
Abstract
Drug-induced liver injury (DILI) constitutes one of the most frequent reasons of restricted-use warnings as well as withdrawals of drugs in postmarketing and poses an important concern for the pharmaceutical industry. The current hepatic in vivo and in vitro models for DILI detection have shown clear limitations, mainly for studies of long-term hepatotoxicity. For this reason, we here evaluated the potential of using Upcytes human hepatocytes (UHH) for repeated-dose long-term exposure to drugs. The UHH were incubated with 15 toxic and non-toxic compounds for up to 21 days using a repeated-dose approach, and, in addition to conventional examination of effects on viability, the mechanisms implicated in cell toxicity were also assessed by means of high-content screening. The UHH maintained the expression and activity levels of drug-metabolizing enzymes for up to 21 days of culture and became more sensitive to the toxic compounds after extended exposures, showing inter-donor differences which would reflect variability among the population. The assay also allowed to detect the main mechanisms implicated in the toxicity of each drug as well as identifying special susceptibilities depending on the donor. UHH can be used for a long-term repeated detection of DILI at clinically relevant concentrations and also offers key mechanistic features of drug-induced hepatotoxicity. This system is therefore a promising tool in preclinical testing of human relevance that could help to reduce and/or replace animal testing for drug adverse effects.
Similar content being viewed by others

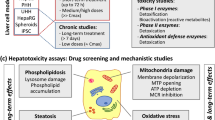

Introduction
Drug-induced liver injury (DILI) is a serious concern for clinicians, pharmaceutical industry and regulatory authorities. Hepatotoxicity is a major cause of acute liver failure and a leading reason for attrition of drug candidates and post-marketing black box warnings or drug withdrawals (Bjornsson 2015; Reuben et al. 2010). Safety assessment of new drugs is essential for the pharmaceutical development process, but represents a major bottleneck as high attrition rates reveal the poor predictive capacity of preclinical screenings. Known interspecies differences in drug metabolism, pharmacokinetics and toxicity targets as well as the increasing pressure to use animal-free models have greatly encouraged the development of human hepatic cell-based models for hepatotoxicity predictions (Prieto et al. 2006; Vanhaecke et al. 2011). For acute toxicity risk assessments, a wide variety of in vitro assays amenable to high-throughput applications are already available (Funk and Roth 2017; Gomez-Lechon and Tolosa 2016; Gomez-Lechon et al. 2014), but animal testing is still largely used to assess long-term effects of drugs. Despite the potential limitations of cell models, current legislation strongly promotes their application to predict chronic hepatotoxicity, which results in the need for smaller amounts of test compounds, accelerated testing and reduced costs (Prieto et al. 2006; Vanhaecke et al. 2011).
Primary human hepatocytes (PHH) are the gold standard in vitro model system for studying liver biology and function as well as for hepatotoxicity studies (Gomez-Lechon et al. 2014); however, when maintained in conventional 2D monolayer cultures, PHH de-differentiate and rapidly lose hepatocyte-specific functions, which severely limits their use for long-term studies. Alternatively, 3D cultures have been successfully proposed to prolong differentiated phenotype of hepatocytes enabling long-term toxicity predictions (Bell et al. 2017; Vorrink et al. 2018); however, further optimization is required to increase throughput testing. Moreover, the availability of fresh PHH is highly limited in contrast to established cell lines. Recently, alternative human hepatic cell models have been proposed for long-term toxicity studies after repeated-dosing (Kramer et al. 2015). Although the HepaRG hepatocarcinoma cell line exhibits a prolonged expression of hepatic functionality, the time-consuming differentiation procedure may be a major drawback (Broeders et al. 2015; Kramer et al. 2015; Mueller et al. 2014). On the other hand, human pluripotent stem cell-derived hepatocytes are a stable and unlimited available source of hepatic cells, but end stage differentiation to mature cells is not fully reached yet (Holmgren et al. 2014). More recently, Upcyte technology has provided the generation of easy handling human hepatocytes that present functional features combined with the advantage of being derived on a large scale, which implies virtually unlimited availability of hepatocytes from different donors (Burkard et al. 2012; Levy et al. 2015; Norenberg et al. 2013; Sison-Young et al. 2015; Tolosa et al. 2016b). Their potential suitability for preclinical drug metabolism and for hepatotoxicity assessments has also been recently shown (Tolosa et al. 2016b).
In this study, we evaluated the suitability of functionally characterized Upcyte human hepatocytes (UHH) from three different donors to investigate hepatotoxicity in response to repeated-dose drug exposure. Compounds with well-documented in vivo hepatotoxicity through different mechanisms of hepatotoxicity were screened in UHH at low concentrations comparable to the plasma levels reached during therapeutic use, which have a clinical relevance. To this end, the evaluation of a battery of pre-lethal toxicity endpoints and acute and repeated-dose assays up to 3 weeks for drug hepatotoxicity testing were performed by multiparametric high-content screening (HCS) analyses. Combined, our data indicate the suitability of UHH for long-term repeated-dose studies that can detect the potential hepatotoxicity of drug candidates in humans.
Materials and methods
Cell cultures
Cryopreserved UHH from three different female Caucasian donors (donor 1: 43 years old; donor 2: 9 years old; donor 3: 48 years old) were obtained from Medicyte (Heidelberg, Germany) and cultured as previously described (Tolosa et al. 2016b). UHH were seeded in 96-well plates (1.2 × 104 cells/well) for HCS assays or in 24-well plates (1.3 × 105 cells/well) for the mRNA and activity measurements. Each point of the measurements is indicated in Fig. 1.

Scheme of experimental procedure. UHH from three donors were seeded in 96-well plates for HCS assays or in 24-well for the mRNA and activity measurements and pre-cultured for 3 days. For the mRNA expression and activity measurements, five time points of control cells were collected and determined. For the toxicity studies, cells were exposed to tested drugs for different time points: one single-dose exposure for 24 h (Day 1) and repeated-dose exposures for 3, 7, 14 or 21 days (two, four, seven or ten repeated doses, respectively)
Undifferentiated HepaRG cells (Biopredic International, Rennes, France) were seeded at a density of 2 × 104 cells/cm2 and were cultured for two weeks as previously described (Tolosa et al. 2016a). Then, cells were differentiated by culturing for another 2-week period in the same medium supplemented with 2% DMSO. Cells were collected for mRNA and activity measurements immediately after the differentiating period (day 0), and 7 and 14 days after differentiation.
HepG2 cells were seeded at a density of 1.5 × 104 cells/cm2 and cultured as previously described (Tolosa et al. 2016a). Cells were allowed to grow until 80% confluence and then collected for mRNA and activity measurements.
Transcriptomic and functional characterization of UHH
mRNA expression, drug-metabolizing activities and albumin secretion were determined in UHH after several days of culture as described in detail in supplementary information.
Selection of drugs and treatments
Eleven drugs with previous reports of hepatotoxicity (amiodarone, azathioprine, chlorpromazine, flutamide, isoniazid, ketoconazole, ketotifen, methotrexate, simvastatin, tacrine and valproate) were included in the study (Supplementary Table S1). These drugs belong to different therapeutic groups and are commonly used in chronic or long-term treatments. In addition, four non-hepatotoxic drugs (betaine, dexamethasone, isoproterenol, streptomycin) were selected as negative controls. For each drug, three concentrations, corresponding to the Cmax (therapeutic peak plasmatic concentration), 0.2 × Cmax and 5 × Cmax, were used. Information on mechanisms of toxicity, therapeutic class, Cmax and concentrations used are summarized in Table S1.
Cells were exposed to tested drugs for different time points: one single-dose exposure for 24 h (Day 1) and repeated-dose exposures for 3, 7, 14 or 21 days (two, four, seven or ten repeated doses, respectively). The scheme of treatment is represented in Fig. 1. The stock solutions of drugs were prepared in DMSO or water and were freshly diluted in the culture medium to obtain the desired final concentrations. The final concentration of DMSO in the culture medium never exceeded 0.5% (v/v) and control cultures were treated with the same amount of solvent.
HCS assay for toxicity assessment
Following treatments, cells were simultaneously loaded with several fluorescent dyes to measure multiple biomarkers of cell toxicity. Different combinations of fluorescent probes were used to identify specific mechanisms of toxicity according to previously described HCS assays (Donato et al. 2012; Tolosa et al. 2012, 2015). Information about the probes is summarized in Supplementary Table S2.
After incubating with dyes, cells were imaged by INCELL6000 (GE Healthcare, Barcelona, Spain) and the collected images were analyzed using the INCELL Workstation analysis module (Tolosa et al. 2012). Representative images of treated cells over time are provided in Supplementary Fig. 1S.
Data analysis and interpretation
The minimal effective concentration (MEC) was defined as the lowest concentration to produce a significant change (p ≤ 0.05) in all the analyzed parameters compared to the control (solvent-treated) cells. For all the compounds and studied parameters, the MEC led to at least a 20% variation in fluorescence intensity or in the corresponding morphological parameter compared to the untreated cultures. The cumulative index (CI) for each compound and parameter was also calculated as the overall score at the concentrations at which the effect was above the MEC, as previously defined (Schoonen et al. 2005).
All the data are expressed as mean ± SEM values and represent triplicate measurements. A Student’s t test was used for statistical evaluations between two groups, whereas ANOVA followed by Tukey’s multiple comparison test was used when comparing 3 or more groups. The level of significance was chosen as p < 0.05 and calculated with GraphPad Prism v.6.1.
For the multivariate analysis, all the statistical analyses and data plots were run with the free software MetaboAnalyst 3.0 (Xia and Wishart 2016). Principal component analysis (PCA) was used to visualize the natural interrelationship among the samples by performing pairwise comparisons (i.e., control vs. toxic compounds). PLS-DA (projection of latent structures-discriminant analysis) was employed to develop classificatory/predictive models based on the altered patterns aimed to discriminate between hepatotoxic and non-hepatotoxic compounds, as well as differentiating the effects on distinct donors of UHH.
Results
Long-term culture of Upcyte hepatocytes over the time
In the present study, we evaluated UHH as a model for the detection of long-term hepatotoxicity after repeated-dose exposure. UHH maintained their morphology after 3 weeks in culture (Supplementary Fig. 2S). Additionally, UHH cultures maintained stable levels of albumin secretion over 3 weeks with similar levels of PHH (data not shown). Prior to assessing the effects of drugs on these cultures, we first established that UHH cultures from different donors remained viable throughout the entire 3-week period (Fig. 2S).

Inter-donor differences in gene expression profile and activities of drug-metabolizing enzymes. a The mRNA levels of 14 genes of phase I and phase II enzymes were analyzed in UHH from 3 donors at day 0, 1, 3, 7, 14 or 21 of culture. b CYP activities were measured in UHH at different times of culture and compared to those of HepG2 (day 2 of culture), HepaRG (differentiated cells after 0, 7 and 14 days) and PHH (day 1 of culture). c Phase II activities (pmol/mg/min) in UHH (day 3 of culture)
Transcriptomic and functional characterization of drug-metabolizing enzymes
The mRNA expression of 14 genes related to drug metabolism was comparatively analyzed in UHH from three different donors. The results in Fig. 2 show some inter-donor differences in expression level of major hepatic cytochrome P450 (CYP) enzymes involved in drug metabolism. UHH from donor 1 showed the highest expression of CYP2A6, CYP2C19 and CYP3A4 genes, whereas donor 2 showed the highest expression of CYP2C9 and CYP2E1 and, the lowest CYP mRNA levels usually corresponded to UHH 3. Regarding the temporal evolution of the expression profile, donor 2 showed a progressive increase in mRNA levels of CYP enzymes up to 14 or 21 days of culture, which was not observed in the other donors. Inter-donor differences were also observed in the expression levels of P450 oxidoreductase and phase II enzymes. The highest expression of UGT1A1 and UGT2B7 genes was found in UHH 2, particularly after prolonged time in culture (up to 21 days). Similarly, the expression of GSTA1, GSTA2 and GSTM1 genes was generally higher in UHH 2 than in cells from the other donors (Fig. 2a).
High inter-donor variability among the UHH preparations was also observed in CYP and phase II activities. Very low levels, if any, of CYP1A2, CYP2C19 and CYP2D6 activities were observed in UHH (Supplementary Fig. 3S); whereas measurable levels of all other CYP activities were found up to 21 day of culture (Fig. 2b). The CYP2C9 and CYP2E1 activities in UHH were lower than those of PHH, whereas the CYP2A6, CYP2B6 and CYP3A4 activities were comparable in both cell types. When compared to HepaRG cells, the UHH showed higher CYP2A6, CYP2B6 and CYP2E1 activities, but lower activity levels of CYP2C9, CYP2D6 and CYP3A4. Overall, the activity levels of CYPs in UHH were markedly higher than in HepG2 cells. Regarding phase II enzymes, UGT1A1 activity levels in UHH were comparable to those in HepaRG and PHH, whereas UGT2B7 and GST activities were lower in UHH (Fig. 2c). As observed for CYPs, phase II activities in UHH were higher than in HepG2 cells.
Repeated-dose toxicity study in UHH
Potential toxic effects of 15 selected drugs were examined in UHH from three donors. Cells were exposed for different periods (up to 21 days) to three different concentrations of each drug (corresponding to 0.2-, 1- and 5-fold Cmax value) following a repeated-dose regimen described in Fig. 1. After treatments, eight parameters indicative of cell toxicity (viability, mitochondrial membrane potential (MMP), intracellular calcium, ROS, mitochondrial superoxide, GSH, phospholipids and lipids) were analyzed using a HCS-based assay and data were subjected to non-supervised multivariate data analysis techniques to evaluate the presence of any pattern allowing us to differentiate between hepatotoxic and non-hepatotoxic compounds. The PCA scores plots showed an almost complete separation between non-hepatotoxic or hepatotoxic compounds (Fig. 3a). When a supervised analysis of the HCS data was performed considering toxic and non-toxic compounds, the PLS-DA scores plots showed a clear separation between groups (Fig. 3b), demonstrating the capability of our strategy to discriminate drugs able to induce cell damage, even at low, in vivo-relevant concentrations (close to therapeutic blood levels).

Multivariate data analysis overview of the changes induced by model hepatotoxic and non-hepatotoxic compounds. a PCA scores plots using two principal components corresponding to data obtained from UHH treated with compounds acting through different mechanisms after repeated-dose exposure. b PLS-DA scores plot corresponding to a model aimed at the discrimination between toxic and non-toxic compounds
Mechanistic study of repeated-dose toxicity in UHH
The multiparametric analysis by HCS allowed to clearly distinguish between toxic and non-toxic compounds and also offered important mechanistic information. MEC values for each endpoint indicative of cell damage after repeated-dose exposure are summarized in Fig. 4. Although significant changes in cell viability were in general observed only for some compounds (i.e., methotrexate) and/or after long-treatment, other effects were early induced by some compounds at non-lethal concentrations. According to the score system created from MEC, the main mechanisms implicated in toxicity for each compound may be outlined and time-exposure effects could be also distinguished. For instance, in all the hepatotoxic compounds lower MEC values were observed as the time-exposure to the drugs increased, showing the effects of a chronic exposure. On the other hand, it allowed differentiating between mechanisms that are implicated in the acute or short-term toxicity (i.e., changes in calcium homeostasis) and those which are significantly detected as the cells are repeatedly exposed to the drugs (i.e., phospholipidosis). The assessment of the toxicity of test compounds to UHH revealed donor- and time-dependent effects in several parameters, although in the overall all drugs responded as toxic as expected in agreement with the described mechanisms of toxicity for each compound (Supplementary Table 1S).

Mechanisms implicated in drug-induced hepatotoxicity over time. UHH cells were exposed to 3 concentrations of each drug (0.2 ×, 1 ×, or 5 × Cmax) and eight parameters indicative of cell toxicity were evaluated by HCS after 1, 3, 7, 14 or 21 days of treatment. Effects obtained for each parameter and time (mean of 3 UHH donors) are indicated as the MEC (minimal effective concentration) of each compound that produces a significant change over untreated cells: red (0.2 × Cmax), orange (Cmax), yellow (5 × Cmax) or green (no effect). The outlined square highlights those endpoints that were described in the literature as the mechanism implicated in DILI. VIA viability, MMP mitochondrial membrane potential, CA intracellular calcium concentration, ROS ROS production, MSX mitochondrial superoxide generation, PLP phospholipidosis, LIP lipid overaccumulation
Interdonor differences in DILI
Next, in order to highlight the potential interdonor differences in the response to test compounds, we analyzed the data differentiating between donors. The PCA score plot revealed a separation between toxic and non-toxic compounds independently of the donor (Fig. 5a). When only the hepatotoxic compounds were analyzed, the PLS-DA score plot showed a clear separation among UHH donors, suggesting differences in their susceptibility to drugs (Fig. 5b).

UHH inter-donor variable response to hepatotoxic compounds. a Multivariate data analysis overview of the changes induced by model hepatotoxic and non-hepatotoxic compounds in the three donors used in this study. b PCA scores plots using two principal components corresponding to data obtained from three different donors treated with hepatotoxic drugs after repeated-dose exposure. c Cumulative index (CI) for each of the analyzed parameters indicative of hepatotoxicity in the different donors. aAt least p < 0.05 (donor 2 compared to donor 1); bat least p < 0.05 (donor 3 compared to donor 1); bat least p < 0.05 (donor 3 compared to donor 2) (Student’s t test). The statistical significance is indicated in supplementary Table S4
Then, we determined the CI for each of the different studied mechanisms of toxicity and distinct donors (Fig. 5c). For instance, donor 3 showed the highest effects on lipid and phospholipid accumulation as well as in cell viability, whereas other parameters remained in the same values (i.e., mitochondrial superoxide, MMP) or even lower (i.e., calcium accumulation) than the mean for other donors, indicating a special sensitivity of donor 3 for chronic effects such as steatosis or phospholipidosis. By contrast, donor 2 seemed to be the least sensitive one when exposed to hepatotoxic compounds, showing the lowest CI for most of the parameters analyzed. Finally, donor 1 seemed to show the highest CI for parameters altered after short exposure such as intracellular calcium concentration, indicating a particular susceptibility of this donor regarding calcium homeostasis, but showed the lower response to drug-induced steatosis or phospholipidosis.
Differential effects depending on the donor and exposure time after repeated exposure to valproate, flutamide and chlorpromazine
The evaluation of the toxicity of valproate by means of HCS demonstrated that lipid overaccumulation, intracellular calcium concentration, ROS production and MMP were the most sensitive parameters to be altered after exposure to it (Fig. 6), although GSH and mitochondrial superoxide were also significantly reduced at longer periods of incubation (Supplementary Fig. 4S). These effects were observed even at low concentrations and short exposures, in absence of reductions in cell viability. Differential effects among donors were detected, with donor 3 being the less susceptible to valproate effects. Donor 2 was particularly sensitive to alterations in calcium homeostasis (at shorter exposure periods or lower concentrations) and showed the highest increases in ROS levels, whereas the biggest effects on lipid accumulation and MMP were found in donor 1, confirming the inter-donor variability.

Mechanisms implicated in valproate-induced hepatotoxicity in UHH. a Differential representative effects of valproate at 5 × Cmax concentration on lipid overaccumulation, ROS production, intracellular calcium concentration and MMP in three distinct donors. Statistical significance (ANOVA followed by Tukey’s test) between groups is indicated as follows: a(donor 2 vs. donor 1), b(donor 3 vs. donor 1) and c(donor 2 vs. donor 3). b Mean values of the effects of valproate at the different concentrations used after 14-day repeated-dose incubation. The dotted line indicates the minimum effective concentration value. c Mechanisms implicated in valproate-induced hepatotoxicity. The detected mechanisms by HCS are indicated in grey. CPT carnitine palmitoyltransferase, MPT mitochondrial permeability transition
Similarly, interdonor differences in flutamide toxicity were observed. Donor 3 was relatively insensitive to flutamide, whereas effects in several parameters were observed in other donors. In particular, the highest changes in intracellular calcium, MMP and mitochondrial superoxide level were found in donor 1, whereas the highest GSH depletion corresponded to donor 2 (Fig. 7a). Marked differences depending on the donor were also detected in other parameters (Supplementary Fig. 5S). In all cases, significant effects were detected after 14 or 21 days of exposure to the drug revealing the importance of the period on incubation to detect hepatotoxicity at therapeutic concentrations. As an average of all donors, intracellular calcium and ROS were the parameters most affected by flutamide treatment (Fig. 7b).

Mechanisms implicated in flutamide-induced hepatotoxicity in UHH. a Differential representative effects of flutamide at 5 × Cmax concentration on intracellular calcium concentration, MMP, mitochondrial superoxide production, and GSH depletion in three distinct donors. Statistical significance (ANOVA followed by Tukey’s test) between groups is indicated as follows: a(donor 2 vs. donor 1), b(donor 3 vs. donor 1) and c(donor 2 vs. donor 3). b Mean values of the effects of flutamide at the different concentrations used after 14-day repeated-dose incubation. The dotted line indicates the minimum effective concentration value. c Mechanisms implicated in flutamide-induced hepatotoxicity. The detected mechanisms by HCS are indicated in grey. MCR mitochondrial respiratory chain
When the toxicity of chlorpromazine was analyzed, a significant effect in phospholipid accumulation, mainly after long-term incubation (14–21 days), was observed in all donors, although the effects in donor 1 were significantly lower than in the other two donors (Fig. 8). Some interdonor differences were also observed in other parameters such as intracellular calcium or ROS levels (Supplementary Fig. 6S). The study of chlorpromazine-induced toxicity in UUH revealed induction of oxidative stress, mitochondrial injury and phospholipidosis, which are in concordance with the described toxicity mechanisms for this drug.

Mechanisms implicated in chlorpromazine-induced hepatotoxicity in UHH. a Differential representative effects of chlorpromazine at 5 × Cmax concentration on phospholipidosis, mitochondrial superoxide and ROS production and GSH depletion in three distinct donors. Statistical significance (ANOVA followed by Tukey’s test) between groups is indicated as follows: a(donor 2 vs. donor 1), b(donor 3 vs. donor 1) and c(donor 2 vs. donor 3). b Mean values of the effects of chlorpromazine at the different concentrations used after 14-day repeated-dose incubation. The dotted line indicates the minimum effective concentration value. c Mechanisms implicated in chlorpromazine-induced hepatotoxicity. The detected mechanisms by HCS are indicated in grey
Discussion
DILI may occur after several weeks or months of treatment and usually requires metabolism of the drug (Antherieu et al. 2011). Despite this, many in vitro studies are performed between 24 and 48 h, therefore exposing the cells to a range of acute doses not comparable to therapeutic concentrations (Germano et al. 2014). Repeated-dose administration is a more relevant scenario for therapeutics, being usually evaluated in in vivo studies which yield information on general characteristics of the toxicity, delayed responses and/or cumulative effects (Truisi et al. 2015). To mimic long-term repeated exposures in vitro, cell models which retain hepatic phenotype for a sufficiently long time should be used. In the present study, we explored the feasibility of the combined use of UHH and a HCS-based multiparametric testing strategy to evaluate drug toxicity and to identify the mechanisms underlying these toxic effects after repeated-dose exposure. The study revealed that the repeated long-term exposure of UHH from three different donors at clinically relevant concentrations of model hepatotoxic drugs resulted in toxic effects and allowed identifying the main mechanisms implicated and detecting differences among donors which reflects the inter-individual variability which exists in the clinic.
In our study, a range of concentrations around Cmax and a repeated exposure of the cells were used with the idea of mimicking drug administration in humans, and, thus, improving the prediction of DILI in preclinical development. Other models such as HepaRG (Antherieu et al. 2011; Josse et al. 2008), differentiated induced pluripotent stem cells (Holmgren et al. 2014), hepatocytes cultured in sandwich configuration (Germano et al. 2014) or 3D cultures of PHH (Bell et al. 2017; Vorrink et al. 2018) have been used for repeated-dose exposures for up to 14 days. The UHH have revealed important advantages over existing models because allowed longer treatments (up to 21 days) without loss of hepatic phenotype and the possibility to choose between UHH originated from distinct individuals to assess the differential response depending on the donor.
The possibility of testing drug effects on different donors represents a major advantage of UHH over the widely used HepG2 or HepaRG cell lines. When different UHH were used for long-term repeated exposures to hepatotoxic compounds, distinct responses were observed, indicating that inter-individual susceptibility exists towards hepatotoxicity. It is well-known that for many drugs hepatotoxic events affect only to a reduced group of susceptible patients and several individual factors may predispose to such idiosyncratic reactions (e.g., genetics, underlying diseases, metabolic profile, etc.) (Gomez-Lechon et al. 2016). Analysis of gene expression and activities of drug-metabolizing enzymes in UHH show the interdonor variability that has been previously described for PHH cultures as a direct reflection of in vivo variability (Kostadinova et al. 2013; Sioud and Melien 2007). Such differences in metabolic capability may result in variable responses to drug-induced hepatotoxicity, particularly for bioactivable drugs or those with high metabolic rates. In addition to variations in drug metabolism, interdonor differences in other genes of relevance to toxicity should be considered. For example, we previously found differences in the expression of genes related to lipid metabolism among UHH cultures (Tolosa et al. 2016b), which may explain the interdonor differences observed in drug-induced accumulation of neutral lipid or phospholipids (Fig. 5c).
The toxicity of tested drugs was estimated as not only reductions in cell viability, but also as alterations in several parameters that are indicative of cell damage via different mechanisms (mitochondrial dysfunction, oxidative stress, impairment of calcium homeostasis, lipid accumulation or phospholipidosis). The importance of repeated long-term exposure, which is more representative of the clinical setting, was evident in the present study since there are effects only detected after chronic exposure or the effects are clearly increased after prolonged periods of incubation, suggesting that DILI may require time to develop. In fact, time-dependent accumulation of intracytoplasmic vesicles corresponding to steatosis and phospholipidosis induced by amiodarone has been described in other models (Pomponio et al. 2015). Recently, Bell et al. (2017) compared the capacity of several cell hepatic models to predict drug hepatotoxicity and proposed long-term treated 3D PHH spheroids as the most sensitive system. Interestingly, our results are comparable to those reported in PHH spheroids after repeated exposure for 14 days to low concentrations of model drugs close to therapeutic Cmax values (Bell et al. 2017; Vorrink et al. 2018) (Supplementary table 5S), evidencing the high sensitivity of our proposal for drug hepatotoxicity screenings. These findings support the idea that when developing in vitro strategies for the prediction of hepatotoxicity, not only the cell culture system and dosing protocol should be carefully selected (e.g., PHH vs. cell lines, 2D vs. 3D cultures, acute vs. long-term exposure…), but also the type of assay used to detect toxicity. Once again, the high sensitivity of multiparametric assays that detect subtle changes indicative of toxicity at low concentrations of the drug that do not induce important alterations in cell viability is evidenced (Germano et al. 2014; Tolosa et al. 2012).
The proposed strategy allows distinguishing between DILI and non-DILI compounds under chronic exposure as well as identifying the underlying mechanisms of hepatotoxicity. The anticonvulsant agent valproate is known to induce steatosis as well as rare but potentially fatal idiosyncratic hepatotoxicity (Saruwatari et al. 2010). Although the mechanisms of liver injury by valproate are still not well understood, the role of mitochondrial dysfunction, including interference with β-oxidation and oxidative stress, derived from enhanced ROS formation and/or altered GSH homeostasis, have been suggested (Silva et al. 2008). Accordingly, increased lipid accumulation, ROS generation, GSH depletion and mitochondrial toxicity were reproduced in UHH. Interestingly, the lower sensitivity of donor 3 to valproate-induced toxicity, compared to donors 1 and 2, may be related to its lower level of CYP2C9 activity (Fig. 2a), the major enzyme involved in the formation of 4-ene-valproic acid (Kiang et al. 2006). Although considered a minor route of valproate metabolism, formation of 4-ene-valproic acid and its subsequent oxidation in the mitochondria to form 2,4-diene metabolite are key bioactivation events that lead to the depletion of GSH and CoA pools and, in turn, inhibition of β-oxidation (Ghodke-Puranik et al. 2013). Moreover, high UGT2B7 activity shown in donor 3 (Fig. 2c) can contribute to valproate detoxification (Saruwatari et al. 2010).
Similarly, toxic effects found in the UHH exposed to flutamide completely coincided with those reported in the literature (Ball et al. 2016; Kashimshetty et al. 2009). This antiandrogen drug used for the treatment of prostate cancer is a known mitochondrial toxicant that received a black box warning because of the risk of rare episodes of idiosyncratic hepatotoxicity (Walgren et al. 2005). The mechanisms of liver damage in susceptible patients remain outstanding but mitochondrial dysfunction and oxidative stress are likely to be involved (Kashimshetty et al. 2009). Flutamide hepatotoxicity has been related to bioactivation by CYPs with formation of several reactive metabolites, including the mitotoxic metabolite 2-hydroxyflutamide that has been related to idiosyncratic hepatotoxicity in flutamide-treated patients (Ball et al. 2016; Kang et al. 2008). We recently reported that flutamide induced differential toxicity depending on the specific CYP overexpressed and its level in a model of transduced HepG2 cells (Tolosa et al. 2018). Then, interdonor differences in sensitivity observed in UHH may be explained by individual variations in CYP activities. In fact, the high susceptibility of donor 1 to flutamide-induced mitochondrial effects may be due, at least in part, to its relative high levels of CYP3A4 and CYP2C19 activities, both involved in flutamide bioactivation.
Chlorpromazine is an antipsychotic neuroleptic agent that causes rare but severe episodes of liver injury through multiple mechanisms, including phospholipidosis or cholestasis (Selim and Kaplowitz 1999). In addition to parent compound toxicity, chlorpromazine is bioactivated by oxidation to electrophilic metabolites that deplete GSH pool and induce mitochondrial dysfunction (MacAllister et al. 2013; Mennecozzi et al. 2015). Accordingly to its described phospholipid binding capacity, marked phospholipid accumulation was observed in two of the three UHH preparations treated with therapeutic concentrations of the drug for 2–3 weeks. An in vitro biokinetics study revealed that chlorpromazine accumulates in hepatic cells after repeated-dose exposure and that maintenance of high intracellular drug concentrations depend on metabolic rates (Broeders et al. 2015). Then, the relative insensitivity of donor 1 to chlorpromazine-induced phospholipidosis may be explained by its high CYP activity levels, as the drug undergoes extensive metabolism by hepatic CYPs, including 7-hydroxylation, 5-sulfoxidation and N-demethylation (Wojcikowski et al. 2010).
In conclusion, the UHH faithfully maintain the expression and activity levels of the drug-metabolizing enzymes over time and turned out to be a sensitive system to detect drug hepatotoxicity after a repeated-dose strategy at clinically relevant concentrations. Moreover, we showed that these improved results are not only due to the chronic toxicity, but also to the detection of the underlying mechanisms of drug-induced toxicity detected by HCS before cytotoxicity occurs. Combined, the data presented here suggested that the UHH are a suitable model for the investigation of long-term hepatotoxicity which includes the possibility to choose between different donors with diverse genotypes which would reflect the variability among the population and could be used to improve the prediction in preclinical development.
Abbreviations
- DILI:
-
Drug-induced liver injury
- PHH:
-
Primary human hepatocytes
- UHH:
-
Upcytes human hepatocytes
- HCS:
-
High-content screening
- C max :
-
Therapeutic peak plasmatic concentration
- MEC:
-
Minimum effective concentration
- CI:
-
Cumulative index
- PCA:
-
Principal component analysis
- PLS-DA:
-
Projection of latent structures-discriminant analysis
- MMP:
-
Mitochondrial membrane potential
- CYP:
-
Cytochrome P450
References
Antherieu S, Rogue A, Fromenty B, Guillouzo A, Robin MA (2011) Induction of vesicular steatosis by amiodarone and tetracycline is associated with up-regulation of lipogenic genes in HepaRG cells. Hepatology 53(6):1895–1905. https://doi.org/10.1002/hep.24290
Ball AL, Kamalian L, Alfirevic A, Lyon JJ, Chadwick AE (2016) Identification of the additional mitochondrial liabilities of 2-hydroxyflutamide when compared with its parent compound, flutamide in HepG2 cells. Toxicol Sci 153(2):341–351. https://doi.org/10.1093/toxsci/kfw126
Bell CC, Lauschke VM, Vorrink SU et al (2017) Transcriptional, functional, and mechanistic comparisons of stem cell-derived hepatocytes, HepaRG cells, and three-dimensional human hepatocyte spheroids as predictive in vitro systems for drug-induced liver injury. Drug Metab Dispos 45(4):419–429. https://doi.org/10.1124/dmd.116.074369
Bjornsson ES (2015) Drug-induced liver injury: an overview over the most critical compounds. Arch Toxicol 89(3):327–334. https://doi.org/10.1007/s00204-015-1456-2
Broeders JJ, Parmentier C, Truisi GL et al (2015) Biokinetics of chlorpromazine in primary rat and human hepatocytes and human HepaRG cells after repeated exposure. Toxicol In Vitro 30(1 Pt A):52–61. https://doi.org/10.1016/j.tiv.2014.08.012
Burkard A, Dahn C, Heinz S et al (2012) Generation of proliferating human hepatocytes using Upcyte(R) technology: characterisation and applications in induction and cytotoxicity assays. Xenobiotica 42(10):939–956. https://doi.org/10.3109/00498254.2012.675093
Donato MT, Tolosa L, Jimenez N, Castell JV, Gomez-Lechon MJ (2012) High-content imaging technology for the evaluation of drug-induced steatosis using a multiparametric cell-based assay. J Biomol Screen 17(3):394–400. https://doi.org/10.1177/1087057111427586
Funk C, Roth A (2017) Current limitations and future opportunities for prediction of DILI from in vitro. Arch Toxicol 91(1):131–142. https://doi.org/10.1007/s00204-016-1874-9
Germano D, Uteng M, Pognan F, Chibout SD, Wolf A (2014) Determination of liver specific toxicities in rat hepatocytes by high content imaging during 2-week multiple treatment. Toxicol In Vitro. https://doi.org/10.1016/j.tiv.2014.05.009
Ghodke-Puranik Y, Thorn CF, Lamba JK et al (2013) Valproic acid pathway: pharmacokinetics and pharmacodynamics. Pharmacogenet Genomics 23(4):236–241. https://doi.org/10.1097/FPC.0b013e32835ea0b2
Gomez-Lechon MJ, Tolosa L (2016) Human hepatocytes derived from pluripotent stem cells: a promising cell model for drug hepatotoxicity screening. Arch Toxicol 90(9):2049–2061. https://doi.org/10.1007/s00204-016-1756-1
Gomez-Lechon MJ, Tolosa L, Conde I, Donato MT (2014) Competency of different cell models to predict human hepatotoxic drugs. Expert Opin Drug Metab Toxicol 10(11):1553–1568. https://doi.org/10.1517/17425255.2014.967680
Gomez-Lechon MJ, Tolosa L, Donato MT (2016) Metabolic activation and drug-induced liver injury: in vitro approaches for the safety risk assessment of new drugs. J Appl Toxicol 36(6):752–768. https://doi.org/10.1002/jat.3277
Holmgren G, Sjogren AK, Barragan I et al (2014) Long-term chronic toxicity testing using human pluripotent stem cell-derived hepatocytes. Drug Metab Dispos 42(9):1401–1406. https://doi.org/10.1124/dmd.114.059154
Josse R, Aninat C, Glaise D et al (2008) Long-term functional stability of human HepaRG hepatocytes and use for chronic toxicity and genotoxicity studies. Drug Metab Dispos 36(6):1111–1118. https://doi.org/10.1124/dmd.107.019901
Kang P, Dalvie D, Smith E, Zhou S, Deese A, Nieman JA (2008) Bioactivation of flutamide metabolites by human liver microsomes. Drug Metab Dispos 36(7):1425–1437. https://doi.org/10.1124/dmd.108.020370
Kashimshetty R, Desai VG, Kale VM et al (2009) Underlying mitochondrial dysfunction triggers flutamide-induced oxidative liver injury in a mouse model of idiosyncratic drug toxicity. Toxicol Appl Pharmacol 238(2):150–159. https://doi.org/10.1016/j.taap.2009.05.007
Kiang TK, Ho PC, Anari MR, Tong V, Abbott FS, Chang TK (2006) Contribution of CYP2C9, CYP2A6, and CYP2B6 to valproic acid metabolism in hepatic microsomes from individuals with the CYP2C9*1/*1 genotype. Toxicol Sci 94(2):261–271. https://doi.org/10.1093/toxsci/kfl096
Kostadinova R, Boess F, Applegate D et al (2013) A long-term three dimensional liver co-culture system for improved prediction of clinically relevant drug-induced hepatotoxicity. Toxicol Appl Pharmacol 268(1):1–16. https://doi.org/10.1016/j.taap.2013.01.012
Kramer NI, Di Consiglio E, Blaauboer BJ, Testai E (2015) Biokinetics in repeated-dosing in vitro drug toxicity studies. Toxicol In Vitro 30(1 Pt A):217–224. https://doi.org/10.1016/j.tiv.2015.09.005
Levy G, Bomze D, Heinz S et al (2015) Long-term culture and expansion of primary human hepatocytes. Nat Biotechnol 33(12):1264–1271. https://doi.org/10.1038/nbt.3377
MacAllister SL, Young C, Guzdek A, Zhidkov N, O’Brien PJ (2013) Molecular cytotoxic mechanisms of chlorpromazine in isolated rat hepatocytes. Can J Physiol Pharmacol 91(1):56–63. https://doi.org/10.1139/cjpp-2012-0223
Mennecozzi M, Landesmann B, Palosaari T, Harris G, Whelan M (2015) Sex differences in liver toxicity-do female and male human primary hepatocytes react differently to toxicants in vitro? PLoS One 10(4):e0122786. https://doi.org/10.1371/journal.pone.0122786
Mueller D, Kramer L, Hoffmann E, Klein S, Noor F (2014) 3D organotypic HepaRG cultures as in vitro model for acute and repeated dose toxicity studies. Toxicol In Vitro 28(1):104–112. https://doi.org/10.1016/j.tiv.2013.06.024
Norenberg A, Heinz S, Scheller K, Hewitt NJ, Braspenning J, Ott M (2013) Optimization of upcyte(R) human hepatocytes for the in vitro micronucleus assay. Mutat Res 758(1–2):69–79. https://doi.org/10.1016/j.mrgentox.2013.09.008
Pomponio G, Savary CC, Parmentier C et al (2015) In vitro kinetics of amiodarone and its major metabolite in two human liver cell models after acute and repeated treatments. Toxicol In Vitro 30(1 Pt A):36–51. https://doi.org/10.1016/j.tiv.2014.12.012
Prieto P, Baird AW, Blaauboer BJ et al (2006) The assessment of repeated dose toxicity in vitro: a proposed approach. The report and recommendations of ECVAM workshop 56. Altern Lab Anim 34(3):315–341
Reuben A, Koch DG, Lee WM, Acute Liver Failure Study G (2010) Drug-induced acute liver failure: results of a U.S. multicenter, prospective study. Hepatology 52(6):2065–2076. https://doi.org/10.1002/hep.23937
Saruwatari J, Ishitsu T, Nakagawa K (2010) Update on the genetic polymorphisms of drug-metabolizing enzymes in antiepileptic drug therapy. Pharmaceuticals (Basel) 3(8):2709–2732. https://doi.org/10.3390/ph3082709
Schoonen WG, Westerink WM, de Roos JA, Debiton E (2005) Cytotoxic effects of 100 reference compounds on Hep G2 and HeLa cells and of 60 compounds on ECC-1 and CHO cells. I mechanistic assays on ROS, glutathione depletion and calcein uptake. Toxicol In Vitro 19(4):505–516. https://doi.org/10.1016/j.tiv.2005.01.003
Selim K, Kaplowitz N (1999) Hepatotoxicity of psychotropic drugs. Hepatology 29(5):1347–1351. https://doi.org/10.1002/hep.510290535
Silva MF, Aires CC, Luis PB et al (2008) Valproic acid metabolism and its effects on mitochondrial fatty acid oxidation: a review. J Inherit Metab Dis 31(2):205–216. https://doi.org/10.1007/s10545-008-0841-x
Sioud M, Melien O (2007) Treatment options and individualized medicine. Methods Mol Biol 361:327–340. https://doi.org/10.1385/1-59745-208-4:327
Sison-Young RL, Mitsa D, Jenkins RE et al (2015) Comparative proteomic characterization of 4 human liver-derived single cell culture models reveals significant variation in the capacity for drug disposition, bioactivation, and detoxication. Toxicol Sci 147(2):412–424. https://doi.org/10.1093/toxsci/kfv136
Tolosa L, Pinto S, Donato MT et al (2012) Development of a multiparametric cell-based protocol to screen and classify the hepatotoxicity potential of drugs. Toxicol Sci 127(1):187–198. https://doi.org/10.1093/toxsci/kfs083
Tolosa L, Carmona A, Castell JV, Gomez-Lechon MJ, Donato MT (2015) High-content screening of drug-induced mitochondrial impairment in hepatic cells: effects of statins. Arch Toxicol 89(10):1847–1860. https://doi.org/10.1007/s00204-014-1334-3
Tolosa L, Gomez-Lechon MJ, Jimenez N, Hervas D, Jover R, Donato MT (2016a) Advantageous use of HepaRG cells for the screening and mechanistic study of drug-induced steatosis. Toxicol Appl Pharmacol 302:1–9. https://doi.org/10.1016/j.taap.2016.04.007
Tolosa L, Gomez-Lechon MJ, Lopez S et al (2016b) Human upcyte hepatocytes: characterization of the hepatic phenotype and evaluation for acute and long-term hepatotoxicity routine testing. Toxicol Sci 152(1):214–229. https://doi.org/10.1093/toxsci/kfw078
Tolosa L, Jimenez N, Perez G, Castell JV, Gomez-Lechon MJ, Donato MT (2018) Customised in vitro model to detect human metabolism-dependent idiosyncratic drug-induced liver injury. Arch Toxicol 92(1):383–399. https://doi.org/10.1007/s00204-017-2036-4
Truisi GL, Consiglio ED, Parmentier C et al (2015) Understanding the biokinetics of ibuprofen after single and repeated treatments in rat and human in vitro liver cell systems. Toxicol Lett 233(2):172–186. https://doi.org/10.1016/j.toxlet.2015.01.006
Vanhaecke T, Pauwels M, Vinken M, Ceelen L, Rogiers V (2011) Towards an integrated in vitro strategy for repeated dose toxicity testing. Arch Toxicol 85(5):365–366. https://doi.org/10.1007/s00204-011-0711-4
Vorrink SU, Zhou Y, Ingelman-Sundberg M, Lauschke VM (2018) Prediction of drug-induced hepatotoxicity using long-term stable primary hepatic 3D spheroid cultures in chemically defined conditions. Toxicol Sci 163(2):655–665. https://doi.org/10.1093/toxsci/kfy058
Walgren JL, Mitchell MD, Thompson DC (2005) Role of metabolism in drug-induced idiosyncratic hepatotoxicity. Crit Rev Toxicol 35(4):325–361
Wojcikowski J, Boksa J, Daniel WA (2010) Main contribution of the cytochrome P450 isoenzyme 1A2 (CYP1A2) to N-demethylation and 5-sulfoxidation of the phenothiazine neuroleptic chlorpromazine in human liver—a comparison with other phenothiazines. Biochem Pharmacol 80(8):1252–1259. https://doi.org/10.1016/j.bcp.2010.06.045
Xia J, Wishart DS (2016) Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr Protoc Bioinform 55:14-10. https://doi.org/10.1002/cpbi.11
Acknowledgements
This work has been supported by the Institute of Health Carlos III (Plan Estatal de I+D+i 2013–2016) and cofinanced by the European Regional Development Fund “A way to achieve Europe” (FEDER) through Grants PI16/00333 and CP16/00097. L.T. was supported by ISCIII CP16/00097. The authors acknowledge financial support from the ALIVE Foundation. The scientific support from the Cytomics Unit from the Instituto de Investigación Sanitaria La Fe and the technical advice of Astrid Nörenberg and Timo Johannsen are acknowledged.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Tolosa, L., Jiménez, N., Pelechá, M. et al. Long-term and mechanistic evaluation of drug-induced liver injury in Upcyte human hepatocytes. Arch Toxicol 93, 519–532 (2019). https://doi.org/10.1007/s00204-018-2349-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00204-018-2349-y