
Abstract
An immunotherapeutic strategy is discussed supporting anti-tumor activity toward malignancies overexpressing ganglioside D3. GD3 can be targeted by NKT cells when derived moieties are presented in the context of CD1d. NKT cells can support anti-tumor responses by secreting inflammatory cytokines and through cytotoxicity toward CD1d+GD3+ tumors. To overexpress GD3, we generated expression vector DNA and an adenoviral vector encoding the enzyme responsible for generating GD3 from its ubiquitous precursor GM3. We show that DNA encoding α-N-acetyl-neuraminide α-2,8-sialyltransferase 1 (SIAT8) introduced by gene gun vaccination in vivo leads to overexpression of GD3 and delays tumor growth. Delayed tumor growth is dependent on CD1d expression by host immune cells, as shown in experiments engaging CD1d knockout mice. A trend toward greater NKT cell populations among tumor-infiltrating lymphocytes is associated with SIAT8 vaccination. A single adenoviral vaccination introduces anti-tumor activity similarly to repeated vaccination with naked DNA. Here, greater NKT tumor infiltrates were accompanied by marked overexpression of IL-17 in the tumor, later switching to IL-4. Our results suggest that a single intramuscular adenoviral vaccination introduces overexpression of GD3 by antigen-presenting cells at the injection site, recruiting NKT cells that provide an inflammatory anti-tumor environment. We propose adenoviral SIAT8 (AdV-SIAT8) can slow the growth of GD3 expressing tumors in patients.
Similar content being viewed by others

Introduction
Gangliosides are glycosphingolipids found within lipid rafts of the plasma membrane [1] and contribute to cell adhesion and motility [2]. This is particularly important during neuronal development [3]. Of particular interest is disialoganglioside D3 (GD3), generated by the addition of a sialic acid residue to endogenous ganglioside M3 (GM3) by the enzyme α-N-acetyl-neuraminide α-2,8-sialyltransferase 1 or GD3 synthase, the expression of which occurs in the Golgi [4] and separates GD3 expressing from non-expressing cells.
During development, the ganglioside is found in the cerebral cortex, hippocampus, thalamus and cerebellum of the brain, whereas expression in adults is found in the neural retina [5]. GD3 is also expressed by neuroendocrine tumor cells, particularly melanoma [6]. We have similarly reported overexpression of GD3 in lung tumors that develop in lymphangioleiomyomatosis [7]. When expressed in tumors, the ganglioside supports metastasis, promotes angiogenesis and interacts with growth factor receptors [8]. Circulating gangliosides shed by tumors can be cytotoxic by introducing mitochondrial damage in cells [9] when the ganglioside is not deactivated by O-acetylation [10]. Preferential expression of GD3 by tumors or immune privileged tissue [11] renders the molecule suited as a target of immunotherapeutic anti-tumor vaccines.
Previous attempts to target GD3 have involved antibody or anti-idiotype administration [12, 13]. Phase III clinical trials offered a unique attempt at immunotherapy for small cell lung cancer [14]. The success of antibody-based therapy can, however, be limited, as humoral responses depend on additional immune components for complement-dependent cytotoxicity or antibody-dependent cellular cytotoxicity [15]. Moreover, the targeted tumor type, small cell lung cancer (SCLC), is a particularly aggressive tumor type that evades treatment [16]. Meanwhile, these trial therapies revealed limited autoimmune side effects, suggesting that GD3 membrane expression is quite tumor-specific and that targeting GD3 is relatively safe, though safety of a more aggressive approach using anti-GD3 chimeric antigen-receptor-transduced T cells has yet to be shown [17]. Treatment efficacy may be more clearly revealed when targeting less aggressive GD3-expressing tumors and when activating an effector mechanism beyond antibodies and their derivatives.
We thus set out to design a vaccine to recruit natural killer T (NKT) cells to the tumor. Classical NKT cells express a restricted T cell receptor repertoire with an invariant Vα14Jα18 chain in mice and Vα24Jα18 in humans. Upon recognition of cognate antigen, NKTs release type 1 cytokines including interferon-γ (IFN-γ) or type 2 cytokines including IL-4 to induce adaptive immune responses. Production of IL-10 or IL-17A and others is also possible [18].
NKT cells can thus support ongoing immune responses within the tumor environment through cytokine production to assist cytotoxic T cell responses against the tumor. NKT cells can also be directly cytotoxic toward tumor cells expressing a natural NKT ligand and appropriate antigen-presenting moieties [19]. GD3 can be presented to both CD4+ and CD4− NKT cells in the context of cluster of differentiation 1d (CD1d) [20]. Presentation of lipid antigens in the context of CD1d is generally limited to professional antigen-presenting cells [21]. However, expression has been observed on the surface of tumor cells including renal cell carcinomas and melanomas [22, 23]. Taken together, NKT cells can restrict tumor growth [24].
Invariant NKT cells can develop different cytokine profiles depending in part on the interaction affinity with the antigen-presenting cell [19]. Meanwhile, the majority of circulating NKT cells is of the effector memory type [25]. Key to GD3-enhanced stimulation of anti-tumor responses is then whether GD3 expression by antigen-presenting cells can stimulate NKT cell recruitment and cytokine secretion where needed.
The GD3 synthase (SIAT8) or alpha-N-acetyl-neuraminide alpha-2,8-sialyltransferase 1 was overexpressed using an expression vector, and it was assessed whether expression of the enzyme is a sufficient and non-cytotoxic means to elicit GD3 expression. Wildtype and CD1d knockout mice were then gene gun-vaccinated with a combination of SIAT8 and adjuvant inducible heat shock protein 70 (HSP70i) encoding DNA before a subcutaneous B16 mouse melanoma tumor challenge. Tumor growth and depigmentation were followed, and NKT cell influx to tumors was monitored. The SIAT8 enzyme was likewise overexpressed by adenoviral transduction. Tumor regression after a single vaccination of adenovirus has been reported in mice [26]. This application carries particular significance for lung tumors, as adenovirus is responsible for natural pulmonary infections [27]. Administering adenovirus type 5-based vectors has proven safe in mice and humans [28]. Here, resulting cytokine profiles among NKT cells were measured by quantitative reverse transcription–polymerase chain reaction (qRT-PCR), or mice were again subjected to a B16 tumor challenge. A novel vaccination principle is tested to help us understand the role of SIAT8 in defining GD3 expression levels and tumor growth, probe NKT cell responses to GD3, and assess treatment efficacy for tumors overexpressing GD3. These experiments provide important insights for a vaccination strategy that can elicit NKT cell responses against GD3-expressing tumors, potentially keeping tumor growth in check.
Materials and Methods
Human and mouse SIAT8 cloning and expression verification
RNA was harvested from human melanoma cell lines M14 (American Type Culture Collection, Manassas, VA) and reverse-transcribed, and cDNA clone MGC-36044 (ATCC) served as a template for amplification of mouse GD3 synthase using primers (Invitrogen, Darmstadt, Germany) for human F: 5′ATGAGCCCCTGCGGGCGGGCCCGGCG 3′ R: 5′CTAGGAAGTGGGCTGGAGTGAGGTATC3′, and mouse F: 5′ATGAGCCCCTGCGGGCGGGCCCTAC3′, R: 5′CTAGGAAGTGGGCTGTGGTGACGGC3′ SIAT8 enzymes. Amplified SIAT8 DNA including a stop codon was cloned into the pEF6/V5-His TOPO TA expression vector and introduced into chemically competent One Shot TOP10 E. coli (Invitrogen). Human embryonic kidney 293 (HEK293) cells were transfected with the human or mouse SIAT8 plasmid using Lipofectamine LTX Reagent (Invitrogen). Cells were indirectly stained using K-18 goat polyclonal antibodies to GD3 synthase (Santa Cruz), and mouse IgG3 antibody clone R24 to GD3 (Invitrogen). Fluorescently labeled, species-specific secondary antibodies (Southern Biotechnologies) were added before fluorocytometric analysis.
Complement-dependent cytotoxicity assay
Complement-dependent cytotoxicity (CDC) assays were performed by plating SIAT8-transfected HEK293 cells in triplicate in 96-well plates. Adherent cells were subjected to 5 μg/ml azide-free R24 antibody (Invitrogen) and normal human serum complement solution (Quidel Corporation, San Diego, CA) in culture medium. Maximum cytotoxicity values were established from wells incubated with 0.5% Triton-X-100 (Sigma-Aldrich, St. Louis, MO). Background signal was determined from untreated wells. After 16 h, cells were subjected to an MTT assay (Bioassay Systems, Hayward, CA) to quantify viability. Briefly, cells were pre-incubated with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) substrate before solubilizing the precipitate (Bioassay Systems). Absorbance was measured at 570 nm in a Multiskan microplate reader (Thermo Scientific, Rockford, IL).
Gene gun vaccination
C57BL/6 mice or C57BL/6-Cd1d1 tm1.1Aben/J (CD1d knockout) mice were gene gun-vaccinated with plasmid DNA encoding adjuvant human inducible heat shock protein 70 (HSP70i) [29] combined 1:1 with plasmid encoding mouse SIAT8 (mSIAT8), human tyrosinase-related protein-1ee/ng (a kind gift from Dr. JA Guevara-Patiño, Loyola University Chicago, Chicago, IL, USA) [30], or empty expression vector DNA. Bullets were prepared using endotoxin-free plasmid DNA precipitated onto spermidine-coated gold beads (Sigma-Aldrich, St. Louis, MO) [31], used to coat silicone tubing (Bio-Rad, Hercules, CA). Hair was removed with Nair (Church and Dwight Company Incorporated, Ewing, NJ). Vaccinations were administered as 4.2 μg of total DNA using a Helios Gene Gun (Bio-Rad, Hercules, CA) under isoflurane anesthesia (E-Z Systems Corporation, Palmer, PA). Mice were vaccinated weekly for a total of 5 times, and tumor challenges were performed after a 9-day interval unless stated otherwise. Animal experiments were performed in accordance with the Guide for the Care and Use of Laboratory Animals (2011).
Adenoviral vectors and vaccination
Recombinant human adenovirus type 5 (hAdV-5) vectors were constructed by inserting human and murine SIAT8 cDNAs into the plasmid pAdapt-CMV. The adaptor plasmids were linearized, digested with PacI and co-transfected with PacI-linearized plasmid pWE/Ad-rITR into PER.C6 helper cells. The hAdV-5 sequences in both plasmids facilitate the generation of recombinant adenovirus through homologous recombination. Upon the appearance of cytopathic effects, cell lysates were prepared by freeze/thawing and used to infect fresh PER.C6 cells. The hAdV-vectors were purified by double CsCl gradient banding and stored at −80 °C. Viral titers were determined by plaque assay on 911 cells [32]. In vitro transduction was performed at 2 × 107 pfu/105 cells for 72 h. For vaccinations, C57BL/6 mice were included at 6 weeks of age and vaccinated with mouse SIAT8-encoding adenovirus (AdV-mSIAT8), empty vector adenovirus (AdV-EV), phosphate-buffered saline (PBS) or 1 µg αGalCer. Mice received 0 or 108 AdV plaque forming units (pfu) intramuscularly in the posterior thigh; αGalCer was administered intraperitoneally. Tissues were harvested after 10 days unless animals were subjected to a subcutaneous B16 melanoma tumor challenge performed 10 days after (prophylactic) or just before (therapeutic) vaccination. In the latter case, a second vaccination was administered on day 7.
Tumor challenge
Mice were challenged with 105 B16F10 melanoma cells (source: American Type Tissue Collection, ATCC), transfected or not with an expression plasmid encoding the SIAT8 enzyme into both flanks. Initial experiments were performed with untransfected GD3+ B16 cells alone. Where SIAT8-transfected cells were used, tumor cells transfected with empty vector alone were introduced into the opposite flank. Tumor growth was monitored daily using calipers for 3 weeks or until tumors reached 1500 mm3. Tumor volume was calculated as 0.5(length × width × height) [33]. Mice were imaged to evaluate depigmentation at euthanasia, and tumor tissues were harvested for further analysis.
FACS analysis
B16 tumors were mechanically homogenized using cell strainers (Fisher Brand, Houston, TX) followed by Lympholyte (Cedarlane Labs, Burlington, NC) separation. Lymphocytes with or without prior phorbol 12-myristate 13-acetate (PMA)/ionomycin treatment were then immunostained using rat antibody 30-F11 to CD45 (BioLegend, San Diego, CA) and rat antibody 6D5 to CD19 (BioLegend) to distinguish non-B cell leukocytes and primary mouse antibody PK136 to NK1.1 (BioLegend, San Diego, CA), hamster antibody H57-597 to TCRβ (Biolegend), rat antibody RM5-5 to mouse CD4 (BD Biosciences, San Jose, CA), rat antibody 53-6.7 to mouse CD8α and hamster antibody 145-2c11 to mouse CD3ε (BD Biosciences) to determine the relative number of T, NKT and NK cell subpopulations. CD3− T cells were gated among NK1.1-positive events to classify NK cells, with the remainder assessed for TCRβ expression and reactivity with αGalCer-loaded dimers (Enzo Life Sciences, Farmingdale, NY and BD Biosciences, respectively) to classify NKT cells or CD4 helper/suppressor T cells or CD8 cytotoxic T cells. CD1d dimer staining adds specificity at the expense of sensitivity, and elsewhere in the manuscript NKT cells are quantified as NK1.1+, CD3+ cells. For intracellular cytokine staining, cells were maintained in fixation/permeabilization buffer (eBioscience, San Diego, CA). Fluorocytometry was performed using Fortessa instrumentation (BD Biosciences) and analyzed using FlowJo software (FlowJo LLC, Ashland, OR).
Immunofluorescent double stains
Optimal cutting temperature (OCT)-embedded mouse skin and tumor samples were cryosectioned (Leica Biosystems, Buffalo Grove, IL), acetone-fixed and stored at −20 °C. After Super Block treatment (ScyTek Laboratories, Inc., West Logan, UT), sections were immunostained using 145-2c11 primary antibodies to CD3ε (BD Biosciences) displayed in red and PK136 to NK1.1 (BD Biosciences) displayed in green. Sections were coverslipped using ProLong Gold antifade reagent with 4′,6-diamidino-2-phenylindole (DAPI) mounting medium (Life Technologies, Grand Island, NY). Cells expressing both markers were counted as NKT cells and counted per microscopic field.
Reverse transcription–quantitative polymerase chain reaction
Ribonucleic acid (RNA) was isolated from 25-µm frozen B16 tissue sections placed in Trizol Reagent (Life Technologies, Grand Island, NY). After sonification, RNA was isolated, and reverse transcription was performed using SuperScriptII enzyme and oligo-dT primers (Life Technologies) at 42 °C for 50 min. Quantitative PCR reactions were set up using SYBR Green (Invitrogen), 200 ng/25 μl reaction cDNA, and commercially optimized qPCR primers (Qiagen, Venlo, NL). Real-time PCR reactions were run on an Applied Biosystems 7500 thermocycler (Life Technologies) following a hot start for 10 min. at 95 °C, and 50 cycles of 95 °C for 15 s, 55 °C for 30 s and 72 °C for 30 s. Data were analyzed using the ΔΔCT method [34].
Statistical analysis
The means of groups were compared using two-sample t tests or ANOVA (when more than two groups were compared), followed by Newman–Keuls multiple comparison test [35]. To adhere to the normality assumption required when comparing mean across groups with small sample sizes, tumor volume and weight data were square-root-transformed to correct for skewness prior to applying hypothesis tests. The results were visually inspected to ensure normality. Statistical analyses were performed using Graphpad Prism software (La Jolla, CA). Statistical significance of the differences in mean between groups is expressed in figures as * p < 0.05; ** p < 0.01; *** p < 0.001.
Results
Introducing GD3 synthase is necessary and sufficient to elicit surface GD3 expression
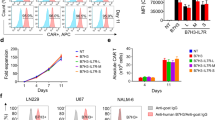
Monosialoganglioside M3 (GM3) is found in most cell types, yet only cells that express GD3 synthase can convert GM3 to GD3. Thus, this enzyme was introduced into GD3− HEK293 cells, and subsequent surface GD3 expression was assessed. In Fig. 1a, expression of GD3 by 93% of cells transfected with hSIAT8 is shown 48 h after transfection. Figure 1b shows complement-dependent cytotoxicity in the presence of the R24 antibody to GD3 was induced in approximately 40% of SIAT8 transfected HEK293 cells within 24 h. An ANOVA indicated significant differences among groups (p < 0.0001). All but the mSIAT8 and hSIAT8 groups differed (p < 0.05) in a Newman-Keuls post-test. In Fig. 1c, GD3− Mus musculus vascular smooth muscle (MOVAS) cells, electroporated to express mouse SIAT8 increased 2.6-fold in mean fluorescence intensity over 2 wks (stained using antibody GD3) relative to cells transduced to express gp100. In Fig. 1d, expression of GD3 on the cell surface of B16 mouse melanoma cells was increased 3.5-fold over background; however, expression of GD3 by B16 cells is variable. Taken together, Fig. 1 shows that SIAT8 leads to GD3 expression and sensitizes cells to CDC.

Expression of GD3 synthase drives membrane GD3 expression. a Human SIAT8 induces GD3 expression by HEK293 cells within 48 h of transfection. b Complement mediated cytotoxicity is observed toward GD3+ HEK293 cells or 624.38 melanoma cells c Mouse SIAT8 electroporated into MOVAS cells likewise leads to overexpression of GD3 d Antibody Mel1 (=R24) to GD3 was used to demonstrate that B16 F10 mouse melanoma cells can express GD3 on the cell surface
Repeated adjuvant-supported SIAT8 gene gun vaccination delays melanoma growth
Overexpression of SIAT8 may be of use to elicit immune responses to the ganglioside that can be protective in a tumor setting. DNA vaccination of mice was performed, followed by tumor growth monitoring upon challenge with B16 melanoma cells transfected with mSIAT8 expression plasmid or empty vector DNA. In supplementary figure S1, SIAT8 transfection mediates elevated expression of GD3 by B16 melanoma cells (Supplementary figure S1a), whereas transfected cells maintain greatly enhanced GD3 expression in vivo (Supplementary figure S1b). In Fig. 2a, mice received plasmids encoding adjuvant HSP70i and either human GD3 synthase, optimized tyrosinase-related protein-1 (TRP-1ee/ng) or empty vector DNA. In Fig. 2b, GD3-overexpressing tumor weights were significantly decreased when comparing the SIAT8 to the negative control group. Meanwhile, in Fig. 2c depigmentation is shown as measured at the time of euthanasia. Significant autoimmunity (depigmentation at the vaccination site) was observed only in the TRP-1ee/ng and HSP70i vaccination group. Thus, protection offered by a DNA vaccine encoding the GD3 synthase enzyme rivals with optimized TRP-1, yet offers the added bonus of anti-tumor protection without autoimmune side effects.

hSIAT8 vaccination significantly delays B16 tumor growth. a C57BL/6 mice were gene gun-vaccinated with mSIAT8 and HSP70i DNA (n = 10), TRP-1ee/ng and HSP70i DNA (n = 5), or HSP70i and empty vector DNA (n = 10). Depigmentation was evaluated at euthanasia. b Tumor growth in mice vaccinated with mSIAT8 and HSP70i was significantly delayed compared to the control group, c Only TRP-1ee/ng and HSP70i vaccination induced significant autoimmune depigmentation as a side effect
Anti-tumor responses in SIAT8 vaccinated mice are CD1d dependent
Immune monitoring was performed on C57BL/6 wildtype and CD1d knockout mice treated with the DNA vaccine. Tumors were harvested at 1.5 cm3 in size or by day 21 after tumor challenge for inclusion in immune monitoring experiments, and partly dissociated to obtain lymphocytes that were subjected to CD45, CD19, CD3, NK1.1, TCRβ and CD1d dimer staining. In Fig. 3a final tumor weights were compared for GD3 overexpressing B16 tumors in wildtype and CD1d knockout mice treated with or without SIAT8 vaccination. As assessed by ANOVA, tumor sizes were significantly different within the C57BL/6 group, and reduced GD3++ tumor sizes were observed for SIAT8 treated C57BL/6 mice but not for CD1d knockout mice. Taken together with Fig. 2, the data support a role for NKT cell-mediated anti-tumor responses to GD3 s vaccination. In Fig. 3b, detection of NKT cells as αGalactosylCeramide (αGalCer)-loaded dimer engaging TCR-β+ cells among the CD45+, CD19− population of lymphocytes is compared to background using unloaded dimers (top panels). Similar analysis applied to a GD3 overexpressing tumor (+ in the table under GD3++) and its contralateral counterpart (− in the table) suggests that NKT cells preferentially home to GD3 overexpressing tumors (lower panels). In Fig. 3c, where NK populations are quantified, or in 3d where T cells were quantified, relative infiltration by these subpopulations did not differ based on GD3 overexpression, presence or absence of CD1d or vaccination strategy. In 3e, NKT cell infiltration is clearly dependent on the presence of CD1d and trends toward an increase in response to GD3 overexpression or SIAT8 vaccination. In situ staining reveals increased intra-tumoral abundance of NKT cells in response to the SIAT8 vaccine in Fig. 3f. Taken together, the data support a role for NKT cells as the main mechanism of response to SIAT8 vaccination.

Efficacy of mSIAT8 vaccination is CD1d dependent. a Contrary to C57BL/6 mice, in CD1d knockout mice, final GD3++ tumor weights were not reduced by SIAT8 vaccination. b NKT cells are detected using loaded (dimers + αGalCer), but not unloaded (unloaded dimers) dimers (top panels) and such NKT cells are generally more abundant in GD3++ tumors than in GD3+ tumors (+ and − in the table, respectively; lower panels). c NK cell and d T cell abundance is independent of elevated GD3 expression, SIAT8 vaccination and the presence or absence of CD1d, whereas e NKT cells are found only in tumors from CD1d expressing mice and trend toward an increase in response to GD3 overexpression and SIAT8 vaccination, as solidified by f immunofluorescent in situ staining of NK1.1+ (green) and CD3+ (red) staining to reveal NKT cells (yellow)
Adenoviral transduction rapidly induces vast GD3 overexpression in GD3− cells
We sought to adjust the strategy to a safe and effective method by means of an adenoviral vector. For proof of principle, SIAT8 was transduced into MOVAS cells. In Fig. 4a, expression of GD3 synthase was measured 72 h post-transduction, when the mean fluorescence intensity for GD3 synthase staining was approximately 17-fold increased over MOVAS cells transduced with AdV-EV. In Fig. 4b, an estimated 450-fold increase in GD3 expression is observed in the cells. Thus, an adenoviral vector is increasingly efficient when compared to naked DNA. Thus, adenoviral vaccination was next tested to introduce SIAT8 in vivo.

Adenoviral SIAT8 transfection leads to rapidly increased GD3 expression in MOVAS cells. MOVAS cells were transduced with adenovirus encoding human or mouse SIAT8 before a intracellular staining and FACS analysis of SIAT8 expression (antibody K-18) or b extracellular staining and FACS analysis of high levels of surface GD3 expression by the majority of cells
SIAT8 vaccination induces measurable changes in immune parameters
Adenoviral vaccination might model the effects of SIAT8 and GD3 overexpression in mice. As repeated vaccination raises a concern for pre-existing immunity toward the virus in patients, such repeats were limited. Mice were intramuscularly injected (αGalCer was introduced intraperitoneally), and injection sites and spleens were subjected to immune monitoring after 10 days. In Fig. 5a, reduced NKT cell numbers among splenocytes were measured in mice treated with AdV-mSIAT8 or with αGalCer as compared to control groups, suggesting that NKT cells were recruited elsewhere. NKT cell frequencies did not increase significantly after in vitro PMA/ionomycin stimulation. In Fig. 5b, a trend toward increased IL-4 production is observed among splenocytes after PMA/ionomycin stimulation in the mSIAT8- and the αGalCer-treated groups. A trend toward PMA/ionomycin-induced expression of IFN-γ was only observed in the αGalCer group (not shown). In Fig. 5c, injection sites were probed for NKT cells. Among injected sites, NKT cells were sparse (yellow arrows) and found only in the AdV-mSIAT8 group. In Fig. 5d, a trend toward increased NKT cell infiltration is observed at the injection site (n = 4); the biopsy site may not fully coincide with the injection site. Enumerated NK1.1+CD3+ cells may consist in part of activated T cells responding to AdV injection, though they were observed at the injection site only in response to SIAT8. It thus appears that a single adenoviral vaccination to overexpress SIAT8 and, consequently, GD3 will elicit demonstrable NKT cell responses.

Mobilization of NKT cell responses by AdV-SIAT8 administration in vivo. Mice (n = 4) were injected with murine SIAT8 encoding or empty vector AdV or PBS, or IP injected with αGalCer before harvesting spleens and the injection site. a NKT cells are underrepresented in spleens from mice treated with αGalCer or with AdVmSIAT8 supporting selective recruitment, as shown for blood-borne antigens by others [49], b NKT cells among splenocytes from αGalCer treated and AdVmSIAT8 trended slightly toward greater IL-4 production per cell upon activation. c NKT cells selectively infiltrate the injection site in AdVmSIAT8-treated mice (yellow arrows), but not in EV- or PBS-treated mice (red and green arrows: NK cells and T cells, respectively). d A trend toward increased NKT cell infiltration was observed among AdV-SIAT8 injected sites (n = 4)
A single adenoviral injection to introduce SIAT8 delays tumor growth
AdV vaccination was performed for mice that were subsequently challenged with B16 melanoma cells. Whereas therapeutic vaccination of rapidly growing B16 tumors was not effective (supplementary figure S3), prophylactic treatment did reduce tumor growth. As shown in Fig. 6a, mice (n = 10) were intramuscularly injected with 108 pfu of AdV-mSIAT8 or not. A bilateral B16 tumor challenge was followed after 10 days, and tumor growth was followed for 18 days. In Fig. 6b, delayed tumor growth is depicted (p < 0.01) compared to the control group. In Fig. 6c, this difference is upheld when comparing tumor weights. In supplementary figure S2, tumors from the AdV-EV-vaccinated group expressed more transcripts encoding IL-10 (2.3-fold), IL-4 (2.2-fold) and IFN-γ (7.4-fold) compared to control animals, with no net effect on tumor growth. However, tumors from AdV-mSIAT8-vaccinated animals also displayed increased IFN-γ and only in this group IL-17A was increased compared to PBS injection (n = 4). Thus, a single adenoviral SIAT8 vaccination can lead to immune activation and reduced tumor growth. This provides support for a therapeutic enzyme vaccination strategy to safely overexpress GD3 and elicit responses to GD3+ tumors.

A single AdV vaccination delays tumor growth in response to AdV-hSIAT8 a Adenoviral vaccination scheme. C57BL/6 mice (n = 10) were subjected to vaccination with AdV-mSIAT8, or AdV-empty vector, or PBS. b Final tumor sizes measured post-challenge were reduced for the AdV-SIAT8 group. c Final tumor weights were likewise reduced in AdV-hSIAT8 mice
Discussion
An interest in targeting GD3 as a tumor antigen can be founded in the opportunity to elicit responses not only against melanoma and small cell lung cancer, but also toward tumors with a more benign physiology such as those observed in lymphangioleiomyomatosis (LAM) [36]. As patients develop LAM due to mutations that enhance GD3 expression, overexpression of this target is consistent [37]. However, patients with LAM do not exhibit spontaneous immune responses to the ganglioside [7]. This further prompted the objective to target GD3.
Overexpressing the enzyme responsible for generating GD3 from GM3 was sufficient to drive surface expression of the disialoganglioside, consistent with studies demonstrating that SIAT8 expression limits GD3 expression [38]. Consequently, expression of SIAT8 may be a prognostic factor in cancer [39]. The remarkably accelerated tumor growth observed among mSIAT8-transduced, GD3++ B16 cells further supports this. Thus, providing the enzyme together with an adjuvant might be sufficient to elicit an immune response to GD3. An important advantage is that the tolerizing consequences of immunizing with the ganglioside itself can be circumvented [40], as observed in response to circulating GD3 shed by ovarian tumors; as melanomas reportedly shed GD3 as well, upregulated expression introduced by SIAT8-encoding DNA may help to overcome such tolerization [41].
Available antibodies to GD3 are generally of the IgG3 isotype. If this reflects the humoral response to naturally occurring antigen, resulting antibodies could bind complement and elicit effective cytotoxic responses. In LAM and in melanoma, humoral responses to GD3 are sparse or absent [7]. Such a shortage can possibly be remedied by SIAT8 vaccination. Under conditions where GD3 is overexpressed, abundant infiltration by macrophages and granulocytes has been reported. Thus, once humoral responses GD3 exist, tumors can likewise be targeted by antibody-dependent cellular cytotoxicity (ADCC). Importantly, antibodies to gangliosides can be cytotoxic by inducing cell detachment and other morphologic changes [42].
Though HSP70i plus SIAT8 gene gun vaccination did not affect NK recruitment, interactions between GD3 with surface siglec-7 may contribute to NK activation and the betterment of anti-tumor responses [43]. Cytotoxic T-lymphocyte (CTL) activation is likewise a feature of αGalCer vaccination [44]. It is possible that CTL were reactive to the SIAT8 enzyme itself. Even so, such responses can be supportive of tumor elimination as well.
Murine SIAT8 was next tested as an adenoviral vaccine. The enzyme was injected with AdV acting as an immune adjuvant [26]. The adenovirus was initially tested in 911 host cells, mediating expression of both the enzyme and GD3 (not shown) [32]. This was next assessed in MOVAS host cells [45] to alleviate possible concerns with infecting murine cells [46], and convincing expression of both enzyme and product was observed, providing incentive to further test adenoviral vaccination in mice.
Most importantly, SIAT8 vaccination appears to elicit NKT cell recruitment to the tumor. NKT cells were not found in tumors from CD1d knockouts where the vaccine failed to offer protection, indicating the latter is a potent strategy to elicit NKT cell responses other than administering αGalCer.
As repeated AdV vaccination may be thwarted in humans due to pre-existing immunity neutralizing incoming virions, we instead selected a single vaccination rapidly followed by a bilateral tumor challenge [47]. Delayed tumor growth was detected as reduced volumes and tumor weights. Complete remissions were not observed, and our studies indicate that SIAT8, particularly when therapeutically applied, will require combination therapy to treat aggressive tumors such as melanomas with a narrow treatment window. That said, different vaccination doses or additional adjuvants may further enhance this NKT cell-based strategy.
It appears that αGalCer vaccination elicits a dual IFN-γ- and IL-4-based response by NKT cells [48]. The SIAT8 vaccine, however, appears to accentuate a more IL-4-based response. As αGalCer is not expressed by mouse or human cells, its xenogeneity may explain this bias. However, even SIAT8 vaccination appears to be accompanied by NKT cell recruitment to the vaccination site. Benefit gained from SIAT8 vaccination may thus be ascribed to either enhanced recruitment of existing iNKT cells or to induction of memory NKT cells.
The effects of adenoviral vaccination on tumor growth associated with enhanced IL-17A and IFN-γ expression suggest that responsive cells generate supportive cytokines on site, where GD3 is in ample supply. Enhanced IL-17A expression may point to mTOR activation blocking invariant NKT cell (iNKT) differentiation from iNKT-17 to iNKT-1 [49]. Though expression by B16 melanoma cells is low [50], some tumors can express CD1d in which case, NKT cells can directly respond to GD3 presented by tumors [51]. In fact, the requirement for CD1d to mediate NKT cell activation in response to GD3 was firmly established [20, 51]. This can explain cytokine abundance and differences in tumor size.
Thus vaccinations based on expression of an enzyme responsible for de novo expression of a tumor antigen can offer a new concept to elicit prophylactic anti-tumor responses, whereas established tumors are less amenable to treatment. Prophylaxis is relevant for diseases associated with loss of TSC function and concomitant GD3 upregulation [7]. Both DNA and adenoviral vaccination are processed by dendritic cells [52], and the observed responses to SIAT8 vaccination in the tumor appear to be primarily NKT cell-based.
While immunotherapy targeting protein antigens can be associated with autoimmune side effects such as depigmentation, SIAT8 did not elicit a similar loss of pigment, even though GD3 is also expressed by melanocytes. This discrepancy is best explained by melanocytes not necessarily colocalizing with CD1d expressing dendritic cells, so that NKT cells will unlikely mediate cytotoxicity toward melanocytes.
The current strategy may be of use to support adoptive T cell therapy with chimeric antigen-receptor-transduced T cells targeting GD3 by generating a favorable anti-tumor cytokine environment, whereas SIAT8-based vaccination itself is more suited for prophylactic applications, for example in familial melanoma. The effects observed support the concept that adenoviral SIAT8 vaccines can interfere with growth of GD3 expressing tumors, without requiring personalized vaccine development as used in dendritic cell-based vaccines to administer αGalCer [3, 53, 54]. Treatment can be directed toward particular locations, and overexpression of the target antigen is renewable and tenable over a longer period of time. Also, there is potential benefit to be gained from supporting humoral responses to a membrane antigen such as GD3.
Abbreviations
- ADCC:
-
Antibody-dependent cellular cytotoxicity
- AdV:
-
Adenovirus
- αGalCer:
-
Alpha galactosylceramide
- ANOVA:
-
Analysis of variance
- ATCC:
-
American Type Tissue Collection
- CD:
-
Cluster of differentiation
- CDC:
-
Complement-dependent cytotoxicity
- CTL:
-
Cytotoxic T-lymphocyte
- DAPI:
-
4′,6-Diamidino-2-phenylindole
- DMEM:
-
Dulbecco’s modified Eagle’s medium
- DNA:
-
Deoxyribonucleic acid
- EV:
-
Empty vector
- GD3:
-
Disialoganglioside 3
- GM3:
-
Monosialoganglioside 3
- gp100:
-
100 kD Glycoprotein
- HEK293 cells:
-
Human embryonic kidney 293 cells
- HSP:
-
Heat shock protein
- IFN:
-
Interferon
- IL:
-
Interleukin
- iNKT cell:
-
Invariant natural killer T cell
- LAM:
-
Lymphangioleiomyomatosis
- MOVAS cells:
-
Mus musculus vascular smooth muscle cells
- MTT:
-
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- OCT:
-
Optimal cutting temperature
- PBS:
-
Phosphate-buffered saline
- pfu:
-
Plaque forming units
- PMA:
-
Phorbol 12-myristate 13-acetate
- qRT-PCR:
-
Quantitative reverse transcription-polymerase chain reaction
- RNA:
-
Ribonucleic acid
- SCLC:
-
Small cell lung cancer
- SIAT8:
-
Alpha-N-acetyl-neuraminide alpha-2,8-sialyltransferase 1
- TRP:
-
Tyrosinase-related protein
References
Matarrese P, Garofalo T, Manganelli V, Gambardella L, Marconi M, Grasso M, Tinari A, Misasi R, Malorni W, Sorice M (2014) Evidence for the involvement of GD3 ganglioside in autophagosome formation and maturation. Autophagy 10:750–765. doi:10.4161/auto.27959
Wang XQ, Sun P, Paller AS (2002) Ganglioside modulation regulates epithelial cell adhesion and spreading via ganglioside-specific effects on signaling. J Biol Chem 277:40410–40419
Motohashi S, Nagoto K, Kunii N, Yamamoto H, Yamasaki K, Okita K, Hanaoka H, Shimizu N, Suzuki M, Yoshino I, Tanaguchi M, Fujisawa T, Nakayama T (2009) A phase I–II study of alpha-galactosylceramide-pulsed IL-2/GM-CSF-cultured peripheral blood mononuclear cells in patients with advanced and recurrent non-small cell lung cancer. J Immunol 182:2492–2501. doi:10.4049/jimmunol.0800126
Spessott W, Crespo PM, Daniotti JL, Maccioni HJ (2012) Glycosyltransferase complexes improve glycolipid synthesis. FEBS Lett 586:2346–2350. doi:10.1016/j.febslet.2012.05.041
Daniotti JL, Martina JA, Zurita AR, Maccioni HJ (1999) Mouse beta 1,3-galactosyltransferase (GA1/GM1/GD1b synthase): protein characterization, tissue expression, and developmental regulation in neural retina. J Neurosci Res 58:318–327
Ariga T, Suetake K, Nakane M, Kubota M, Usuki S, Kawashima I, Yu RK (2008) Glycosphingolipid antigens in neural tumor cell lines and anti-glycosphingolipid antibodies in sera of patients with neural tumors. Neurosignals 16:226–234. doi:10.1159/000111565
Gilbert ER, Eby JM, Hammer AM, Klarquist J, Christensen DG, Barfuss AJ, Boissy RE, Picken MM, Love RB, Dilling DF, Le Poole IC (2013) Positioning ganglioside D3 as an immunotherapeutic target in lymphangioleiomyomatosis. Am J Pathol 183:226–234. doi:10.1016/j.ajpath.2013.04.002
Birklé S, Zeng G, Gao L, Yu RK, Aubry J (2003) Role of tumor-associated gangliosides in cancer progression. Biochimie 85:455–463
Tomassini B, Malisan F, Franchi L, Nicolò C, Calvo GB, Saito T, Testi R (2004) Calnexin suppresses GD3 synthase-induced apoptosis. FASEB J 18:1553–1555
Chen HY, Challa AK, Varki A (2006) 9-O-acetylation of exogenously added ganglioside GD3. The GD3 molecule induces its own O-acetylation machinery. J Biol Chem 281:7825–7833
Streilein JW, Ma N, Wenkel H, Ng TF, Zamiri P (2002) Immunobiology and privilege of neuronal retina and pigment epithelium transplants. Vision Res 42:487–495
Houghton AN, Bajorin DF, Lonberg M, Chapman P (1989) Treatment of human metastatic melanoma with mouse monoclonal antibodies against GD3 ganglioside. Prog Clin Biol Res 288:383–390
Zhan J, Han Q, Wang K (2013) Development of antibody therapeutics for small cell lung cancer. Expert Opin Investig Drugs 22:235–244. doi:10.1517/13543784.2013.750293
Bottomley A, Debruyne C, Felip E, Millward M, Thiberville L, D’Addario G, Rome L, Zatloukal P, Coens C, Giaccone G (2008) Symptom and quality of life results of an international randomised phase III study of adjuvant vaccination with Bec2/BCG in responding patients with limited disease small-cell lung cancer. Eur J Cancer 44:2178–2184. doi:10.1016/j.ejca.2008.06.036
Stevenson GT (2014) Three major uncertainties in the antibody therapy of cancer. Haematologica 99:1538–1546. doi:10.3324/haematol.2013.084640
Byers LA, Rudin CM (2015) Small cell lung cancer: where do we go from here? Cancer 121:664–672. doi:10.1002/cncr.29098
Lo AS, Ma Q, Liu DL, Junghans RP (2010) Anti-GD3 chimeric sFv-CD28/T-cell receptor zeta designer T cells for treatment of metastatic melanoma and other neuroectodermal tumors. Clin Cancer Res 16:2769–2780. doi:10.1158/1078-0432.CCR-10-0043
Robertson FC, Berzofsky JA, Terabe M (2014) NKT cell networks in the regulation of tumor immunity. Front Immunol 5:543. doi:10.3389/fimmu.2014.00543
Wingender G, Krebs P, Beutler B, Kronenberg M (2010) Antigen-specific cytotoxicity by invariant NKT cells in vivo is CD95/CD178-dependent and is correlated with antigenic potency. J Immunol 185:2721–2729. doi:10.4049/jimmunol.1001018
Park JE, Wu DY, Lu SX, Raghupathi G, Schrantz N, Chapman PB (2008) Fine specificity of natural killer T cells against GD3 ganglioside and identification of GM3 as an inhibitory natural killer T-cell ligand. Immunology 123:145–155
Terabe M, Berzofsky JA (2008) The role of NKT cells in tumor immunity. Adv Cancer Res 101:277–348. doi:10.1016/S0065-230X(08)00408-9
Chong TW, Goh FY, Sim MY, Huang HH, Thike DA, Lim WK, The BT, Tan PH (2015) CD1d expression in renal cell carcinoma is associated with higher relapse rates, poorer cancer-specific and overall survival. J Clin Pathol 68:200–205. doi:10.1136/jclinpath-2014-202735
Fiedler T, Walter W, Reichert TE, Maeurer MJ (2002) Regulation of CD1d expression by murine tumor cells: escape from immunosurveillance or alternate target molecules? Int J Cancer 98:389–397
Wen X, Rao P, Carreño LJ, Kim S, Lawrenczyk A, Porcelli SA, Cresswell P, Yuan W (2013) Human CD1d knock-in mouse model demonstrates potent antitumor potential of human CD1d-restricted invariant natural killer T cells. Proc Natl Acad Sci U S A 110:2963–2968. doi:10.1073/pnas.1300200110
Sandberg JK, Bhardwaj N, Nixon DF (2003) Dominant effector memory characteristics, capacity for dynamic adaptive expansion, and sex bias in the innate Valpha24 NKT cell compartment. Eur J Immunol 33:588–596
Schumacher L, Ribas A, Dissette VB, McBride WH, Mukherji B, Economou JS, Butterfield LH (2004) Human dendritic cell maturation by adenovirus transduction enhances tumor antigen-specific T-cell responses. J Immunother 27:191–200
Xiang K, Ying G, Yan Z, Shanshan Y, Lei Z, Hongjun L, Maosheng S (2015) Progress on adenovirus-vectored universal influenza vaccines. Hum Vaccin Immunother 11:1209–1222. doi:10.1080/21645515.2015.1016674
Gao Q, Chen C, Ji T, Wu P, Han Z, Fang H, Li F, Liu Y, Hu W, Gong D, Zhang Z, Wang S, Zhou J, Ma D (2014) A systematic comparison of the anti-tumoural activity and toxicity of the three Adv-TKs. PLoS ONE 9:e94050. doi:10.1371/journal.pone.0094050
Mosenson JA, Zloza A, Nieland JD, Garrett-Mayer E, Eby JM, Huelsmann EJ, Kumar P, Denman CJ, Lacek AT, Kohlhapp FJ, Alamiri A, Hughes T, Bines SD, Kaufman HL, Overbeck A, Mehrotra S, Hernandez C, Nishimura MI, Guevara-Patino JA, Le Poole IC (2013) Mutant HSP70 reverses autoimmune depigmentation in vitiligo. Sci Transl Med. 2013 Feb 27;5(174):174ra28. doi: 0.1126/scitranslmed.3005127
Guevara-Patiño JA, Engelhorn ME, Turk MJ, Liu C, Duan F, Rizzuto G, Cohen AD, Merghoub T, Wolchok JD, Houghton AN (2006) Optimization of a self antigen for presentation of multiple epitopes in cancer immunity. J Clin Invest 116:1382–1390
Denman CJ, McCracken J, Hariharan V, Klarquist J, Oyarbide-Valencia K, Guevara-Patiño JA, Le Poole IC (2008) HSP70i accelerates depigmentation in a mouse model of autoimmune vitiligo. J Invest Dermatol 128:2041–2048. doi:10.1038/jid.2008.45
Fallaux FJ, Kranenburg O, Cramer SJ, Houweling A, Van Ormondt H, Hoeben RC, Van Der Eb AJ (1996) Characterization of 911: a new helper cell line for the titration and propagation of early region 1-deleted adenoviral vectors. Hum Gene Ther 7:215–222
Tomayko MM, Reynolds CP (1989) Determination of subcutaneous tumor size in athymic (nude) mice. Cancer Chemother Pharmacol 24:148–154
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2 −∆∆CT method. Methods 25:402–408
Sokal RR, Rohlf FJ (1994) Biometry: the principles and practices of statistics in biological research, 3rd edn. W.H. Freeman and Company, New York
Dilling DF, Gilbert ER, Picken MM, Eby JM, Love RB, Le Poole IC (2012) A current viewpoint of lymphangioleiomyomatosis supporting immunotherapeutic treatment options. Am J Respir Cell Mol Biol 46:1–5. doi:10.1165/rcmb.2011-0215TR
Yamauchi Y, Furukawa K, Hamamura K, Furukawa K (2011) Postive feedback loop between PI3K-Akt-mTORC1 signalling and the lipogenic pathway boost Akt signaling: induction of the lipogenic pathway by a melanoma antigen. Cancer Res 71:4989–4997. doi:10.1158/0008-5472.CAN-10-4108
Ruan S, Raj BK, Lloyd KO (1999) Relationship of glycosyltransferases and mRNA levels to ganglioside expression in neuroblastoma and melanoma cells. J Neurochem 72:514–521
Ruckhäberle E, Karn T, Rody A, Hanker L, Gätje R, Metzler D, Holtrich U, Kaufmann M (2009) Gene expression of ceramide kinase, galactosyl ceramide synthase and ganglioside GD3 synthase is associated with prognosis in breast cancer. J Cancer Res Clin Oncol 135:1005–1013. doi:10.1007/s00432-008-0536-6
Webb TJ, Li X, Giuntoli RL, Lopez PH, Heuser C, Schnaar RL, Tsuji M, Kurts C, Oelke M, Schneck JP (2012) Molecular identification of GD3 as a suppressor of the innate immune response in ovarian cancer. Cancer Res 72:3744–3752. doi:10.1158/0008-5472.CAN-11-2695
Sela BA, Iliopoulos D, Guerry D, Herlyn D, Koprowski H (1989) Levels of disialogangliosides in sera of melanoma patients monitored by sensitive thin-layer chromatography and immunostaining. J Natl Cancer Inst 81:1489–1492
Horwacik I, Rokita H (2015) Targeting of tumor-associated gangliosides with antibodies affects signaling pathways and leads to cell death including apoptosis. Apoptosis 20:679–688. doi:10.1007/s10495
Nicoll G, Avril T, Lock K, Furukawa K, Bovin N, Crocker PR (2003) Ganglioside GD3 expression on target cells can modulate NK cell cytotoxicity via siglec-7-dependent and –independent mechanisms. Eur J Immunol 33:1642–1648
Hermans IF, Silk JD, Gileadi U, Salio M, Mathew B, Ritter G, Schmidt R, Harris AL, Old L, Cerundolo V (2003) NKT cells enhance CD4+ and CD8+ T cell responses to soluble antigen in vivo through direct interaction with dendritic cells. J Immunol 171:5140–5147
Mackenzie NC, Zhu D, Longley L, Patterson CS, Kommareddy S, MacRae VE (2011) MOVAS-1 cell line: a new in vitro model of vascular calcification. Int J Mol Med 27:663–668. doi:10.3892/ijmm.2011.631
Clausen BE, Brand A, Karram K (2015) Surmounting limited gene delivery into primary immune cell populations: efficient cell-type-specific adenoviral transduction by CAR. Eur J Immunol 45:1596–1599. doi:10.1002/eji.201545685
Wan Y, Emtage P, Foley R, Carter R, Gauldie J (1999) Murine dendritic cells transduced with an adenoviral vector expressing a defined tumor antigen can overcome anti-adenovirus neutralizing immunity and induce effective tumor regression. Int J Oncol 14:771–776
Chen Q, Ross AC (2015) Α-Galactosylceramide stimulates splenic lymphocyte proliferation in vitro and increases antibody production in vivo in late neonatal-age mice. Clin Exp Immunol 179:188–196. doi:10.1111/cei.12447
Zhang L, Tschumi BO, Corgnac S, Ruegg MA, Hall MN, Mach JP, Romero P, Donda A (2014) Mammalian target of rapamycin complex 1 orchestrates invariant NKT cell differentiation and effector function. J Immunol 193:1759–1765. doi:10.4049/jimmunol.1400769
Shimizu K, Goto A, Fukui M, Taniguchi M, Fujii S (2007) Tumor cells loaded with alpha-galactosylceramide induce innate NKT and NK cell-dependent resitance to tumor implantation in mice. J Immunol 178:2853–2861
Wu DY, Segal NH, Sidobre S, Kronenberg M, Chapman PB (2003) Cross-presentation of disialoganglioside GD3 to natural killer T cells. J Exp Med 198:173–181
Ada G, Ramshaw I (2003) DNA vaccination. Expert Opin Emerg Drugs 8:27–35
Kurosaki M, Horiguchi S, Yamasaki K, Uchida Y, Motohashi S, Nakayama T, Sugimoto A, Okamoto Y (2011) Migration and immunological reaction after the administration of αGalCer-pulsed antigen-presenting cells into the submucosa of patients with head and neck cancer. Cancer Immunol Immunother 60:207–215. doi:10.1007/s00262-010-0932-z
Barral P, Sanchez-Nino MD, van Rooijen N, Cerundolo V, Batista FD (2012) The location of splenic NKT cells favours their rapid activation by blood-borne antigen. EMBO J 31:2378–2390. doi:10.1038/emboj.2012.87
Acknowledgements
Support by a Department of Defense Idea Development Award TS110036 to I. Caroline Le Poole, Ph.D. is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest to report.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Eby, J.M., Barse, L., Henning, S.W. et al. Alpha-N-acetyl-neuraminide alpha-2,8-sialyltransferase 1 can support immune responses toward tumors overexpressing ganglioside D3 in mice. Cancer Immunol Immunother 66, 63–75 (2017). https://doi.org/10.1007/s00262-016-1920-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00262-016-1920-8