
Abstract
Aims/hypothesis
Nicotinamide nucleotide transhydrogenase (NNT) is involved in mitochondrial NADPH production and its spontaneous inactivating mutation (NntTr [Tr, truncated]) is usually considered to be the main cause of the lower glucose tolerance of C57BL/6J vs C57BL/6N mice. However, the impact of this mutation on glucose tolerance remains disputed. Here, we singled out the impact of NntTr from that of other genetic variants between C57BL/6J and C57BL/6N mice on mitochondrial glutathione redox state (EGSH), glucose-stimulated insulin secretion (GSIS) and glucose tolerance.
Methods
Male and female N5BL/6J mice that express wild-type Nnt (NntWT) or NntTr (N5-WT and N5-Tr mice) on the C57BL/6J genetic background were obtained by crossing N5BL/6J NntWT/Tr heterozygous mice. C57BL/6J and C57BL/6N mice were from Janvier Labs. The Nnt genotype was confirmed by PCR and the genetic background by whole genome sequencing of one mouse of each type. Glucose tolerance was assessed by IPGTT, ITT and fasting/refeeding tests. Stimulus–secretion coupling events and GSIS were measured in isolated pancreatic islets. Cytosolic and mitochondrial EGSH were measured using the fluorescent redox probe GRX1–roGFP2 (glutaredoxin 1 fused to redox-sensitive enhanced GFP).
Results
The Nnt genotype and genetic background of each type of mouse were confirmed. As reported previously in C57BL/6N vs C57BL/6J islets, the glucose regulation of mitochondrial (but not cytosolic) EGSH and of NAD(P)H autofluorescence was markedly improved in N5-WT vs N5-Tr islets, confirming the role of NNT in mitochondrial redox regulation. However, ex vivo GSIS was only 1.2–1.4-times higher in N5-WT vs N5-Tr islets, while it was 2.4-times larger in C57BL/6N vs N5-WT islets, questioning the role of NNT in GSIS. In vivo, the ITT results did not differ between N5-WT and N5-Tr or C57BL/6N mice. However, the glucose excursion during an IPGTT was only 15–20% lower in female N5-WT mice than in N5-Tr and C57BL/6J mice and remained 3.5-times larger than in female C57BL/6N mice. Similar observations were made during a fasting/refeeding test. A slightly larger (~30%) impact of NNT on glucose tolerance was found in males.
Conclusions/interpretation
Although our results confirm the importance of NNT in the regulation of mitochondrial redox state by glucose, they markedly downsize the role of NNT in the alteration of GSIS and glucose tolerance in C57BL/6J vs C57BL/6N mice. Therefore, documenting an NntWT genotype in C57BL/6 mice does not provide proof that their glucose tolerance is as good as in C57BL/6N mice.
Graphical abstract

Similar content being viewed by others



Introduction
The stimulation of insulin secretion by glucose (glucose-stimulated insulin secretion, GSIS) and other nutrients in pancreatic beta cells is critical to blood glucose homeostasis [1], and its alteration is key to the development of type 2 diabetes [2,3,4]. Following glucose uptake, glycolysis and mitochondrial metabolism increase the beta cell ATP/ADP ratio, thereby closing ATP-sensitive K+ channels. Consequent plasma membrane depolarisation leads to opening of voltage-gated Ca2+ channels and Ca2+ influx that triggers insulin granule exocytosis [1]. Nutrients also amplify Ca2+-stimulated exocytosis through the action of metabolic coupling factors [5, 6], among which cytosolic and mitochondrial NADPH may play an important role [7,8,9,10].
So far, despite increasing availability of human islets and beta cell lines [11, 12], studies about the mechanism of GSIS and its alterations in type 2 diabetes still largely rely on genetically modified mice. In this context, the frequent use of C57BL/6J (BL/6J) mice and their intercrossing with other strains may strongly impact data interpretation and validity [13]. Indeed, compared with C57BL/6N (BL/6N) mice and other strains, BL/6J mice have several genetic variants that affect glucose homeostasis [14,15,16,17], including an inactivating in-frame 5-exon deletion in the Nnt gene encoding nicotinamide nucleotide transhydrogenase (NNT). This inner mitochondrial membrane protein has a dual forward–reverse mode of operation [18,19,20]. When the NADH/NAD+ ratio is high and mitochondria are hyperpolarised, NNT catalyses the transfer of a hydride from NADH to NADP+, thereby contributing to mitochondrial NADPH production. By contrast, when the NADH/NAD+ ratio is low and mitochondria are depolarised, for example in the presence of a mitochondrial uncoupler, the enzyme switches to the reverse mode and oxidises mitochondrial NADPH to produce NADH. NNT is therefore essential in the regulation of mitochondrial NADPH production and glutathione redox state (EGSH), and hence in cell antioxidant defences [18, 19, 21,22,23,24].
Interestingly, compared with BL/6N mice, BL/6J mice present a mild deficit in corticosterone that worsens with age [25], and an impaired glucose tolerance that results from low GSIS without insulin resistance [15, 17, 26, 27]. Their deficit in corticosterone clearly results from the lack of NNT activity [25] and is mirrored by familial glucocorticoid deficiency in humans with NNT mutations [23, 28]. By contrast, the extent to which their impaired glucose tolerance results from the lack of NNT activity is questionable. Initial linkage analysis studies in BL/6J mice intercrossed with C3H/HeH mice revealed several genetic loci linked to glucose intolerance [14, 15], among which the inactivating mutation of Nnt contributed, with seven other loci, to ~35% of the variation of blood glucose during an IPGTT [15]. However, the normalisation of glucose tolerance and plasma insulin levels by baculovirus-mediated transgenic expression of Nnt in BL/6J mice led to the widely-accepted conclusion that their Nnt mutation is at the root of their glucose intolerance [26]. Since then, the differences in glucose homeostasis between BL/6J and BL/6N mice fed a normal or high-fat diet have been the subject of several sometimes controversial studies [27, 29,30,31,32,33], but the predominant role of Nnt truncation in the alteration of glucose tolerance in BL/6J mice has been only rarely questioned [29].
We previously compared BL/6J and BL/6N mice to evaluate the role of NNT in the glucose-mediated regulation of islet NADPH, mitochondrial and cytosolic EGSH and GSIS [33]. We showed that NNT is necessary for the regulation of NADPH and mitochondrial EGSH by glucose in islets, with little impact on cytosolic EGSH. We, however, unexpectedly discovered that, at low glucose, islet NNT operates in the reverse mode and decreases NADPH while increasing mitochondrial EGSH, as it does in the presence of a mitochondrial uncoupler. We also confirmed that BL/6J islets present a defect of insulin secretion in response to glucose compared with BL/6N islets. However, in contrast with other studies [15, 34], this defect occurred downstream of a normal Ca2+ rise. These results are compatible with the idea that, in response to glucose, NNT produces NADPH which, through a regulation of EGSH, amplifies exocytosis [8, 9, 35]. However, although adenovirus-mediated NNT overexpression in mildly-trypsinised BL/6J islets fully restored the glucose regulation of islet NADPH/NADP+ ratio and mitochondrial EGSH, it failed to significantly improve insulin secretion [33], raising doubts about a major role of NNT in GSIS.
To settle the debate on the role of NNT in redox regulation, insulin secretion and glucose tolerance, we have now compared islets from mice that express the wild-type (WT) or truncated (Tr) forms of Nnt (NntWT or NntTr) on the BL/6J background verified by whole genome sequencing (WGS). This allowed us to isolate the effects of the Nnt genotype on islet NADPH, mitochondrial EGSH, GSIS and glucose tolerance while avoiding the confounding effect of other genetic variants between BL/6N and BL/6J mice [21].
Methods
Animals
Animal experiments were authorised by the Université catholique de Louvain Health sciences sector institutional committee on animal experimentation (project 2017/MD/UCL/014). C57BL/6NRj (BL/6N) mice bearing WT Nnt (NntWT) and C57BL/6JRj (BL/6J) mice bearing truncated Nnt (NntTr) were from Janvier Labs (France). N5BL/6J mice (BL/6J mice with NntWT/Tr or NntTr/Tr alleles) were recovered, with kind authorisation of T. Huang (Department of Neurology, Stanford University School of Medicine, Stanford, CA, USA), by the Jackson Laboratory (USA) using cryopreserved sperm B6-gNnt (stock number 906309) and C57BL/6J female mice (https://www.jax.org/strain/000664). These N5BL/6J mice had been generated by repetitive crossing of C57BL/6NHsd mice with C57BL/6J mice over five generations while selecting for NntWT expression and C57BL/6J background by the speed congenic method [21]. In this study, mice homozygous for NntWT or NntTr (N5-WT or N5-Tr mice) were obtained by crossing heterozygous N5BL/6J NntWT/Tr mice. Their Nnt genotype was assessed by tail-DNA PCR, and the genome of one mouse of each type was sequenced (electronic supplementary material [ESM] Methods). Mice were housed in the same room on a 12 h light/dark cycle with ad libitum access to standard chow and water. We used age-matched (3–12 months old) female mice, unless specified otherwise. Whenever possible, N5-WT and N5-Tr mice from the same litter were compared. Besides these selection criteria, mice were not randomised in a controlled manner. Experimenters were not blinded, except for the technician performing insulin RIA.
Islet isolation
Islets were isolated by density-gradient centrifugation (Histopaque 1077, Sigma, USA) following pancreas digestion with collagenase. They were cultured for 1–3 days at 37°C and 5% CO2 in RPMI 1640 medium (Invitrogen, Thermo Fisher Scientific, USA) supplemented with 2 mmol/l glutamine, 100 U/ml penicillin, 100 mg/ml streptomycin, and 5 g/l BSA fraction V (Roche Diagnostics, Germany).
Reagents and Solutions
Experiments were conducted in a Krebs solution (KRB) containing (in mmol/l) NaCl (120), KCl (4.8), CaCl2 (2.5), MgCl2 (1.2), NaHCO3 (20), 1 g/l BSA and d-glucose (0–30, noted G0–G30 in figures). When KCl was increased to 30 mmol/l, NaCl was reduced to 94.8 mmol/l. The solutions were gassed with 95% O2/5% CO2 throughout the experiments to maintain pH 7.4. Aldrithiol, carbonyl cyanide-trifluoromethoxyphenylhydrazone (FCCP), diazoxide and dithiothreitol were from Sigma.
Measurement of NAD(P)H autofluorescence, intracellular Ca2+, and mitochondrial and cytosolic glutathione oxidation
After culture, islets from N5-WT and N5-Tr mice were perifused (~1 ml/min) simultaneously at 37°C in a chamber placed on the stage of an inverted microscope. NAD(P)H autofluorescence (λex/em 360/470 nm) was measured every 10 s and normalised, in each experiment, to the mean fluorescence level measured in N5-WT islets during the initial G0.5 (0.5 mmol/l glucose) step. To assay intracellular Ca2+ concentration ([Ca2+]i), the fluorescence ratio (λex/em 340/510 and 380/510 nm) was measured every 5 s in islets preloaded for 2 h with 2 μmol/l Fura-2 LR acetoxymethyl ester (Teflabs, USA). To measure glutathione oxidation, islets were infected by adenoviruses encoding untargeted GRX1–roGFP2 (glutaredoxin 1 fused to redox-sensitive enhanced GFP) as described [36, 37]. After 48 h, the fluorescence ratio of (mt-)GRX1–roGFP2 (λex/em 400/535 and 480/535 nm) was measured every 30s. The results were normalised to the fluorescence ratio of the probe in the presence of 10 mmol/l dithiothreitol (set to 0%, maximally reduced state), and that in the presence of 100 μmol/l aldrithiol (set to 100%, maximally oxidised state). Images were acquired using a ×20 objective and an Evolve512 camera (Photometrics, USA).
Insulin secretion
Groups of 100 islets (BL/6N, N5-WT or N5-Tr) were perifused with KRB (~1 ml/min). Insulin was measured in 2 min outflow collections by RIA using rat insulin as standard. After the experiment, islets were collected and DNA content was measured as described [38]. Insulin content was determined using the Insulin Ultra Sensitive Kit 62IN2PEG (Cisbio, France).
IPGTT and ITT
Glucose or insulin challenge were performed in the morning after a 16 h fast. Glucose (2 mg/kg) or insulin (0.75 IU/kg Actrapid, Novo Nordisk, Denmark) were injected intraperitoneally using a 25G needle. Blood glucose was measured before injection and every 15 min after injection using Freestyle Precision Neo glucometer and strips (Abbott, Belgium). For each IPGTT, the AUC above baseline was computed by the trapezoidal method, where baseline was the mean blood glucose level at time 0 for mice of the same genotype. For plasma insulin measurements, blood was collected from tail puncture using heparinised capillaries (Brand, Germany, Cat. no. 749311) before and 30 min after glucose injection. Insulin was measured in plasma by ELISA (Crystal Chem, USA, Cat. no. 90080).
Statistics
Results are shown as means ± SD for the number of islets, preparations or mice indicated. Outliers were excluded using Grubb’s test. The statistical significance of differences between groups was evaluated by two-tailed unpaired t test for two-group comparison, or by two-way ANOVA (for repeated measurements when the comparison was made between selected time points in the same trace) followed by Sidak’s test when more than two groups were compared. We used GraphPad Prism (USA) version 8.
Results
WGS
The genetic background and Nnt variant expressed in BL/6N, BL/6J, N5-WT and N5-Tr mice was assessed by PCR and WGS. As expected, the BL/6N and N5-WT mice were homozygous for WT Nnt (NntWT/WT), while the BL/6J and N5-Tr mice were homozygous for truncated Nnt (NntTr/Tr) (ESM Fig. 1). By contrast, except for Nnt, the genome of the BL/6N mouse similarly differed from that of N5-WT, N5-Tr and BL/6J mice (see ESM Results; ESM Tables 1–3; ESM Figs 1–5).
NNT is functional in N5-WT but not N5-Tr islets
Comparing islets from BL/6N and BL/6J mice, we previously showed that functional NNT is key to the glucose regulation of mitochondrial EGSH, with a much lesser impact on the glucose regulation of cytosolic EGSH by exogenous H2O2 [33]. To evaluate NNT activity in islets from N5-WT and N5-Tr mice, we tested the effects of glucose and the mitochondrial uncoupler FCCP on mitochondrial and cytosolic EGSH using the redox probes (mt-)GRX1–roGFP2 [39].
In the mitochondrial matrix, the normalised mt-GRX1–roGFP2 fluorescence ratio was ~26% in both types of female islets. It increased by ~60% upon glucose removal and was significantly reduced upon subsequent glucose stimulation in N5-WT islets but not in N5-Tr islets (Fig. 1a/ESM Fig. 6). The probe oxidation also markedly increased upon addition of FCCP in N5-WT but not N5-Tr islets (Fig. 1b). These results were reproduced in two islets from one male mouse of each type (ESM Fig. 7a). They recapitulate our observations in BL/6N and BL/6J islets infected with an adenovirus encoding Nnt vs uninfected BL/6J islets (see Figure 2ABF in [33]). Thus, NNT is functional and allows for the regulation of mitochondrial EGSH by glucose and FCCP in N5-WT but not N5-Tr islets.

Effects of glucose and FCCP on mitochondrial and cytosolic glutathione oxidation in islets from female N5-WT and N5-Tr mice. Two days after infection with Ad-mt-GRX1–roGFP2 (a, b) or Ad-GRX1–roGFP2 (c), the islets were perifused with a Krebs solution containing various glucose concentrations (Gn=n mmol/l glucose), 15 μmol/l H2O2 or 10 μmol/l FCCP as indicated. The traces were normalised to the ratio of the maximally reduced probe in the presence of 10 mmol/l dithiothreitol (set to 0%) and the ratio of the maximally oxidised probe in the presence of 100 μmol/l aldrithiol (set to 100%), as shown in ESM Fig. 6a. Data are means ± SD for 11 islets/6 preparations (a); 5 islets/3 preparations (b); or 7 islets/4 preparations (c). **p<0.01; *** p<0.001 vs the previous condition or vs G30; †††p<0.001 vs N5-WT islets. Individual traces are shown in electronic supplementary material [ESM] Fig. 7
We previously showed that, contrary to its impact on mitochondrial EGSH, NNT has negligible effect on cytosolic EGSH under control conditions [33]. However, when the GRX1–roGFP2 oxidation ratio was increased by 10 μmol/l exogeneous H2O2, removal of glucose further increased this oxidation while stimulation with 30 mmol/l glucose reduced it (Fig. 1c/ESM Fig. 6). These effects were similar in N5-WT and N5-Tr islets, thereby invalidating our earlier observation that the oxidation of cytosolic glutathione upon glucose removal in the presence of H2O2 was slightly lower in BL/6J than BL/6N islets (see Figure 3E in [33]).
Functional NNT enhances the glucose- and FCCP-mediated changes in NAD(P)H autofluorescence in N5-WT vs N5-Tr islets
We next measured the effects of glucose and FCCP on NAD(P)H autofluorescence, which measures NADH + NADPH levels in whole islets. In both islet types, NAD(P)H significantly increased upon stepwise glucose stimulation and rapidly decreased upon mitochondrial uncoupling with FCCP (Fig. 2a/ESM Fig. 8). However, the NAD(P)H autofluorescence of size-matched islets exposed to non-stimulating glucose concentrations (0.5–5 mmol/l) was slightly, although not significantly, higher in N5-Tr vs N5-WT islets, while it was similar in both types of islets exposed to 10–30 mmol/l glucose. Therefore, the rise in NAD(P)H at 10 and at 30 mmol/l glucose was significantly lower in N5-Tr vs N5-WT islets (Fig. 2b). Moreover, although the decrease in NAD(P)H induced by FCCP was initially alike in both islet types, it rapidly became slower in the N5-Tr islets, so that, after 10 min of FCCP treatment, NAD(P)H autofluorescence remained significantly higher than in 0.5 mmol/l glucose in N5-Tr but not N5-WT islets. These differences between N5-Tr vs N5-WT islets recapitulate those we reported in BL/6J vs BL/6N islets (see Figure 1B in [33]), suggesting that, as in that model, the lack of NNT prevents mitochondrial NADPH oxidation at low glucose and in the presence of FCCP without affecting NADH production (see Figure 1 C,D,F in [33]).

Dynamic effects of glucose on NAD(P)H autofluorescence, intracellular Ca2+ concentration and insulin secretion in N5-WT, N5-Tr and BL/6N islets as indicated. (a) NAD(P)H autofluorescence in islets perifused with a Krebs solution containing various glucose concentrations ranging from 0.5 to 30 mmol/l glucose (Gn=n mmol/l glucose) or G30 plus 10 μmol/l FCCP, as indicated on top of the figure. In each experiment, N5-WT and N5-Tr islets of similar sizes were perifused simultaneously in the same chamber, and their NAD(P)H autofluorescence levels were normalised to the level of fluorescence in WT islets during perifusion with G0.5 (raw fluorescence levels per pixel are shown in ESM Fig. 6b). (b) glucose-induced rise in NAD(P)H autofluorescence above G0.5 computed from data shown in (a). (c, d) Fura2-LR fluorescence ratio and dynamic GSIS in N5-WT and N5-Tr islets during stepwise stimulation with glucose. The experiments ended by membrane depolarisation with 30 mmol/l extracellular K+ in the presence of 250 μmol/l diazoxide (K30Dz). The AUC during the second half, except for the last min, of each condition was computed for statistical analysis of differences between groups and conditions. (e–h) Dynamic GSIS in female N5-Tr vs N5-WT islets (e, f), male N5-Tr vs N5-WT islets (g) or female BL/6N vs N5-WT islets (h). The AUC during the first phase (min 10–25) and second phase (min 25–70, horizontal dotted line) of GSIS was computed for statistical analysis of differences between groups and conditions. Data are means ± SD for 9 islets/5 preparations of each type (a, b) 8 islets/4 preparations (c), and for 5 (d), 3–4 (e), 8 (f), 4–6 (g) or 3 (h) islet preparations. (a) ***p˂0.001 vs G0.5 in the same islet type; ††p<0.01 vs WT islets. (b) †p<0.05 and †††p<0.001 vs WT islets. (c–h) *p<0.05; **p˂0.01; ***p<0.001 for the effect of glucose vs G0.5; †p˂0.05 vs N5-WT islets; ‡‡p<0.01 and ‡‡‡p<0.001 for the effect of K30Dz in G30. (d–f) The insulin to DNA content ratio, available for five out of ten islet preparations, was 0.15 ± 0.02 ng insulin/ng DNA in N5-WT islets and 0.14 ± 0.06 ng insulin/ng DNA in N5-Tr islets. (g) The insulin to DNA content ratio was 0.13 ± 0.07 ng insulin/ng DNA in male N5-WT islets and 0.15 ± 0.07 ng insulin/ng DNA in male N5-Tr islets. (h) The insulin to DNA content ratio was 0.27 ± 0.03 ng insulin/ng DNA in BL/6N islets and 0.25 ± 0.02 ng insulin/ng DNA in N5-WT islets. Individual traces are shown in ESM Fig. 8
Our results so far indicated that restoring the NntWT/WT genotype in islets with a BL/6J background suffice to restore the glucose and FCCP regulation of mitochondrial glutathione oxidation and NAD(P)H levels to a similar extent as that previously observed in BL/6N mice. We therefore considered it to be a good model to test the precise role of NntWT on GSIS in BL/6J mice.
Functional NNT on a BL/6J background does not restore BL/6N-like GSIS
We previously showed that GSIS is significantly greater in BL/6N vs BL/6J islets because of differences in Ca2+-induced exocytosis and metabolic amplification of insulin secretion in the presence of a normal rise in [Ca2+]i (see Figure 4 C,I in [33]). Accordingly, the [Ca2+]i changes following stepwise glucose stimulation were almost identical in N5-WT and N5-Tr islets (Fig. 2c and related panel in ESM Fig. 8). Under these conditions, the stimulation of insulin secretion by stepwise glucose stimulation or by a sudden increase in glucose concentration from 0.5 to 10 or 15 mmol/l was slightly (1.2–1.4 times), although not significantly, higher in N5-WT vs N5-Tr islets, as was the response to 30 mmol/l extracellular K+ (Fig. 2d–f and related panels in ESM Fig. 8). We also tested GSIS in islets from male mice (Fig. 2g and related panel in ESM Fig. 8), in which the second phase of insulin secretion in response to 15 mmol/l glucose was ~1.4 times larger in N5-WT vs N5-Tr islets, but this difference was again not statistically significant (p = 0.27). In both females and males, the insulin to DNA content ratio was similar in N5-WT and N5-Tr islets (see legend to Fig. 2). The lack of any significant difference in GSIS was surprising, as we had expected that, because of their NntWT expression, N5-WT islets would have a greater GSIS, as do BL/6N islets. We therefore compared GSIS in islets from female BL/6N and N5-WT mice (Fig. 2h and related panel in ESM Fig. 8) and found that both first and second phases of GSIS were 2.4 times greater in BL/6N vs N5-WT islets despite their similar insulin to DNA content ratios (see legend to Fig. 2). Altogether, these results suggest that, despite its unequivocal role in the glucose regulation of NADPH and mitochondrial EGSH, NNT contributes only partly to the higher GSIS in BL/6N vs BL/6J islets.
Glucose tolerance is only moderately improved in N5-WT vs N5-Tr mice
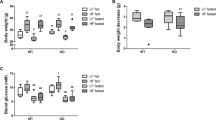
So far, the reduction of GSIS and the alteration of glucose tolerance in BL/6J mice have been largely, although not exclusively, attributed to their lack of functional NNT [13, 15, 26]. Considering our GSIS data, we questioned that hypothesis by evaluating glucose tolerance in N5-WT vs N5-Tr mice. We first checked the glycaemic response during an IPGTT in female N5-WT and N5-Tr mice and compared them to BL/6N and BL/6J mice as controls (Fig. 3a–d/ESM Fig. 7). As expected, glucose tolerance was significantly better in BL/6N than BL/6J mice, with a peak blood glucose at ~13 mmol/l in BL/6N vs ~27 mmol/l in BL/6J mice. By contrast, glucose tolerance was only slightly, although significantly, better in N5-WT than in N5-Tr mice and remained close to that observed in BL/6J mice, with a peak blood glucose at ~22 mmol/l in N5-WT vs ~25 mmol/l in N5-Tr mice. Similar differences in glucose tolerance between these four types of mice were observed in fasting–refeeding tests (ESM Fig. 9a–b). In another series of IPGTT experiments (Fig. 3e–f) the increase in plasma insulin at min 30 of the IPGTT was not significantly different while blood glucose was lower in N5-WT than N5-Tr mice, suggesting that in vivo GSIS may be slightly larger in N5-WT vs N5-Tr mice. Body weight (ESM Fig. 10a), insulin sensitivity evaluated by ITT (Fig. 3g–h and related panels in ESM Fig. 7), and beta cell mass (see ESM Methods and ESM Fig. 10 b) were similar in N5-WT and N5-Tr mice.

Effect of NNT on glucose tolerance in female mice. (a, b; c, d; and i, j) Blood glucose levels and corresponding AUC during an IPGTT after an overnight fast in N5-Tr, N5-WT, BL/6N and BL/6J mice as indicated. (e, f) Blood glucose and plasma insulin levels in the fasted state (0 min) and 30 min following glucose injection during an IPGTT in N5-WT and N5-Tr mice. (g, h) Blood glucose levels during an ITT in N5-WT and N5-Tr mice, and corresponding slope of the curves during the first 30 min. Mice were used between 3 and 6 months of age. Data are means ± SD or means ± SD with individual data points for 8 (a, b), 5 (c, d), 5 (e–h), and 4 (i, j) mice of each type. (b, d, h, j) The statistical significance of difference between groups was assessed by unpaired two-tailed t- test: †p<0.05 and †††p<0.001 vs N5-WT or BL/6N mice. (e) **p˂0.01 and ***p<0.001 vs time 0; †p˂0.05 vs N5-WT mice. (f) *p<0.05 for global time effect by two-way ANOVA (before post hoc testing). Individual traces are shown in ESM Fig. 7
To ascertain whether, despite their functional NNT, N5-WT mice display BL/6J-like glucose tolerance, we conducted a new series of IPGTTs comparing side-by-side BL/6N vs N5-WT mice. As shown in Fig. 3i–j and related panels in ESM Fig. 7, the AUC during IPGTT was markedly higher in N5-WT than BL/6N mice, with a peak of blood glucose at ~23 vs ~13 mmol/l.
Altogether, these in vivo results are compatible with our in vitro GSIS data and suggest that the lack of functional NNT only contributes to a moderate extent to the alteration of glucose tolerance in BL/6J vs BL/6N female mice, in agreement with a previous study [15]. Because mouse glucose tolerance is known to be worse in males than females [40] and in vitro GSIS was slightly but not significantly higher in N5-WT vs N5-Tr male islets, we also evaluated the glucose tolerance of N5-WT and N5-Tr male mice. As shown in Fig. 4a,b and related panels in ESM Fig. 7, the AUC during IPGTT was 30% lower in N5-WT than N5-Tr male mice, with a peak of blood glucose at ~20 mmol/l 15 min after injection in N5-WT vs ~26 mmol/l 30 min after injection in N5-Tr mice. Furthermore, the blood glucose during a fasting–refeeding test was slightly (~15%) but not significantly lower in N5-WT than N5-Tr male mice (ESM Fig. 9c). In another series of IPGTT experiments (Fig. 4c,d), plasma insulin at min 30 of the IPGTT was 20% higher while blood glucose was significantly lower in N5-WT compared with N5-Tr mice, strongly suggesting in vivo GSIS is improved in N5-WT vs N5-Tr male mice. By contrast, the slope of blood glucose measured during the 30 first min of the ITT was similar in both types of mice (Fig. 4e–f and related panels in ESM Fig. 7). These results indicate that, although the lack of NNT has a stronger impact on glucose tolerance in male vs female N5BL/6J mice, it contributes to a maximum of ~30% of the difference in AUC during IPGTT in BL/6J vs BL/6N mice. For ease of comparison, Table 1 lists the AUC during IPGTTs in the different types of mice tested in this study and their relative increase above the AUC in female BL/6N mice.

Effect of NNT on glucose tolerance in male mice. (a, b) Blood glucose levels and corresponding AUC during an IPGTT after an overnight fast in N5-WT and N5-Tr mice. (c, d) Blood glucose and plasma insulin levels in the fasted state (0 min) and 30 min following IPGTT. (e, f) Blood glucose levels during an ITT in N5-WT and N5-Tr mice and corresponding slope of the curves during the first 30 min. Mice were used between 3 and 6 months of age. Data are means ± SD for seven mice (a, b), 2–5 mice, because of the exclusion of data below the detection limit of insulin assay (c, d), and 6–8 mice (e, f). (b, f) Unpaired two-tailed t test. ††p<0.01 vs N5-WT mice. (c, d) *p<0.05 and ***p<0.001 vs time 0; †p<0.05 and †††p<0.001 vs N5-WT mice 30 min following IPGTT. Individual traces are shown in ESM Fig. 7
Discussion
This study comparing N5BL/6J mice with or without functional NNT (N5-WT and N5-Tr mice) on a WGS-confirmed BL/6J background argues against a major role for NNT in GSIS and glucose tolerance despite confirmation of its role in the regulation of NADPH and mitochondrial EGSH by glucose and FCCP. Indeed, glucose tolerance in vivo and GSIS in vitro differed only slightly between N5-WT and N5-Tr mice, whereas BL/6N mice had much better glucose tolerance and much higher GSIS than N5-WT mice even though both types of mice express functional NNT. These results have major implications regarding current views on the role of mitochondrial NADPH in GSIS, and about the importance of NNT genotyping in BL/6 mice of unclear origin in metabolic studies [13].
We have previously shown that, in beta cells, NNT reduces NADPH and increases mitochondrial EGSH at non-stimulating glucose concentrations by operating in the reverse mode, as it does when mitochondria are uncoupled with FCCP. Upon glucose stimulation, NADPH progressively switches from the reverse to the forward mode of operation, thereby leading to an increase in NADPH and a progressive reduction in mitochondrial EGSH across a wide range of glucose concentrations [33]. Here, we used these effects of extreme low and high glucose and of FCCP on mt-GRX1–roGFP2 oxidation and NAD(P)H autofluorescence to validate the different functional status of NNT in N5-WT and N5-Tr mouse islets. Our comparison of N5-WT to N5-Tr islets indeed fully reproduced the differences in NADPH autofluorescence and mitochondrial EGSH regulation we previously reported between BL/6N and BL/6J mice. The functional status of NNT in isolated islets therefore corresponded to the genotype of the mice, validating the N5BL/6J mouse model for testing the impact of NNT function on other processes, such as glucose tolerance and GSIS.
It has been proposed that mitochondrial NADPH not only affects the mitochondrial redox state but could also, through the action of substrate shuttles, increase NADPH in the cytosol [41,42,43,44]. Cytosolic NADPH can amplify insulin secretion in a GRX1-dependent manner [7, 8], for example through redox-regulated de-SUMOylating of exocytotic proteins by sentrin/small ubiquitin-like modifier specific peptidase 1 [35]. However, the protective effect of glucose against cytosolic glutathione oxidation by exogenous H2O2 was not significantly altered by the lack of NNT. This glucose effect may therefore depend on cytosolic sources of NADPH, such as the pentose phosphate pathway [9], and on other mitochondrial enzymes that provide both NADPH and metabolic intermediates for mitochondrial export to the cytosol [41]. Further investigation of that question was beyond the scope of this study.
Our data show that normal NNT expression only slightly improved GSIS on the BL/6J background. Interestingly, N5 islets with or without NNT activity kept a lower GSIS than BL/6N islets. Moreover, female N5-WT mice displayed significantly lower glucose tolerance than BL/6N mice although both types of mice express NntWT. The AUC during IPGTT was, however, slightly but significantly higher in N5-Tr vs N5-WT mice. Moreover, when we compared all types of female mice tested in this study, the AUC during IPGTT was similar in BL/6J mice and N5-Tr mice and slightly but significantly higher than in N5-WT mice. This suggests that, contrary to what has been suggested by Freeman et al [26] but in agreement with the seminal paper by Toye et al in which the impact of NNT truncation with seven other genetic loci only accounted for 35% of the increase in IPGTT AUC in BL/6J mice [15], NNT is responsible for only a small part (~15–20%) of the increase in IPGTT AUC in BL/6J vs BL/6N female mice, a proportion that may rise to about 30% in males. We cannot totally exclude the possibility that epistatic interactions modulate the impact of Nnt inactivation on insulin secretion and glucose tolerance depending on the mouse genetic background. However, we consider it more likely that genetic variants that are common to BL/6J, N5-WT and N5-Tr mice vs BL/6N mice, or vice versa, contribute largely to the differences in glucose tolerance between BL/6N and N5-WT mice, as discussed earlier [13, 17]. Among other things, the impact of variants that affect Adcy5 and Rptor may be worth investigating. Adcy5 encodes adenylate cyclase 5, which is involved in cyclic AMP production and modulation of insulin secretion, and presents a variant that confers a higher risk of developing type 2 diabetes in humans [45, 46]. Rptor, which encodes Raptor, involved in the mTORC1 signalling pathway, is important for the maturation of beta cells and maintenance of their secretory function [47]. In addition, changes in glucose tolerance may also result from variants in genes that affect the gut microbiota [48], or from variants that affect gene expression through epigenetic changes.
Finally, compared with N5-Tr female mice that displayed discreetly lower glucose tolerance compared with N5-WT female mice, N5-Tr male mice were markedly less glucose tolerant compared with N5-WT males during an IPGTT. These results are in good agreement with studies showing that male mice are more prone to developing glucose intolerance than female mice [40]. Nevertheless, the effect of lack of NNT on glucose tolerance in male mice only accounted for 30% of the difference in AUC between BL/6J and BL/6N female mice.
In conclusion, on a WGS-confirmed BL/6J background, the lack of NNT activity in isolated islets significantly alters their regulation of NADPH autofluorescence and mitochondrial EGSH by glucose and FCCP. However, it only contributes to a minor extent (by ~15% in females to ~30% in males) to the alteration of glucose tolerance during an IPGTT, while it has little impact on GSIS in isolated islets from female mice. Therefore, although testing for NntWT expression in BL/6 mice with unknown or complex history of BL/6J and BL/6N intercrossing is important to assess their regulation of mitochondrial NADPH and EGSH, it will not guarantee that their glucose tolerance is as good as in BL/6N mice (if they express NntWT), or as bad as in BL/6J mice (if they express NntTr).
Abbreviations
- BL/6J:
-
C57BL/6J
- BL/6N:
-
C57BL/6N
- [Ca2+]i :
-
Intracellular Ca2+ concentration
- EGSH :
-
Glutathione redox state
- FCCP:
-
Carbonyl cyanide-trifluoromethoxyphenylhydrazone
- GRX1:
-
Glutaredoxin 1
- GSIS:
-
Glucose-stimulated insulin secretion
- mt:
-
Mitochondria-targeted
- NNT:
-
Nicotinamide nucleotide transhydrogenase
- N5-Tr:
-
N5/BL mice expressing NntTr
- N5-WT:
-
N5/BL mice expressing NntWT
- roGFP2:
-
Redox-sensitive enhanced GFP
- Tr:
-
Truncated
- WGS:
-
Whole genome sequencing
- WT:
-
Wild-type
References
Henquin JC (2000) Triggering and amplifying pathways of regulation of insulin secretion by glucose. Diabetes 49(11):1751–1760. https://doi.org/10.2337/diabetes.49.11.1751
Ferrannini E, Mari A (2014) β-Cell function in type 2 diabetes. Metabolism 63(10):1217–1227. https://doi.org/10.1016/j.metabol.2014.05.012
Vistisen D, Witte DR, Tabak AG et al (2014) Patterns of obesity development before the diagnosis of type 2 diabetes: the Whitehall II cohort study. PLoS Med 11(2):e1001602. https://doi.org/10.1371/journal.pmed.1001602
Taylor R, Al-Mrabeh A, Zhyzhneuskaya S et al (2018) Remission of Human Type 2 Diabetes Requires Decrease in Liver and Pancreas Fat Content but Is Dependent upon Capacity for β Cell Recovery. Cell Metab 28(4): 547–556; e541-e543. https://doi.org/10.1016/j.cmet.2018.07.003
Prentki M, Matschinsky FM, Madiraju SR (2013) Metabolic signaling in fuel-induced insulin secretion. Cell Metab 18(2):162–185. https://doi.org/10.1016/j.cmet.2013.05.018
Nicholls DG (2016) The Pancreatic β-Cell: A Bioenergetic Perspective. Physiol Rev 96(4):1385–1447. https://doi.org/10.1152/physrev.00009.2016
Ivarsson R, Quintens R, Dejonghe S et al (2005) Redox control of exocytosis: regulatory role of NADPH, thioredoxin, and glutaredoxin. Diabetes 54(7):2132–2142. https://doi.org/10.2337/diabetes.54.7.2132
Reinbothe TM, Ivarsson R, Li DQ et al (2009) Glutaredoxin-1 mediates NADPH-dependent stimulation of calcium-dependent insulin secretion. Mol Endocrinol 23(6):893–900. https://doi.org/10.1210/me.2008-0306
Spegel P, Sharoyko VV, Goehring I et al (2013) Time-resolved metabolomics analysis of beta-cells implicates the pentose phosphate pathway in the control of insulin release. Biochem J 450(3):595–605. https://doi.org/10.1042/BJ20121349
Ferdaoussi M, MacDonald PE (2017) Toward connecting metabolism to the exocytotic site. Trends Cell Biol 27(3):163–171. https://doi.org/10.1016/j.tcb.2016.10.003
Hart NJ, Powers AC (2019) Use of human islets to understand islet biology and diabetes: progress, challenges and suggestions. Diabetologia 62(2):212–222. https://doi.org/10.1007/s00125-018-4772-2
Andersson LE, Valtat B, Bagge A et al (2015) Characterization of stimulus-secretion coupling in the human pancreatic EndoC-betaH1 beta cell line. PLoS One 10(3):e0120879. https://doi.org/10.1371/journal.pone.0120879
Fontaine DA, Davis DB (2016) Attention to Background Strain Is Essential for Metabolic Research: C57BL/6 and the International Knockout Mouse Consortium. Diabetes 65(1):25–33
Kayo T, Fujita H, Nozaki J, E X, Koizumi A (2000) Identification of two chromosomal loci determining glucose intolerance in a C57BL/6 mouse strain. Comp Med 50(3):296–302
Toye AA, Lippiat JD, Proks P et al (2005) A genetic and physiological study of impaired glucose homeostasis control in C57BL/6J mice. Diabetologia 48(4):675–686. https://doi.org/10.1007/s00125-005-1680-z
Mekada K, Abe K, Murakami A et al (2009) Genetic differences among C57BL/6 substrains. Exp Anim 58(2):141–149. https://doi.org/10.1538/expanim.58.141
Simon MM, Greenaway S, White JK et al (2013) A comparative phenotypic and genomic analysis of C57BL/6J and C57BL/6N mouse strains. Genome Biol 14(7):R82. https://doi.org/10.1186/gb-2013-14-7-r82
Hoek JB, Rydstrom J (1988) Physiological roles of nicotinamide nucleotide transhydrogenase. Biochem J 254(1):1–10. https://doi.org/10.1042/bj2540001
Rydstrom J (2006) Mitochondrial NADPH, transhydrogenase and disease. Biochim Biophys Acta 1757(5–6):721–726
Leung JH, Schurig-Briccio LA, Yamaguchi M et al (2015) Structural biology. Division of labor in transhydrogenase by alternating proton translocation and hydride transfer. Science 347(6218):178–181. https://doi.org/10.1126/science.1260451
Kim A, Chen CH, Ursell P, Huang TT (2010) Genetic modifier of mitochondrial superoxide dismutase-deficient mice delays heart failure and prolongs survival. Mamm Genome 21(11–12):534–542
Arkblad EL, Tuck S, Pestov NB et al (2005) A Caenorhabditis elegans mutant lacking functional nicotinamide nucleotide transhydrogenase displays increased sensitivity to oxidative stress. Free Radic Biol Med 38(11):1518–1525. https://doi.org/10.1016/j.freeradbiomed.2005.02.012
Roucher-Boulez F, Mallet-Motak D, Samara-Boustani D et al (2016) NNT mutations: a cause of primary adrenal insufficiency, oxidative stress and extra-adrenal defects. Eur J Endocrinol 175(1):73–84. https://doi.org/10.1530/EJE-16-0056
Ronchi JA, Francisco A, Passos LA, Figueira TR, Castilho RF (2016) The Contribution of Nicotinamide Nucleotide Transhydrogenase to Peroxide Detoxification Is Dependent on the Respiratory State and Counterbalanced by Other Sources of NADPH in Liver Mitochondria. J Biol Chem 291(38):20173–20187. https://doi.org/10.1074/jbc.M116.730473
Meimaridou E, Goldsworthy M, Chortis V et al (2018) NNT is a key regulator of adrenal redox homeostasis and steroidogenesis in male mice. J Endocrinol 236(1):13–28. https://doi.org/10.1530/JOE-16-0638
Freeman HC, Hugill A, Dear NT, Ashcroft FM, Cox RD (2006) Deletion of nicotinamide nucleotide transhydrogenase: a new quantitive trait locus accounting for glucose intolerance in C57BL/6J mice. Diabetes 55(7):2153–2156. https://doi.org/10.2337/db06-0358
Fergusson G, Ethier M, Guevremont M et al (2014) Defective insulin secretory response to intravenous glucose in C57Bl/6J compared to C57Bl/6N mice. Mol Metab 3(9):848–854. https://doi.org/10.1016/j.molmet.2014.09.006
Meimaridou E, Kowalczyk J, Guasti L et al (2012) Mutations in NNT encoding nicotinamide nucleotide transhydrogenase cause familial glucocorticoid deficiency. Nat Genet 44(7):740–742. https://doi.org/10.1038/ng.2299
Wong N, Blair AR, Morahan G, Andrikopoulos S (2010) The deletion variant of nicotinamide nucleotide transhydrogenase (Nnt) does not affect insulin secretion or glucose tolerance. Endocrinology 151(1):96–102. https://doi.org/10.1210/en.2009-0887
Nicholson A, Reifsnyder PC, Malcolm RD et al (2010) Diet-induced obesity in two C57BL/6 substrains with intact or mutant nicotinamide nucleotide transhydrogenase (Nnt) gene. Obesity(SilverSpring) 18(10):1902–1905
Fisher-Wellman KH, Ryan TE, Smith CD et al (2016) A Direct Comparison of Metabolic Responses to High-Fat Diet in C57BL/6J and C57BL/6NJ Mice. Diabetes 65(11):3249–3261. https://doi.org/10.2337/db16-0291
Attane C, Peyot ML, Lussier R et al (2016) Differential Insulin Secretion of High-Fat Diet-Fed C57BL/6NN and C57BL/6NJ Mice: Implications of Mixed Genetic Background in Metabolic Studies. PLoS One 11(7):e0159165. https://doi.org/10.1371/journal.pone.0159165
Santos LRB, Muller C, de Souza AH et al (2017) NNT reverse mode of operation mediates glucose control of mitochondrial NADPH and glutathione redox state in mouse pancreatic beta-cells. Mol Metab 6(6):535–547. https://doi.org/10.1016/j.molmet.2017.04.004
Freeman H, Shimomura K, Cox RD, Ashcroft FM (2006) Nicotinamide nucleotide transhydrogenase: a link between insulin secretion, glucose metabolism and oxidative stress. Biochem Soc Trans 34(Pt 5):806–810. https://doi.org/10.1042/BST0340806
Ferdaoussi M, Dai X, Jensen MV et al (2015) Isocitrate-to-SENP1 signaling amplifies insulin secretion and rescues dysfunctional beta cells. J Clin Invest 125(10):3847–3860. https://doi.org/10.1172/JCI82498
Gutscher M, Pauleau AL, Marty L et al (2008) Real-time imaging of the intracellular glutathione redox potential. Nat Methods 5(6):553–559. https://doi.org/10.1038/nmeth.1212
Takahashi HK, Santos LR, Roma LP et al (2014) Acute nutrient regulation of the mitochondrial glutathione redox state in pancreatic beta-cells. Biochem J 460(3):411–423. https://doi.org/10.1042/BJ20131361
Roma LP, Duprez J, Jonas JC (2015) Glucokinase activation is beneficial or toxic to cultured rat pancreatic islets depending on the prevailing glucose concentration. Am J Physiol Endocrinol Metab 309(7):E632–E639. https://doi.org/10.1152/ajpendo.00154.2015
Meyer AJ, Dick TP (2010) Fluorescent protein-based redox probes. Antioxid Redox Signal 13(5):621–650. https://doi.org/10.1089/ars.2009.2948
Mauvais-Jarvis F (2018) Gender differences in glucose homeostasis and diabetes. Physiol Behav 187:20–23. https://doi.org/10.1016/j.physbeh.2017.08.016
MacDonald MJ, Fahien LA, Brown LJ, Hasan NM, Buss JD, Kendrick MA (2005) Perspective: emerging evidence for signaling roles of mitochondrial anaplerotic products in insulin secretion. Am J Physiol Endocrinol Metab 288(1):E1–15
Jitrapakdee S, Wutthisathapornchai A, Wallace JC, MacDonald MJ (2010) Regulation of insulin secretion: role of mitochondrial signalling. Diabetologia 53(6):1019–1032. https://doi.org/10.1007/s00125-010-1685-0
Maechler P (2013) Mitochondrial function and insulin secretion. Mol Cell Endocrinol 379:12–18. https://doi.org/10.1016/j.mce.2013.06.019
Dey S, Sidor A, O’Rourke B (2016) Compartment-specific Control of Reactive Oxygen Species Scavenging by Antioxidant Pathway Enzymes. J Biol Chem 291(21):11185–11197. https://doi.org/10.1074/jbc.M116.726968
Roman TS, Cannon ME, Vadlamudi S et al (2017) A Type 2 Diabetes-Associated Functional Regulatory Variant in a Pancreatic Islet Enhancer at the ADCY5 Locus. Diabetes 66(9):2521–2530. https://doi.org/10.2337/db17-0464
Dupuis J, Langenberg C, Prokopenko I et al (2010) New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat Genet 42(2):105–116. https://doi.org/10.1038/ng.520
Yin Q, Ni Q, Wang Y et al (2020) Raptor determines beta-cell identity and plasticity independent of hyperglycemia in mice. Nat Commun 11(1):2538. https://doi.org/10.1038/s41467-020-15935-0
Ussar S, Griffin NW, Bezy O et al (2015) Interactions between Gut Microbiota, Host Genetics and Diet Modulate the Predisposition to Obesity and Metabolic Syndrome. Cell Metab 22(3):516–530. https://doi.org/10.1016/j.cmet.2015.07.007
Acknowledgements
We thank F. B. Aissa and F. Knockaert (Université catholique de Louvain) for expert technical help, and the Genomics Platform of Université catholique de Louvain for genome sequencing and analysis. We also thank our colleague P. Gilon (Université catholique de Louvain, IREC Institute, Pole of endocrinology, diabetes and nutrition) for reading and commenting on the paper.
Data and resource availability
The article and ESM files contain all data generated during this study and information to access resources.
Authors’ relationships and activities
J-CJ is a member of the Diabetologia Editorial Board. The other authors declare that there are no relationships or activities that might bias, or be perceived to bias, their work.
Funding
This study was funded by the Action de Recherche Concertée 18/23-094 from the Communauté française de Belgique, and Grant SFD/MSD 2016 from the Société Francophone du Diabète (Paris, France) to J-CJ. J-CJ is Research Director at the Fonds de la Recherche Scientifique-FNRS, Brussels, Belgium. We also thank the Foundation against Cancer (2010-101), Belgium, for their support to the Genomics Platform of University of Louvain and de Duve Institute, and the Fonds de la Recherche Scientifique-FNRS Equipment Grant U.N035.17 for the ‘Big data analysis cluster for NGS at UCLouvain’.
Author information
Authors and Affiliations
Contributions
A-FC and HC performed the experiments. A-FC, HC and J-CJ conceived and designed the experiments and analysed the data. A-FC and J-CJ wrote the paper, all authors edited and approved the paper and. J-CJ is the guarantor of this work and, as such, had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Close, AF., Chae, H. & Jonas, JC. The lack of functional nicotinamide nucleotide transhydrogenase only moderately contributes to the impairment of glucose tolerance and glucose-stimulated insulin secretion in C57BL/6J vs C57BL/6N mice. Diabetologia 64, 2550–2561 (2021). https://doi.org/10.1007/s00125-021-05548-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-021-05548-7