingless-type MMTV integration site family, member 3A (Wnt3a) and 5a (Wnt5a), secreted frizzled-related protein 1 (Sfrp1) and 4 (Sfrp4), dikkopf-1 (Dkk1), angiopoetin-1 (Angpt1), thyroid peroxidase (Tpo), N-Cadherin (Cdh2), integrin β1 (Itgb1) and α4 (Itga4), vascular cell adhesion molecule-1 (Vcam1), sec1 family domain containing 1 (Scfd1) and 2 (Scfd2), Nestin, glial fibrillary acidic protein (Gfap), chemokine (C-X-C motif) ligand 12 (Cxcl12), β-catenin (Ctnnb1), E-cadherin (Cdh1), Jagged 1 (Jag1) and 2 (Jag2), Delta-like 1 (Dll1), 2 (Dll2), and 4 (Dll4), and chemokine (C–C motif) ligand 5 (CCL5). This list was obtained from literature describing hematopoietic stem cell niche molecules [7], Notch signaling [8], and de novo secretion of chemokines from MSCs [9]. Relative expression of the indicated transcripts in co-cultured MSCs with COLO 320 was analyzed by qRT-PCR compared with that of single-cultured MSC. The Y-axis was a hemi-logarithmic scale. Six transcripts of Vcam1, Cxcl12, Cdh1, Jag1 and 2, and Ccl5 were upregulated in co-cultured MSCs compared with single-cultured MSCs (a). (b) Western blot analysis was conducted in COLO 320 cells co-cultured with MSCs, which was corresponding to Fig. 5e. Co-cultured COLO 320 cells without any treatment were used as a control. Data are representative of three independent experiments. (c) CXCL12 transcripts were analyzed by qPCR. (d) CpG island methylation at the CXCL12 promoter was quantified by bisulfite-pyrosequencing. The regions upstream from the transcription start site (CXCL12: -87 to -181 bp) were examined. DMTsKO indicates knockout of DNMT-1, -3a, and 3b in HCT116 cells. A normal colon, which represented histopathological normal specimens resected for medical reasons, was used for the negative control. Genomic DNA (1 μg) was modified with sodium bisulfite using an EpiTect Bisulfite kit (Qiagen), and bisulfite pyrosequencing analysis was performed as described previously [10]. Although we hypothesized that epigenetic silencing of CXCL12 as reported by Wendt et al. [11] was exceptionally cancelled by DNA demethylation in COLO 320 cells, further analysis was necessary to test this hypothesis. (e) MicroRNAs, miR-126 (miRBase accession no. MIMAT0000445) and miR-126* (miRBase accession no. MIMAT0000444), the partner to miR-126 that is derived from the same transcript, EGF-like-domain, multiple 7 (EGFL7) were investigated. The total RNA of cells was isolated with the TaqMan MicroRNA Cells-to-CT Kit (Applied Biosystems) according to the manufacturer’s instructions. We used TaqMan MicroRNA Assays to quantify mature miRNA expression. Zhang et al. [12] recently reported that miR-126/miR-126* directly inhibits CXCL12 expression and independently suppresses the sequential recruitment of MSCs into the tumor stroma in a mouse xenograft model. However, since miR-126/miR-126* was expressed approximately 15–20-fold more in COLO 320 cells than in HT-29 cells in our analysis (e), thus miR-126/miR-126* was not likely to be a major regulatory mechanism of CXCL12 in this setting. As for CXCR4 upregulation, it is entirely an open question as to what cell types express it, and what mechanism of production occurs in xenograft tumors with or without MSCs (TIFF 635 kb)
Rights and permissions
About this article

Cite this article
Nakagaki, S., Arimura, Y., Nagaishi, K. et al. Contextual niche signals towards colorectal tumor progression by mesenchymal stem cell in the mouse xenograft model. J Gastroenterol 50, 962–974 (2015). https://doi.org/10.1007/s00535-015-1049-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00535-015-1049-0